Prenatal diagnosis of β-thalassaemia using fetal erythroblasts enriched from maternal blood by a novel gradient
4
0
0
Texte intégral
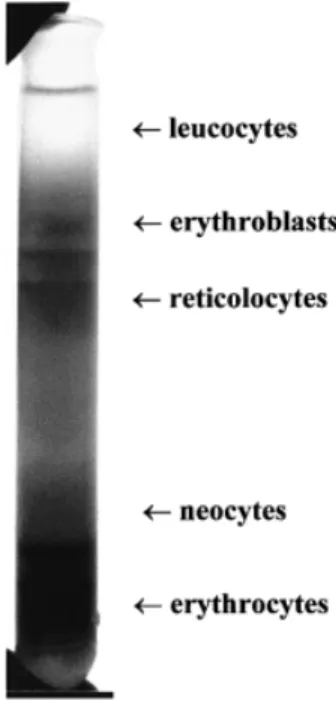
(2) E.Di Naro et al.. Figure 2. Switch diagram of globin chains.. Figure 1. Step density gradient for the enrichment of fetal erythroblasts from maternal blood.. the fetal β-globin genotype in pregnancies at risk for βthalassaemia.. Materials and methods Six couples who are carriers for one of the four most common mutations for β-thalassaemia in the Mediterranean region (β°COD.39, β⫹ IVS1–6, β⫹ IVS1–110, β°IVS1–1) were recruited for this study. Informed written consent was provided by all participants, as was the provision by the ethical committees of our respective institutions. 7.5 ml venous blood samples were drawn prior to CVS. EDTA was used as an anti-coagulant. The gestational age of the participants was 10–11 weeks. For the separation of the blood sample, the following density gradient was used: 35 ml Percoll (Phamarcia Biotech AB, Sweden.), 15 ml Gastrografin (Schering AG, Germany), 25 ml sterile isotonic saline (0.9%NaCl) and 25 ml sterile water. This was aliquoted into 10 centrifuge tubes, 10 ml for each tube. Following careful overlaying of the blood sample (250–300 µl), centrifugation was carried out at 19 000 r.p.m. for 10 min at 4°C. The erythroblast layer (Figure 1) was isolated and washed once by centrifugation with isotonic saline at 5000 r.p.m. for 10 min at room temperature. The cell pellet was subsequently washed two more times with isotonic saline by centrifugation at 5000 r.p.m. for 5 min at room temperature. The cells were then streaked onto four microscope-slides and stained with monoclonal antibodies against zeta (ζ) globin (Isolab Inc, Akron, USA), which is more specific than the γ globin chain, in order to distinguish between maternal and fetal erythroblasts (Figure 2), as previously described (Cheung et al., 1996) (Figure 3). Although ζ globin is depleted by ~week 8 of gestation, the erythroblasts have a life span of 90–120 days. For this reason, ζ globin can be detected on fetal erythroblasts at least until the end of the first trimester. Erythroblasts which had stained strongly positive for anti-ζ globin monoclonal antibodies were detected in four cases while in the remaining two cases only weakly positive erythroblasts were identifiable. Five erythroblasts from each sample were isolated by micromanipulation under a light microscope (Carl Zeiss, Jena, Germany) using an individual glass capillary for each cell. Each single erythro-. 572. Figure 3. Fetal erythroblast stained with monoclonal antibodies against zeta (ζ) globin.. Figure 4. Analysis of the 269 bp polymerase chain reactionamplified fragment of the β globin gene on 8% polyacrylamide gel. Lanes 1–4 ⫽ DNA fragments from four individual erythroblasts from four different samples; lane 5 ⫽ DNA marker VI, the two arrows show fragments of 298 and 234 bp. blast was transferred into a 0.5 ml Eppendorf PCR tube with 5µl sterile water and 50 µl PCR solution. The composition of the PCR solution was: PCR buffer (Perkin Elmer Cetus) 5 µl; Spermidine 0.5 mmol/l (1 mg/ml) 2.5 µl; 30 pm of each primer; and 200 µmol/l dNTP. 1 IU Amplitaq (Perkin Elmer Cetus) was used per reaction. The solution was previously digested with HaeIII (12 IU/50 µl mix) (Gibco-BRL, Life Technologies) at 37°C for 4 h to avoid contamination by carry over. Cellular lysis was performed by freezing at –20°C for 10 min and then by heating at 99°C for 10 min. Each sample was amplified by PCR using 30 cycles of 94°C for 30 s, 55°C for 30 s, and 72°C for 60 s and the following primers: 5⬘ACAGTTATGTTGATGGTTCTTCCATA-3⬘ and 3⬘-GTAAACTTTTAGACAGAAGACTGTT-5⬘. The amplification products were examined on an 8% acrylamide gel which revealed a fragment of the correct 269 bp size for a β globin gene (Figure 4). The sequence of.
(3) Non-invasive prenatal β-thalassaemia diagnosis Table I. Fetal and parental β-globin genotypes Case. Maternal genotype. Paternal genotype. Results from fetal erythroblasts in maternal blood. Results from CVS. 1 2 3 4. IVS1-110/N IVS1-6/N IVS1-6/N IVS1-6/N. IVS1-6/N cod.39/N cod.39/N cod.39/N. Wild type IVS1-6/N Wild type cod.39/N. Wild type IVS1-6/N Wild type cod.39/N. IVSI-110, IVSI-6 and cod.39 are mutations for β-thalassaemia; N ⫽ normal β globin allele; CVS ⫽ chorionic villus sampling.. the fragment was then determined using an automated sequencer, using the fluorescent dye termination process. This was then compared with the fetal globin genotype which had been determined on DNA obtained from CVS using the amplification refractory mutation system (ARMS) (Embury, 1995).. Results In the four instances, we detected erythroblasts which had stained strongly positive with anti-ζ globin antibodies. In each of these cases, five cells were isolated by micromanipulation under a light microscope and we were able to correctly determine fetal genotype in all cells picked, and this was confirmed from the result obtained from the CVS sample. Three different β-thalassaemia mutations were analysed in the sequenced fragments: cod.39 (C→T creating a nonsense codon in exon 2); IVSI-6 (T→C at nucleotide 6, creating an RNA processing mutation) and IVSI-110 (G→A at nucleotide 110, also creating an RNA processing mutation). The parental and fetal globin genotypes of these cases are presented in Table II. In the remaining two cases, where only weakly stained anti-ζ globin erythroblasts were detected, we determined that all the erythroblasts examined were of maternal origin because the results were not confirmed by CVS diagnosis.. Discussion We have shown that fetal erythroblasts can be reliably enriched from maternal blood using a simple, accurate and inexpensive technique which can be carried out in most modestly equipped laboratories. A further advantage of this technique is that it permits the use of smaller blood samples (⬍10 ml) than other methods currently employed, such as MACS or FACS, where generally 15–20 ml blood samples are required (Hahn et al., 1998a). The use of a Percoll gradient for separating fetal erthyroblasts from maternal blood has been reported previously (Takabayashi et al., 1995). However, we used both Percoll and Gastrografin to create a multiple gradient for more definitive separation of the erythroblasts. We have furthermore demonstrated that following immunohistochemical identification on the basis of staining with antiζ globin antibodies, single micromanipulated erythroblasts can be analysed by single cell PCR to determine the fetal genotype for the locus interrogated. In all four cases in which erythroblasts were detected by intense staining for ζ globin, the correct fetal genotype was determined. In those instances, where the erythroblasts were found to be weakly staining, they were shown by genotype analysis to be of maternal origin.. These results indicate that anti-ζ haemoglobin antibodies can be used to distinguish between maternal and fetal erythroblasts. However, since it is unlikely that any antibody will be absolutely specific, the irrefutable identification of fetal cells will have to be achieved by genetic means, such as by the use of highly polymorphic microsatellite markers (Garvin et al. 1998). In those instances in which we observed no intense anti-ζ globin staining, it is possible that no fetal erythroblasts were present in these samples; a phenomenon which both we and other researchers have observed previously (Wachtel et al., 1996; Bianchi et al., 1997; Troeger et al., 1999). It is also possible that the antibody staining was inadequate, as we have observed considerable variation in the reproducibility of this method. This could be addressed by the use of different or other more specific fetal antibodies. Although we do confirm an earlier report by the laboratory of Y.W.Kan regarding the use of enriched fetal erythroblasts for the prenatal diagnosis of fetal haemoglobinopathies (Cheung et al., 1996), the main difference between these two reports is that we have used a different enrichment procedure and that single fetal cells were individually analysed by PCR and not pooled for the PCR analyses as in the former case. Although the analysis of single cells by PCR is more problematic in view of the danger of allele drop out (Hahn et al., 1998b), we have shown that diagnostically accurate results can be obtained by pooling the results obtained from the separate analysis of several individual single cells (Garvin et al., 1998; Troeger et al., 1999). This is confirmed in this study where we examined five individual cells from each case. The robustness of our approach is apparent from the highly successful determination of the fetal genotype in those four cases where fetal erythroblasts could be readily identified by anti-ζ globin immunohistochemistry. As such, this study presents the largest series of prenatal diagnoses performed by these means to date. In conclusion, the rapidity and ease with which these analyses were performed allow us to propose that since the majority of β-thalassaemia mutations are well defined and in certain instances clustered in a small region of the β-globin gene, it should be possible to develop this technique to the extent that it can be used to non-invasively screen the majority of pregnancies at risk for this haemoglobinopathy.. Acknowledgements This work was suported by a grant from M.U.R.S.T. (40%) n.9906042485-005.. 573.
(4) E.Di Naro et al.. References Bianchi, D.W., Mahr, A., Zickwolf, G.K. et al. (1992) Detection of fetal cells with, 47,XY,⫹21 karyotype in maternal peripheral blood. Hum. Genet., 90, 368–370. Bianchi, D.W., Zickwolf, G.K., Yih, M.C. et al. (1993) Erythroid-specific antibodies enhance detection of fetal nucleated erythrocytes in maternal blood. Prenat. Diagn., 13, 293–300. Bianchi, D.W., Zickwolf, G.K., Weil, G.J. et al. (1996) Male fetal progenitor cells persist in maternal blood for as long as 27 years postpartum. Proc. Natl Acad. Sci. USA, 93, 705–708. Bianchi, D.W., Williams, J.M., Sullivan, L.M. et al. (1997) PCR quantitation of fetal cells in maternal blood in normal and aneuploid pregnancies. Am. J. Hum. Genet., 61, 822–829. Cheung, M.C., Goldberg, J.D. and Kan, Y.W. (1996) Prenatal diagnosis of sickle cell anaemia and thalassaemia by analysis of fetal cells in maternal blood. Nature Genet., 14, 264–268. de la Cruz, F., Shifrin, H., Elias, S. et al. (1998) Low false-positive rate of aneuploidy detection using fetal cells isolated from maternal blood. Fetal Diagn. Ther., 13, 380. Elias, S., Price, J., Dockter, M. et al. (1992) First trimester prenatal diagnosis of trisomy 21 in fetal cells from maternal blood. Lancet, 340, 1033. Embury, S.H. (1995) Advances in the prenatal and molecular diagnosis of the hemoglobinopathies and thalassemias. Hemoglobin, 19, 237–261. Ganshirt, D., Garritsen, H., Miny, P. and Holzgreve, W. (1994) Fetal cells in maternal circulation throughout gestation. Lancet, 343, 1038–1039. Garvin, A.M., Holzgreve, W. and Hahn, S. (1998) Highly accurate analysis of heterozygous loci by single cell PCR. Nucleic Acids Res., 26, 3468–3472. Ga¨ nshirt-Ahlert, D., Bo¨ rejesson-Stoll, R., Burschyk, M. et al. (1993) Detection of fetal trisomies 21 and 18 from maternal blood using triple gradient and magnetic cell sorting. Am. J. Reprod. Immun., 30, 194–201. Hahn, S., Sant, R. and Holzgreve, W. (1998a) Fetal cells in maternal blood: current and future perspectives. Mol. Hum. Reprod., 4, 515–521. Hahn, S., Garvin, A.M., Di Naro, E. and Holzgreve, W. (1998b) Allele drop out can occur in alleles differing by a single nucleotide and is not alleviated by preamplification nor minor template increments. Genetic Testing, 2, 351–355. Holzgreve, W. (1997) Will ultrasound-screening and ultrasound-guided procedures be replaced by non-invasive techniques for the diagnosis of fetal chromosome anomalies? [Editorial.] Ultrasound Obstet. Gynecol., 9, 217–219. Holzgreve, W., Garritsen, H.S. and Ganshirt Ahlert, D. (1992) Fetal cells in the maternal circulation. J. Reprod. Med., 37, 410–418. Holzgreve, W., Ghezzi, F., Di Naro, E. et al. (1998) Disturbed Fetomaternal cell traffic in preeclampsia. Obstet. Gynecol., 91, 669–672. Schro¨ der, J., Tilikainen, A. and de la Chapelle, A. (1974) Fetal leucocytes in maternal circulation after delivery. Transplantation, 17, 346–360. Sekizawa, A., Watanabe, A., Kimura, T. et al. (1996) Prenatal diagnosis of the fetal RhD blood type using a single nucleated erythrocyte from maternal blood. Obstet. Gynecol., 87, 501–505. Slunga Tallberg, A., el Rifai, W., Keinanen, M. et al. (1996) Maternal origin of transferrin receptor positive cells in venous blood of pregnant women. Clin. Genet., 49, 196–199. Takabayashi, H., Kuwabara, S., Ukita, T. et al. (1995) Development of noninvasive fetal DNA diagnosis from maternal blood. Prenat. Diagn., 15, 74–77. Troeger, C., Zhong, X.Y., Burgemeister, R. et al. (1999) Approximately half of the erythroblasts in maternal blood are of fetal origin. Mol. Hum. Reprod., 5, 1162–1165. Wachtel, S.S., Sammons, D., Manley, M. et al. (1996) Fetal cells in maternal blood: recovery by charge flow separation. Hum. Genet., 98, 162–166. Weatherall, D.J. (1995) The thalassemias. In Beutler, E., Lichtman, M.A., Coller, B.S. and Kipps, T.J. (eds), Williams Hematology. McGraw-Hill, New York, USA, pp. 581–615. Received on December 13, 1999; accepted on March 23, 2000. 574.
(5)
Figure

Documents relatifs
