Anticipation via canards in excitable systems
19
0
0
Texte intégral
(2)
(3)
(4)
(5)
(6)
Figure
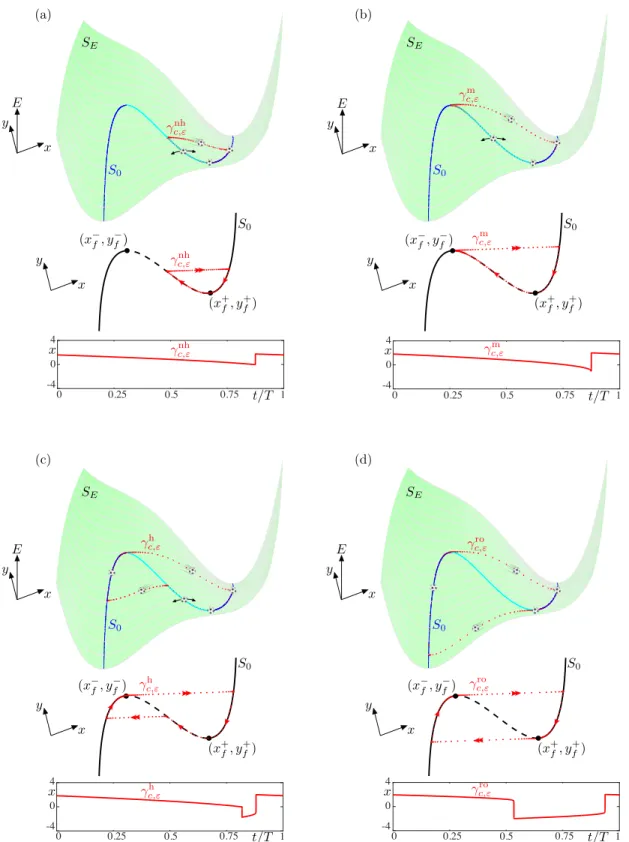
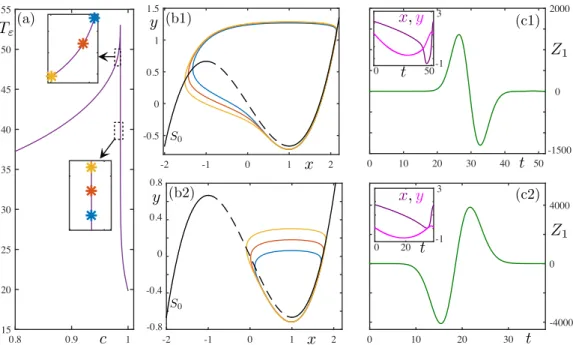
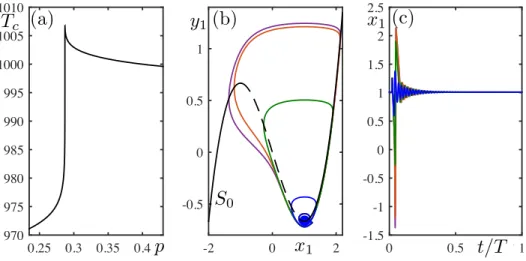
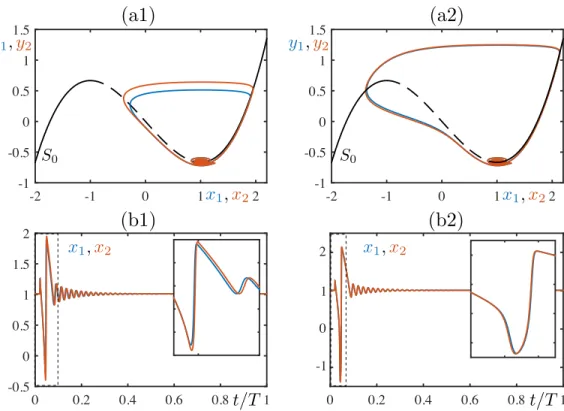
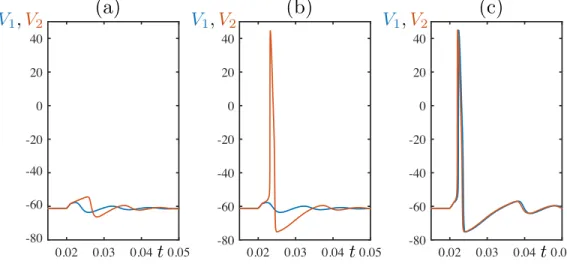
+7
![FIG. 5. Behavior of system (8) under pulse perturbation for p ≈ 0.27141141, α = 0.05, τ ∈ [0, 1]: (a) min(x 1 ) − min(x 2 ) vs τ ∈ [0, 1]](https://thumb-eu.123doks.com/thumbv2/123doknet/13595616.423379/12.918.177.749.74.485/fig-behavior-of-system-pulse-perturbation-α-τ.webp)
![FIG. 6. Behavior of system (8) under pulse perturbation for p ≈ 0.28723689613, α = 0.05, τ ∈ [0, 1]: (a) min(x 1 ) − min(x 2 ) vs τ ∈ [0, 1]](https://thumb-eu.123doks.com/thumbv2/123doknet/13595616.423379/13.918.178.749.72.481/fig-behavior-of-system-pulse-perturbation-α-τ.webp)



Documents relatifs
This process consists in submitting load injection to the black box (see Figure 3). The workload is increased through several steps, until reaching the maximal estimated load C max
Anticipation play a very important role in the human- human as well as human-machine spoken interaction, because it influences the speed of response in conversation
Each sentence was thoroughly assessed by the four in- terpreters who recorded the number of rhythmic groups, the order number of the rhythmic group with the antici- patory nucleus,
The eclectic and powerful representation and calculation capabilities of the DSMs can be combined into a Multi Domain Matrix, MDM, by the DMMs Domain Mapping Matrices in order
Abstract. Systems biology of plants offers myriad opportunities and many challenges in modeling. A number of technical challenges stem from paucity of computational
Thus, working with model systems provide the possibility of reproducing specific crack patterns as a function of the mechanical properties of the system, and key parameters such as
To further these findings, we conducted a new coordinate-based meta-analysis of functional Magnetic Resonance Imaging (fMRI) studies comparing appetitive
2) Global case: In order to understand the accuracy of the Bayesian network, we depict in Figure 5.a estimated session durations versus the actual ones over a ten days period.
