Enhanced visual exploration for real objects compared to pictures during free viewing in the macaque monkey
13
0
0
Texte intégral
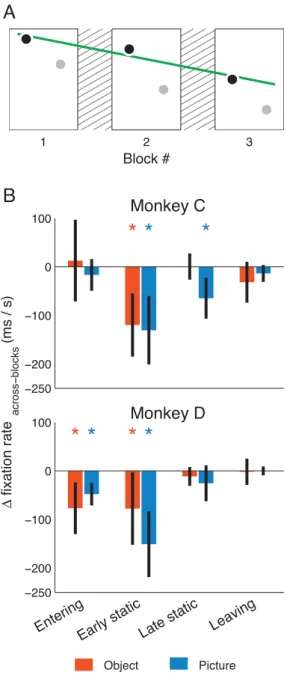
Figure



+3


Documents relatifs