Evolution conjointe des stratégies d'appariement et de dispersion
146
0
0
Texte intégral
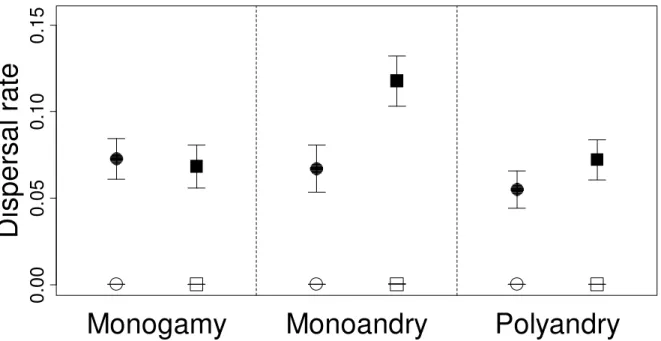
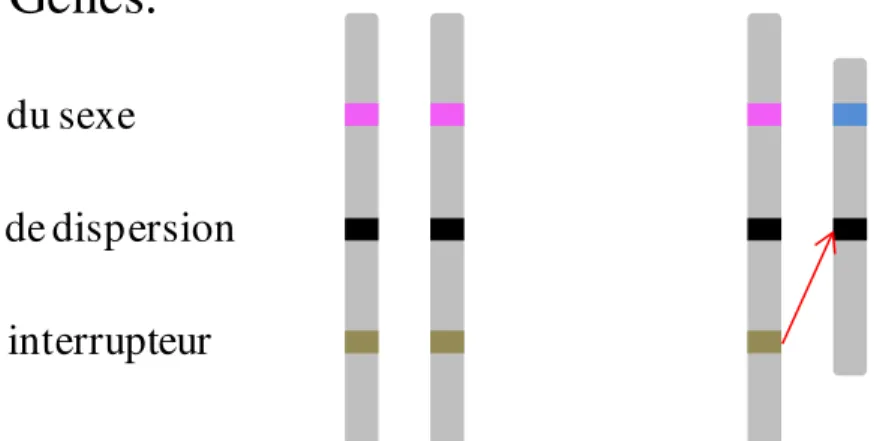
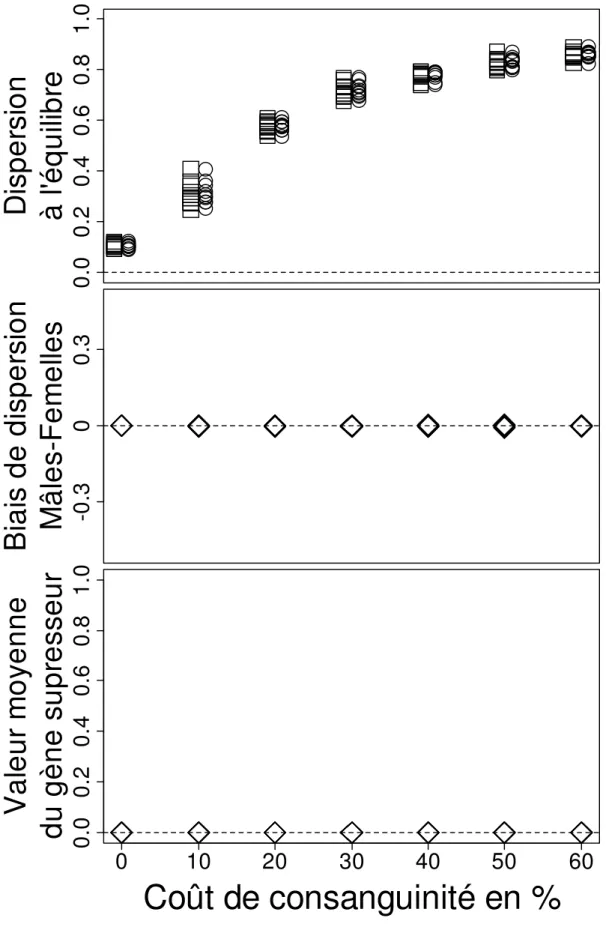
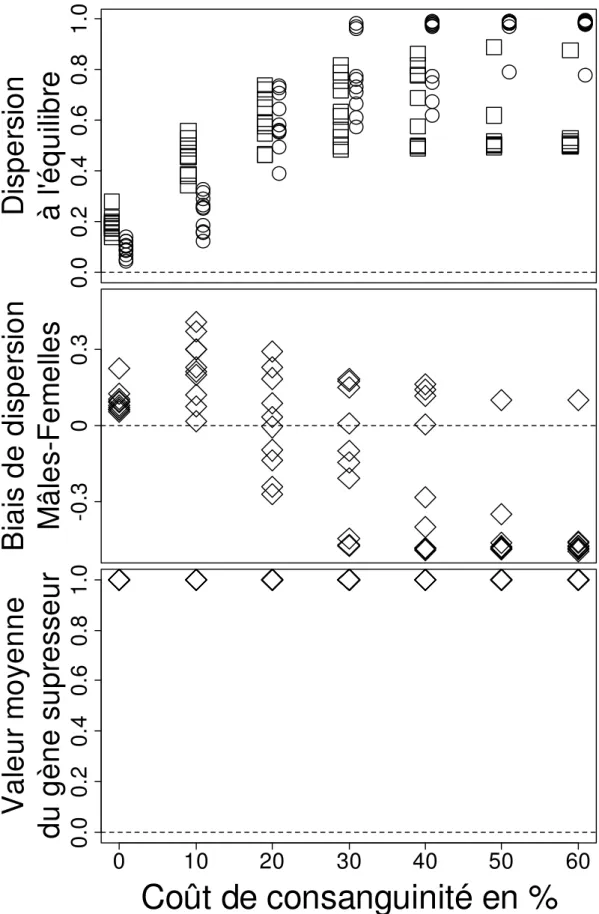
Figure

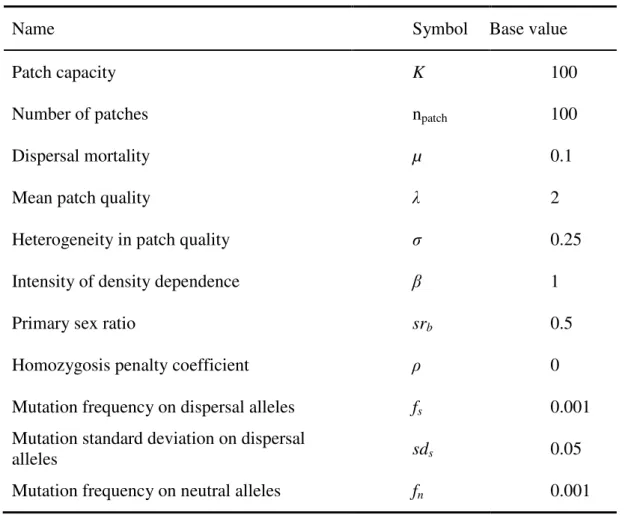
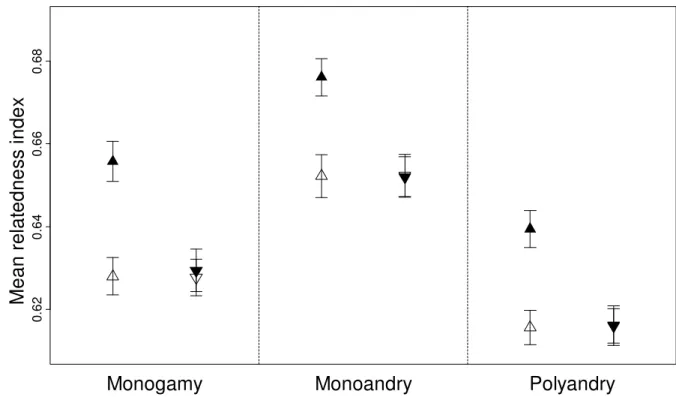
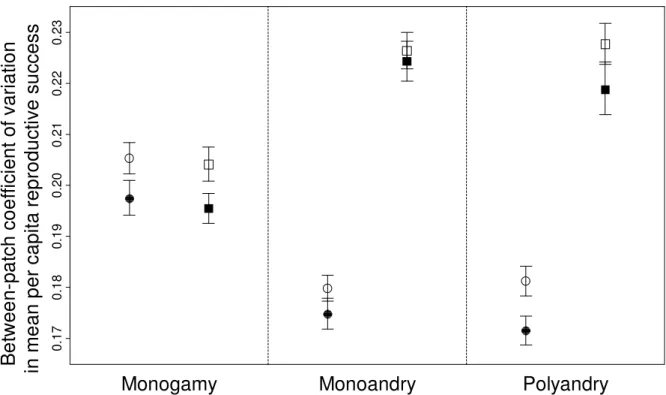
+7
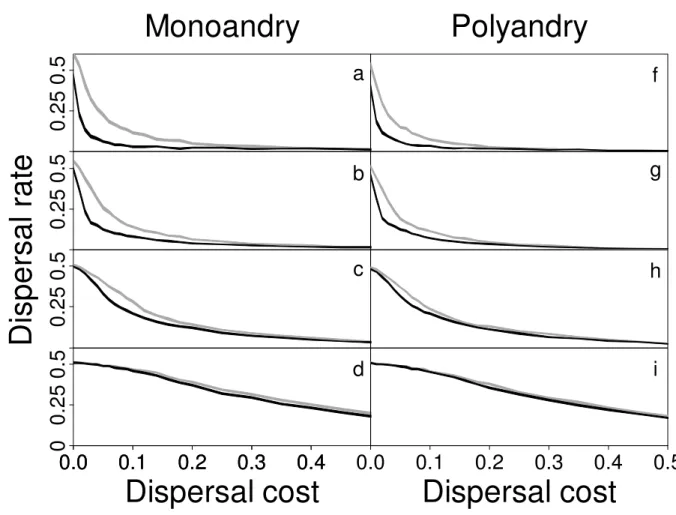

Documents relatifs