Biology of 'Borrelia burgdorferi'-infected and non-infected 'Ixodes ricinus' ticks in the context of climate change
202
0
0
Texte intégral
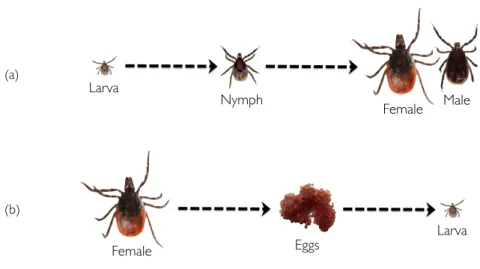
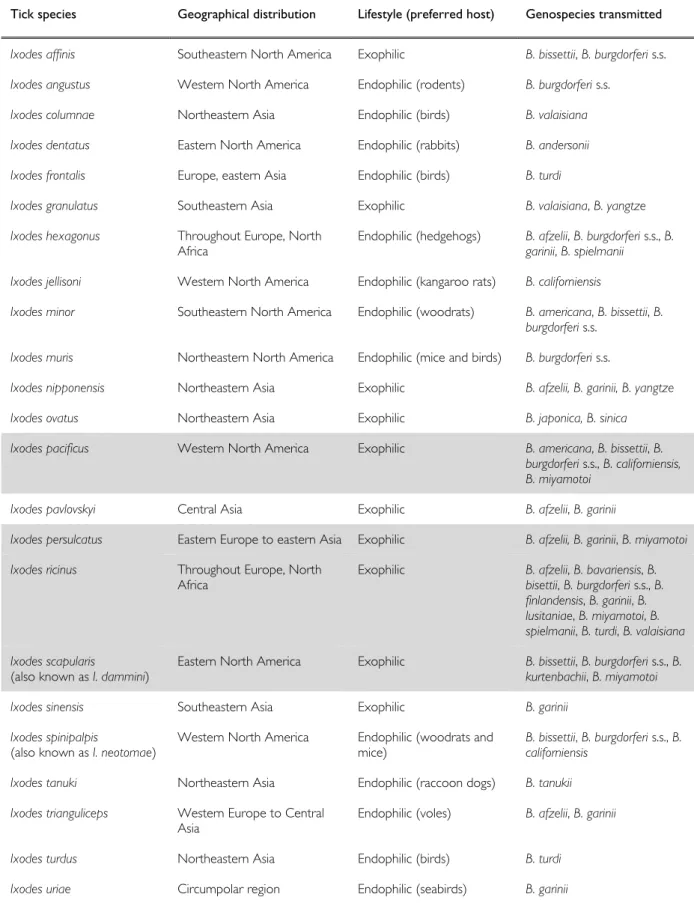
Figure

+7



Outline
Documents relatifs