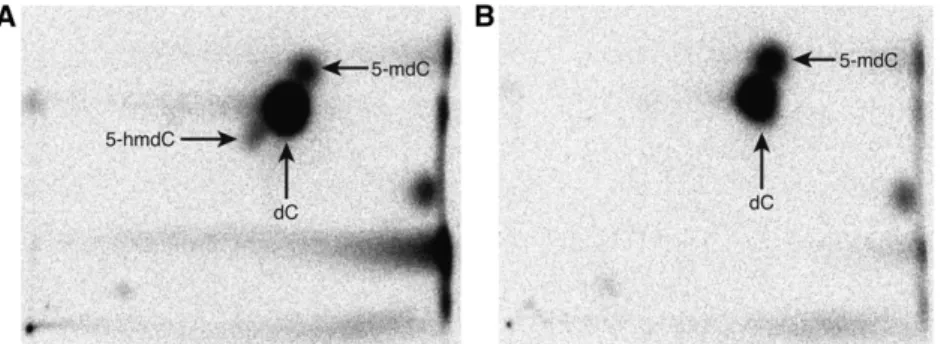
5-Hydroxymethylcytosine Is Not Present in Appreciable Quantities in Arabidopsis DNA
Texte intégral
Figure
Documents relatifs
[r]
Modifier le code de la fonction main pour que la ligne bris´ ee reliant les 4 points soit afficher lors de l’affichage du message ”Dessin courbe de B´ ezier”.. D` es que le
[r]
On the following session, a theatre-play approach was used for presenting the explanation, using the following physical material as stage props: two square
This fact confirms the previously obtained results concerning the growing-finishing period and shows that a better utilization of the diets rich in whey, by maintaining milk
Un sondage réalisé à la même époque dans des éle- vages pratiquant l’insémination artificielle sur 1 6 91 truies de race pure et 194 truies croisées confirme cette
creen, fonctionnant en vraie grandeur, de go cm de diamètre et de 400 - 5 oo y de maille. Les condi- tions de fonctionnement optimales de ce type de tamis
La coloration de la viande a été légèrement trop foncée chez les veaux qui recevaient l’aliment riche en protéines de poisson non additionné de complexant du fer,
