Page | 1
INRA Val de Loire Unité Expérimentale de Bourges 18390 OSMOY ET INRA Occitanie Unité GenPhySE 31320 CASTANET TOLOSAN Spécialité Agriculture
Parcours Enjeux et Défis des Productions Animales
Mémoire de Fin d’Etudes
157
èmepromotion
Maîtres de Stage : Mme Céline MARQUIS et Mme Rachel RUPP
Tuteur : Mme Dorothée BIZERAY FILOCHE
Année
2017-2018
Evaluation du transfert d’immunité passive et de la qualité
du colostrum dans les lignées divergentes « Longévité »
caprines de Bourges
Page | 2
Résumé
Depuis les années 2000, de nouveaux paramètres de sélection chez les caprins apparaissent, comme la morphologie mammaire ou la résistance aux mammites. Au vu de l’importance de la santé et du bien-être animal, une sélection sur la longévité des animaux en élevage est devenue un enjeu majeur pour la filière. Cette longévité est le résultat de mécanismes adaptatifs qui opèrent à différents niveaux d’organisation physiologique et notamment dans le jeune âge. La santé et le bien-être des chevreaux sont les premiers déterminants de la carrière de l’animal et sont sous l’influence de l’ingestion du colostrum. En effet, celui-ci permet d’apporter tous les éléments nécessaires au développement du nouveau-né, au transfert d’immunité et à la croissance de l’animal. Sur ces sujets, les caprins sont peu documentés. L’évaluation du transfert d’immunité passive et de la qualité du colostrum ainsi que leur rôle sur la santé, le bien-être et la croissance des chevreaux est essentielle. Dans cette continuité, le projet ACTIVEGOAT, reposant sur la création de lignées divergentes, a été lancé pour étudier le rôle de la génétique sur la longévité fonctionnelle, notamment dans le jeune âge. Au sein de ce projet, une étude a été mise en place afin d’évaluer le transfert d’immunité passive et la qualité du colostrum sur la santé, le bien-être et la croissance des chevreaux. Pour cette étude, 167 chevreaux, dont 37 primipares, ont été analysés. A l’issue de cette première analyse, il s’avère que la capacité d’absorption des immunoglobulines G par les chevreaux et la qualité du colostrum des mères possèdent une forte variabilité interindividuelle (LEVIEUX, 1984; EHRHARDT et al., 2014). Le transfert
d’immunité passive semble être fortement lié aux modalités de distribution et possède un fort impact sur les troubles digestifs. L’étude a également permis de mettre en évidence un effet du volume de colostrum produit sur la qualité de celui-ci. En revanche, le rôle de la qualité du colostrum sur le transfert d’immunité passive n’a pas pu être analysé suite à un retour de données inexploitables. A ce stade, l’étude n’a pas démontré d’effet de la lignée sur les variables étudiées. Cela pourrait être lié à un effectif insuffisant pour une analyse de paramètres génétiques. Des pistes d’amélioration du protocole ont été évoquées, notamment sur le transfert d’immunité passive et la qualité du colostrum par l’ajout de nouvelles mesures et analyses.
Mots clés : longévité, transfert d’immunité passive, chevreau, colostrum, santé, bien-être, croissance
Abstract
Since the 2000s, new parameters of selection in goats have appeared, such as mammary morphology or mastitis resistance. In view of the importance of health and animal welfare, a selection focusing on longevity of livestock became a major issue for the goat sector. This longevity is the result of adaptive mechanisms that operate at various levels of physiological organization, in particular in young animals. The health and welfare of goat kids are the deciding factors of the animal’s development and are dependent on the ingestion of colostrum. This provides all the elements necessary for the development of the newborn, the transfer of immunity and the growth of the animal. On these matters, goats are poorly documented. The evaluation of the transfer of passive immunity and colostrum quality as well as their roles on health, welfare and growth of goat kids is essential. In this continuity, the ACTIVGOAT project, which is based on divergent lines, was launched with the objective to study the genetic role on functional longevity and in particular in young animals. Within this project, a study was set up to evaluate the transfer of passive immunity and colostrum quality on health, welfare and growth of goat kids. For this study, 167 goat kids, 37 of which are primiparous, were analyzed. At the end of this first analysis, it turns out that the immunoglobulin absorption capacity and the colostrum quality possess a great inter-individuals variability. The transfer of passive immunity seems to be strongly bound to the modalities of distribution of colostrum and possesses a strong impact on digestive disorders. The study also highlighted an effect of the volume of colostrum produced on its quality. However, the role of colostrum quality on passive transfer immunity could not be analyzed following a return of unusable data. At this point, the study didn’t demonstrate any effect of the line on the variables studied. This could be related to insufficient population for an analysis of genetic parameters. New ways for improving the protocol were evoked, in particular on the transfer of passive immunity and colostrum quality by the addition of new measures and analysis.
Page | 3
Remerciements
Je tiens tout d’abord à remercier mes encadrantes de stage, Céline MARQUIS et Rachel RUPP, pour m’avoir donné l’opportunité de réaliser ce stage. Je les remercie également pour leur disponibilité, leur suivi, mais aussi pour tous les conseils, l’aide et les connaissances qu’elles m’ont apporté.
Je remercie également ma tutrice, Dorothée BIZERAY-FILOCHE, ainsi que l’ensemble des enseignants d’UniLaSalle Beauvais qui ont toujours été là en cas de besoin et pour leur soutien.
Un grand merci à l’ensemble des personnes de l’UE de Bourges, pour l’accueil chaleureux qu’ils m’ont offert et leur bonne humeur. Merci de m’avoir permis de visiter l’atelier ovin et m’avoir fait participer à de nombreux travaux d’expérimentation sur l’atelier caprin ainsi que pour tous les bons moments que j’ai passé à l’INRA. Je remercie particulièrement l’équipe de l’atelier caprin : Thierry FASSIER, Anthony RETORD, Pascal CHEVESSON, Philippe MARECHAL, Alain RICHETIN et Didier AUGERAT, pour toute l’aide qu’ils ont su me fournir lors de mes 3 mois et demi de présence sur l’élevage.
Merci également à l’Unité GenPhySE ainsi qu’à l’ensemble des stagiaires et thésards, pour leur accueil chaleureux et l’aide qu’ils ont su m’apporter.
Merci à Christophe HUAU, pour sa disponibilité, sa sympathie, mais aussi pour son aide dans mes débuts sur le logiciel SAS et la collecte de données à l’UE de Bourges.
Merci à Fabien CORBIERE, qui m’a apporté de nombreux conseils sur le protocole ainsi que sur mes résultats et leur interprétation.
Je remercie également Charlotte ALLAIN qui m’a permis d’aller visiter l’UE de La Fage et les caves de Roquefort ; c’était un très bon moment.
Enfin, je tiens à remercier mes parents, mon frère et toute ma famille ainsi que mes amis qui ont su m’encourager et me soutenir tout au long de ces cinq années d’études. Un grand merci à toutes les personnes qui ont participé de près ou de loin à ma formation.
Page | 4
Table des matières
Résumé ... 2
Remerciements ... 3
Table des matières ... 4
Table des illustrations ... 6
Liste des abréviations, symboles et unités ... 8
Glossaire ... 9
Introduction ... 10
1 Contexte et enjeux de l’étude ... 11
1.1 La filière caprine ... 11
1.1.1 Dans le monde et en Europe ... 11
1.1.2 En France ... 11
1.1.3 Deux grandes races laitières ... 12
1.2 Les programmes de sélection ... 13
1.2.1 Organisation des schémas de sélection ... 13
1.2.2 Les caractères en sélection ... 13
1.2.3 La longévité fonctionnelle, nouveau critère de sélection ... 14
1.3 L’immunité et la santé du jeune ... 14
1.3.1 Le colostrum ... 15
1.3.2 Le transfert d’immunité passive ... 19
1.3.3 Santé et bien-être du chevreau ... 22
1.4 Présentation de la problématique ... 26
2 Matériel et méthode ... 28
2.1 Matériel ... 28
2.1.1 Lieu d’expérimentation ... 28
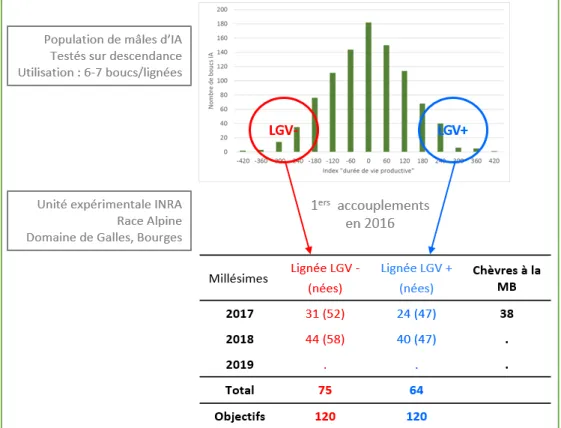
2.1.2 Les lignées divergentes ... 28
2.1.3 La conduite d’élevage ... 29
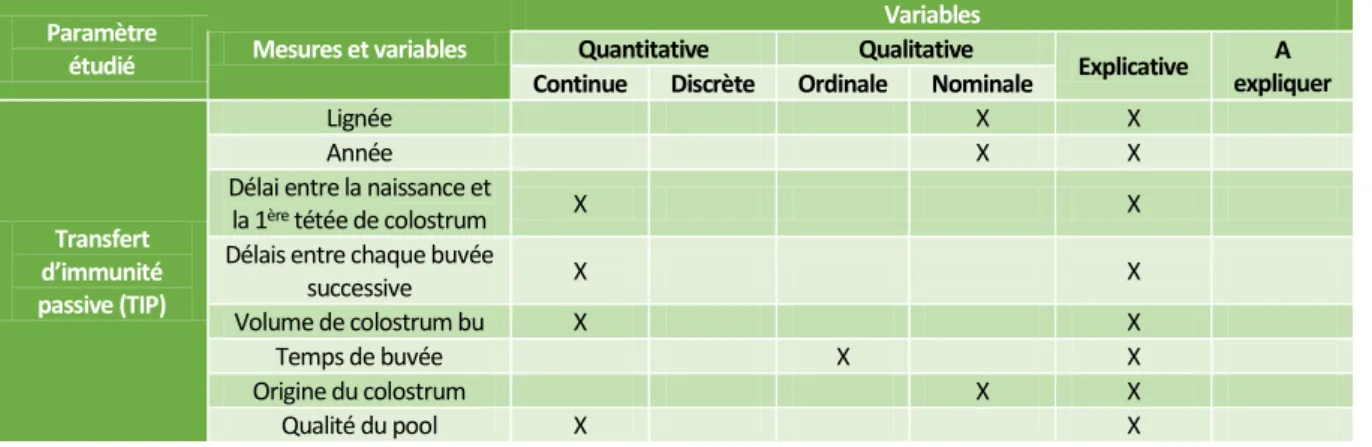
2.2 Mesures et variables ... 29
2.2.1 Mesures du transfert d’immunité passive et de la qualité du colostrum ... 29
2.2.2 Mesures de la santé des chevreaux ... 31
2.2.3 Mesures du comportement et du bien-être des chevreaux ... 33
2.2.4 Mesures de la croissance du chevreau ... 34
2.2.5 Informations supplémentaires ... 34
2.3 Traitements statistiques des données ... 34
2.3.1 Les variables analysées ... 34
2.3.2 Les analyses statistiques ... 35
Page | 5
3.1 Transfert d’immunité passive ... 39
3.1.1 Analyse descriptive ... 39
3.1.2 Analyse de variance ... 40
3.2 Qualité du colostrum ... 41
3.2.1 Analyse descriptive ... 41
3.2.2 Analyse de variance sur les deux traites ... 42
3.2.3 Analyse de variance sur la première traite ... 43
3.3 Santé du chevreau ... 44
3.3.1 Analyse descriptive ... 44
3.3.2 Analyse de variance ... 45
3.4 Comportement du chevreau face à l’Homme ... 47
3.4.1 Analyse descriptive ... 47 3.4.2 Analyse de variance ... 48 3.5 Croissance du chevreau ... 49 3.5.1 Analyse descriptive ... 49 3.5.2 Analyse de variance ... 49 4 Discussion et perspectives ... 51 4.1 Discussion ... 51
4.1.1 Protocole et mesures de l’étude ... 51
4.1.2 Transfert d’immunité passive ... 51
4.1.3 Qualité du colostrum ... 54
4.1.4 Santé du chevreau ... 55
4.1.5 Comportement des chevrettes face à l’Homme ... 56
4.1.6 Croissance du chevreau ... 57
4.1.7 Un effet de la lignée non présent ... 58
4.2 Perspectives ... 59
4.2.1 Transfert d’immunité passive ... 59
4.2.2 Qualité du colostrum ... 59
Conclusion ... 60
Liste des références bibliographiques... 61
Page | 6
Table des illustrations
Figure 1 : La chèvre Alpine ... 12
Figure 2 : La chèvre Saanen ... 12
Figure 3 : Schéma de sélection pour les caprins laitiers de race Alpine et Saanen ... 13
Figure 4 : Les 5 étapes du développement mammaire ... 17
Figure 5 : Mécanismes de transport des immunoglobulines maternelles vers le colostrum ... 17
Figure 6 : Représentation schématique du "trou immunitaire" chez le chevreau ... 20
Figure 7 : Dispositif expérimental pour la création des lignées divergentes ... 28
Figure 8 : Case collective de 15 places et 33 places ... 29
Figure 9 : Test de distance de fuite ... 33
Figure 10 : Balance utilisée pour la pesée des chevreaux et la prise de température ... 34
Figure 11 : Distribution de la variable concentration en IgG du sérum seule et par lignée ... 39
Figure 12 : Concentration en IgG du sérum en fonction de l'année de naissance et de la vigueur à la naissance ... 40
Figure 13 : Concentration en IgG du sérum en fonction du temps de buvée avec et sans le facteur heure de naissance ... 40
Figure 14 : Distribution de la variable concentration en IgG du colostrum seule et par lignée ... 41
Figure 15 : Concentration et masse d’IgG du colostrum en fonction du numéro de la traite ... 42
Figure 16 : Concentration en IgG du colostrum en fonction du volume produit ... 43
Figure 17 : Courbe de régression de la masse d'IgG du colostrum en fonction du volume produit ... 44
Figure 18 : Taux de mortalité sur les 3 premiers mois de vie en fonction du millésime ... 45
Figure 19 : Probabilité d'avoir au moins un écoulement nasal ou oculaire sur 5 semaines en fonction du millésime ... 46
Figure 20 : Probabilité d'avoir au moins une température anormale sur 5 semaines en fonction du TIP.. 46
Figure 21 : Score de propreté de l'arrière train et probabilité d'avoir au moins une diarrhée sur 5 semaines en fonction du TIP ... 47
Figure 22 : Score de distance de fuite en fonction du numéro du test ... 48
Figure 23 : Courbe de la croissance des chevreaux sur 3 mois de vie ... 49
Figure 24 : GMQ 0-60j et 0-90j en fonction du millésime ... 50
Figure 25 : Poids vif à 60 jours 0-90j en fonction du millésime ... 50
Figure 26 : Répartition géographique de la race Alpine ... 69
Figure 27 : Répartition géographique de la race Saanen ... 69
Figure 28 : Durée de vie productive moyenne chez les chèvres de race Alpine et Saanen en fonction de l'année de naissance ... 70
Figure 29 : Planning des mesures de la naissance à la mise bas pour un chevreau né au 15/01/2017 .. 73
Figure 30 : Distribution de la concentration en IgG du sérum (en log) ... 77
Figure 31 : Distribution de la concentration en IgG du colostrum (en log) ... 78
Figure 32 : Courbes-types de la croissance des chevrettes ... 79
Page | 7
Tableau 1 : Répartition des exploitations et du cheptel sur le territoire français en 2016 ... 11
Tableau 2 : Composants chimiques majeurs du colostrum et du lait de chèvre ... 15
Tableau 3 : Quantité d'immunoglobulines IgG, IgM et IgA dans le colostrum et le lait de chèvre de race Majorera ... 15
Tableau 4 : Evolution des concentrations en IgG, IgA et IgM de 0 à 90 jours après mise-bas sur la race Majorera ... 18
Tableau 5 : Définitions des scores de vigueur chez les agneaux ... 21
Tableau 6 : Indicateurs du bien-être chez la chèvre du projet AWIN ... 23
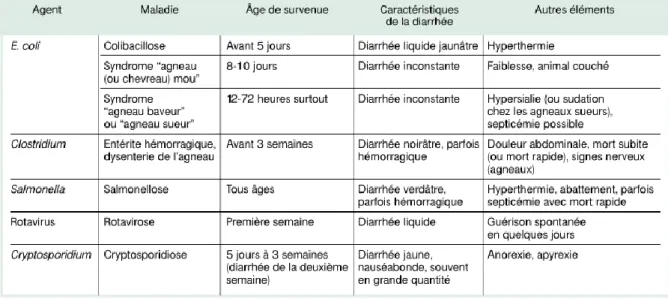
Tableau 7 : Eléments du diagnostic clinique et différentiel des diarrhées néonatales des petits ruminants ... 24
Tableau 8 : Effectifs des chevreaux avec un prélèvement sanguin ... 29
Tableau 9 : Notation du temps de buvée pour finir un biberon de 300ml ... 30
Tableau 10 : Notation de la vigueur du chevreau à 5min après la naissance ... 31
Tableau 11 : Effectifs des chevreaux étudiés par année et par lignée ... 31
Tableau 12 : Notation de la propreté de l'arrière train ... 31
Tableau 13 : Notation de l'écoulement nasal... 32
Tableau 14 : Notation de la distance de fuite ... 33
Tableau 15 : Notation du test de contact au menton ... 33
Tableau 16 : Types de variables en fonction de la mesure étudiée ... 34
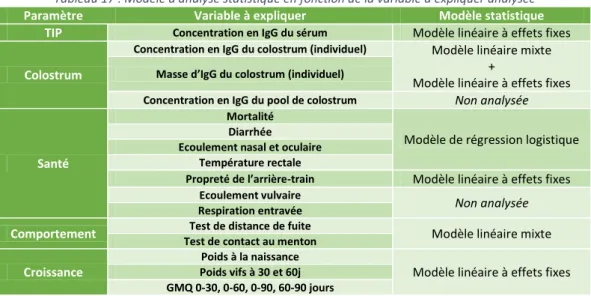
Tableau 17 : Modèle d'analyse statistique en fonction de la variable à expliquer analysée ... 36
Tableau 18 : Tableau des variables étudiées des facteurs associés ... 37
Tableau 19 : Classes pour le score de vigueur et pour le temps de buvée ... 38
Tableau 20 : Classes pour l'âge à la prise de sang et pour la taille de la portée ... 38
Tableau 21 : Classes pour la quantité de colostrum prélevée à la 1ère traite et le transfert d’immunité passive ... 38
Tableau 22 : Classes pour la température rectale ... 38
Tableau 23 : Concentration en IgG du sérum (en g/L) ... 39
Tableau 24 : Taux d'échec et de réussite du transfert d'immunité passive ... 39
Tableau 25 : Concentration en IgG du colostrum des traites 1 et 2 ... 41
Tableau 26 : Qualité du colostrum des traites 1 et 2 ... 42
Tableau 27 : Corrélations entre la concentration en IgG et la concentration en MG des deux traites ... 42
Tableau 28 : Taux d’animaux vivants ou morts au cours des 3 premiers mois de vie ... 44
Tableau 29a : Variabilités des mesures des relevés sanitaires... 44
Tableau 29b : Variabilités des mesures des relevés sanitaires ... 44
Tableau 30 : Corrélations entre la mortalité et les écoulements nasaux et oculaires ... 45
Tableau 31 : Résumé des effets significatifs des facteurs sur les différentes variables ... 45
Tableau 32 : Variabilité des données pour le test de distance de fuite par jour ... 47
Tableau 33 : Corrélations entre les différents tests de distance de fuite ... 48
Tableau 34 : Variabilité des données pour le test de contact au menton par jour ... 48
Tableau 35 : Corrélations entre les différents tests de contact au menton ... 48
Tableau 36 : GMQ et poids vifs (en g) ... 49
Tableau 37 : Moyennes pour 11 traits de production laitière et morphologie de la mamelle, héritabilité et corrélation génétique avec la longévité fonctionnelle ... 71
Page | 8
Liste des abréviations, symboles et unités
% Pourcent
%/h Pourcent par heure
°C Degré Celsius
ADN Acide Désoxyribonucléique
ARN Acide Ribonucléique
CAEV Arthrite-Encéphalite Virale Caprine
cm Centimètre
DAL Distributeur Automatique de Lait EGF Epidermal Growth Factor
g/dm3 Gramme par décimètre cube
g/hab/an Gramme par habitant et par an
g/j Gramme par jour
g/kg Gramme par kilogramme
g/L Gramme par litre
g/ml Gramme par millilitre GEB Groupe Economie du Bétail
GGT Gamma-Glutamyl-Transférase
GMQ Gain Moyen Quotidien
h Heure
h² Héritabilité
IA Insémination Animale
IDELE Institut de l’Elevage
IgA Immunoglobuline A
IGF Insuline-like Growth Factor
IgG Immunoglobuline G
IgM Immunoglobuline M
IMC Index Morphologique Caprin
INRA Institut National de Recherche Agronomique
j Jour
kg Kilogramme
L Litre
m Mètre
MG Matière Grasse
mg/dl Milligramme par décilitre mg/L Milligramme par litre mg/ml Milligramme par millilitre
ml Millilitre
NEC Note d’Etat Corporel QTL Quantitative Trait Loci
rg Corrélation génétique
s Seconde
SCC Comptage de cellules somatiques
TB Taux Butyreux
TEC Tonne Equivalent Carcasse TGF Transforming Growth Factor TIP Transfert d’Immunité Passive
TP Taux Protéique
UI/L Unité Internationale par litre
Page | 9
Glossaire
CAEV : Maladie virale de la chèvre. Elle entraîne une encéphalite chez les animaux de moins de 4 mois et, chez les animaux plus âgés, des arthrites, des mammites et plus rarement, des pneumonies chroniques.
Exocytose : Phénomène durant lequel les substances se trouvant dans le cytoplasme d’une cellule sont enveloppées par la membrane de cette dernière, pour ensuite être expulsées vers l’extérieur.
Héritabilité : Donnée statistique évaluant la part des facteurs génétiques dans la probabilité d’apparition d’un trait phénotypique donné, au sein d’une population donnée. Il s’agit de la part de la variance phénotypique relevant de la variance génotypique, pour un ensemble d’individus.
Immunoglobuline : Glycoprotéines présentant une fonction d’anticorps et jouant donc un rôle essentiel dans la défense de l’organisme face aux agressions extérieures.
Lignée divergente : Lignée d’animaux présentant des phénotypes extrêmes. QTL : Région de l’ADN associé à un caractère quantitatif.
Répétabilité : Fidélité de la mesure dans des conditions où les résultats sont obtenus dans des conditions de mesure identiques pendant un court intervalle de temps.
Reproductibilité : Fidélité de la mesure dans des conditions où les résultats sont obtenus dans des conditions de mesures différentes (lieu, équipement, opérateur…).
Transcytose : Migration, dans des cellules polarisées, entre les domaines basolatéraux et apicaux via des vésicules.
Transfert d’immunité passive : Transfert des immunoglobulines produites par la mère et présentes dans sa circulation sanguine vers sa progéniture. L’immunité est acquise de façon passive lorsque le transfert des immunoglobulines se réalise par ingestion du colostrum de la mère.
Page | 10
Introduction
Depuis le début des années 2000, une évolution des objectifs de sélection génétique des caprins laitiers s’est mise en place. Dans les années 70, cette sélection était principalement basée sur des critères de production laitière et de qualité du lait, notamment sur les taux butyreux et protéiques. Depuis quelques années, l’apparition de nouveaux critères tels que la morphologie de la mamelle et la résistance aux mammites entrent en jeu. Le contexte scientifique actuel montre les impacts de la santé et du bien-être animal sur la rentabilité des élevages (HEMSWORTH et al., 1995; HAENLEIN, 2002; ROCHE
et al., 2009). Ainsi, la longévité des animaux est devenue un enjeu majeur pour la filière, tant d’un
point de vue économique que sociétal.
La longévité fonctionnelle d’un animal reflète ses aptitudes à surmonter des challenges alimentaires et sanitaires (PALHIERE et al., 2018). Ce trait permet de quantifier la durée de vie productive
d’un animal et donc d’accroître la rentabilité d’un élevage. Or, au cours des 20 dernières années, il s’avère que la durée de vie productive des chèvres a diminué de 320 jours en moyenne. La longévité fonctionnelle d’un animal est principalement liée à des mécanismes adaptatifs qui opèrent à différents niveaux d’organisation physiologique, notamment dans le jeune âge. Il faut noter que le début de carrière des animaux d’élevage est une période très importante du fait qu’elle soit non productive. La survie des animaux à ce stade est donc essentielle pour éviter une perte non rentabilisée par la production laitière.
La santé et le bien-être des chevreaux sont les premiers déterminants de la carrière de l’animal. Ces paramètres sont en grande partie sous l’influence d’une ingestion précoce mais également efficace du colostrum. Le colostrum est le premier lait après la mise-bas. Il est une source importante de nutriments, de facteurs de croissance et d’immunoglobulines qui permettent le bon développement du nouveau-né, le transfert d’immunité passive et ainsi la protection contre les agressions extérieures. Il est donc nécessaire d’évaluer le transfert d’immunité passive, la qualité du colostrum et leur rôle sur la santé et le bien-être des chevreaux ainsi que sur leur croissance, sujets très peu documentés chez les caprins en comparaison des bovins (WEAVER et al., 2000; CAJA et al., 2006; KEHOE et al., 2011).
Dans cette continuité, le projet ACTIVEGOAT, démarré en 2016 et reposant sur une sélection d’animaux divergents sur la longévité fonctionnelle, a pour but d’étudier le rôle de la génétique, notamment dans le jeune âge, sur ce critère. A cet égard, le projet a pour objectif l’évaluation du transfert d’immunité passive, de la qualité du colostrum, de la santé et du bien-être des chevreaux ainsi que de leur croissance par le développement de mesures et l’estimation de différents facteurs sur ces paramètres.
Après avoir posé le contexte dans lequel s’inscrit l’étude et justifié les mesures utilisées, une seconde partie présentera plus en détail le protocole mis en place. Les résultats sur les différentes estimations des facteurs de variation (hors génétique), de l’effet du transfert d’immunité passive et de la lignée seront également exposés. Enfin, le protocole et les résultats de l’étude seront ensuite discutés et des pistes d’amélioration seront abordées.
Page | 11
1 Contexte et enjeux de l’étude
1.1 La filière caprine
1.1.1 Dans le monde et en Europe
D’après la FAO, le cheptel caprins mondial était de 1,11 milliards de têtes en 2014 et produisait 18,34 millions de tonnes de lait et 5,524 millions de tonnes de viande (IDELE, CNE, 2017).
En 2014, l’Asie domine le reste du monde en termes de taille de cheptel avec 581 millions de têtes, suivi par l’Afrique, l’Amérique puis l’Europe. La production laitière est également dominée par l’Asie avec 11,31 millions de tonnes de lait, l’Europe étant placé à la 3ème place (
IDELE, CNE, 2017) derrière l’Afrique.
L’Union Européenne comptait, en 2016, 12,8 millions de caprins (EUROSTAT, 2018) pour une
production de 2,25 millions de tonnes de lait et 55 000 TEC (IDELE, CNE, 2017).
En 2017, la France se place 4ème après la Grèce, l’Espagne et la Roumanie en terme de cheptel (A
GRESTE, 2018) mais prend la 1ère place en production laitière (ANICAP, INTERBEV, 2017) avec ses 603 millions de
litres produit en 2016. Cette différence de production et de cheptel peut notamment s’expliquer par le fait que la France possède des schémas de sélection efficaces pour ses races caprines laitières (IDELE, CNE, 2017).
1.1.2 En France
La France compte, en 2016, 4 920 exploitations (>10 chèvres avec activité laitière) (IDELE, CNE,
2017) contre 123 000 exploitations en 1979 (AGRESTE, 2013). 1,213 millions de caprins sont présents
sur le sol français en 2017 dont 831 milliers de chèvres et 285 milliers de chevrettes (AGRESTE, 2018)
soit 1% de plus qu’en 1979 mais 8% de moins qu’en 2012 (AGRESTE, 2013). La France possède ainsi 9,5% du cheptel européen (ANICAP, INTERBEV, 2017).
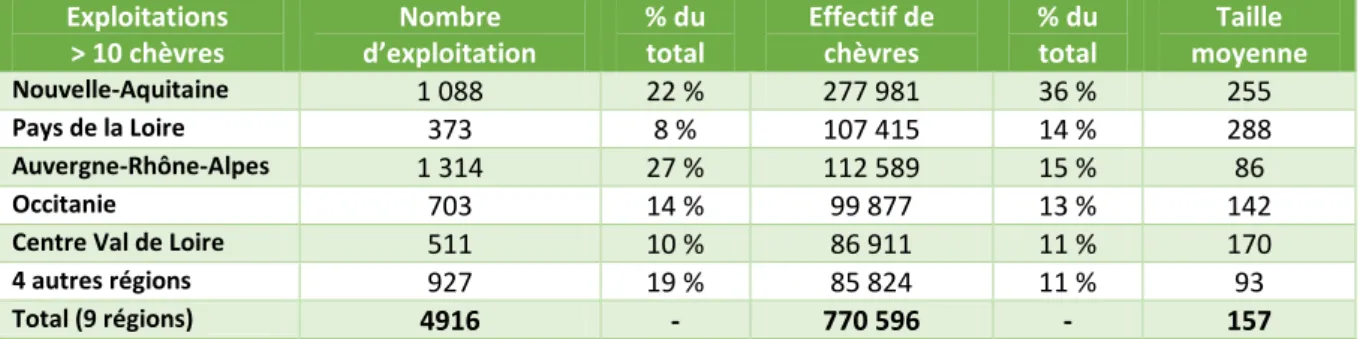
60% des élevages possèdent plus de 200 chèvres et 26% en comptent entre 100 et 199 (IDELE, CNE, 2017). Le cheptel français est regroupé dans 5 régions : Nouvelle-Aquitaine, Pays de la Loire, Auvergne-Rhônes-Alpes, Occitanie et Centre Val de Loire (cf. tab. n°1)
Tableau 1 : Répartition des exploitations et du cheptel sur le territoire français en 2016 (IDELE, CNE, 2017)
Exploitations > 10 chèvres Nombre d’exploitation % du total Effectif de chèvres % du total Taille moyenne Nouvelle-Aquitaine 1 088 22 % 277 981 36 % 255 Pays de la Loire 373 8 % 107 415 14 % 288 Auvergne-Rhône-Alpes 1 314 27 % 112 589 15 % 86 Occitanie 703 14 % 99 877 13 % 142
Centre Val de Loire 511 10 % 86 911 11 % 170
4 autres régions 927 19 % 85 824 11 % 93
Total (9 régions) 4916 - 770 596 - 157
En élevage caprin, les jeunes éleveurs (de moins de 35 ans) ne comptent que pour 15% des exploitations tandis que les éleveurs de 35 à 54 ans sont représentés à 66% et à 19% pour les 55 ans et plus (IDELE, CNE, 2017).
La production laitière française était, en 2016, de 603 millions de litres contre 476 millions en 2000, soit une hausse de 21% sur 16 ans. Cette même année, 100 724 tonnes de fromages de chèvre ont été produites par les industries laitières et 18 à 20 000 tonnes (estimations GEB) ont été produites en ferme (IDELE, CNE, 2017). Le marché du fromage de chèvre se porte bien, celui-ci ayant un taux de
pénétration de 85,1% pour 57 030 tonnes achetées en 2017 (IDELE, CNE, 2017). La France est ainsi le 1er consommateur de fromages de chèvre en Europe (ANICAP,
INTERBEV, 2017) avec 700 g/hab/an de produits issus du lait de chèvre. En revanche, il faut noter que le fromage de chèvre ne représente que 6,5% des volumes de fromages achetés par les ménages, le fromage au lait de vache comptant pour
Page | 12 89,8% des volumes (FRANCEAGRIMER, 2017). En 2016, 465 millions de litres ont été collectés au niveau
national et 99 millions de litres ont été importés pour une exportation de fromages industriels d’environ 20 000 tonnes (IDELE, CNE, 2017). La demande en produits à base de lait de chèvre connaît une croissance régulière, notamment en Europe et aux Etats-Unis (ANICAP, INTERBEV, 2017).
1.1.3 Deux grandes races laitières
En France, les deux principales races caprines sont la chèvre Alpine (cf. fig. n°1) et la chèvre Saanen (cf. fig. n°2). D’après le contrôle laitier de 2016, ces deux races comptent, à elles seules, pour 97% du cheptel caprin français (IDELE, 2017).
La chèvre Alpine est une race originaire du massif alpin (CAPGENES, 2017) et aujourd’hui présente
sur la quasi-totalité du territoire français (cf. Annexe A, fig. n°26). Avec 450 000 chèvres, dont un peu plus de 150 000 soumises au contrôle laitier 2016, il s’agit de la race la plus répandue en France (IDELE, 2017). Cette race est sélectionnée depuis les années 70, dans le cadre d’un schéma collectif d’amélioration des qualités laitières (CAPGENES, 2017).
Caractéristiques de la chèvre Alpine (CAPGENES, 2017; IDELE, 2017): - Poids : 50 à 70 kg
- Durée lactation : 298 j - Lait : 929 kg
- TP : 33,4 g/kg - TB : 37,8 g/kg
La Saanen est originaire de Suisse et est la race laitière la plus répandue dans le monde (CAPGENES, 2017). 350 000 chèvres, dont environ 105 000 chèvres contrôlées, sont exploitées dans le Sud-Est, le Centre et l’Ouest de l’Hexagone (cf. Annexe A, fig. n°27) (IDELE, 2017). Un schéma de sélection a
démarré en France dans les années 70, portant sur la production laitière (quantité de lait et taux) ainsi que la morphologie (CAPGENES, 2017).
Caractéristiques de la chèvre Saanen (CAPGENES, 2017; IDELE, 2017) : - Poids : 50 à 90 kg
- Durée lactation : 311 j - Lait : 985 kg
- TP : 32,2 g/kg - TB : 35,9 g/kg
D’autres races à petits effectifs sont également présentes en France dont : la Pyrénéenne, l’Angora, la Corse, la Rove, la chèvre Poitevine, la chèvre des fossés ainsi que la chèvre de Lorraine.
Page | 13
1.2 Les programmes de sélection
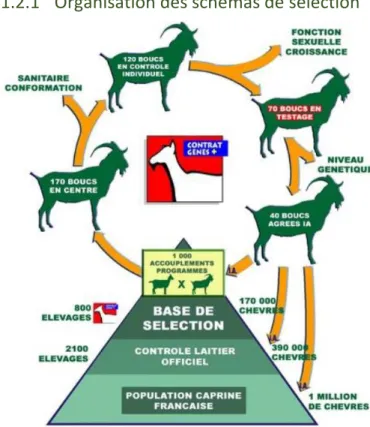
1.2.1 Organisation des schémas de sélection
A l’heure actuelle, un seul organisme dirige les schémas de sélection pour les races laitières Alpine et Saanen (cf. fig. n°3). Il s’agit de l’unique organisme de sélection en caprins ainsi que le seul centre national de production de semence de boucs (CAPGENES, 2017).
Pour ce schéma de sélection, Capgenes regroupe, en 2016, environ 600 éleveurs sélectionneurs et possède une base de sélection comportant 170 000 chèvres sur les 390 000 inscrites au contrôle laitier officiel (CAPGENES, 2017).
L’INRA, l’IDELE ainsi que le Ministère de l’Agriculture ont validé ce schéma de sélection. Une fois cette validation réalisée, les animaux peuvent être répertoriés dans un fichier national racial. Grâce aux données récoltées, l’INRA peut ensuite calculer les valeurs génétiques des reproducteurs et ainsi les indexer sur différents paramètres.
L’efficacité du schéma de sélection se base sur différents facteurs dont l’insémination animale. En 2016, pas moins de 70 000 chèvres ont été inséminées (CAPGENES, 2017). Les boucs utilisés pour
l’amélioration du schéma de sélection doivent être testés dans des stations de contrôle à l’âge d’un mois. Ces jeunes boucs sont issus de schémas d’accouplements entre les meilleures femelles de la base de sélection et les meilleurs mâles reproducteurs. Via les performances des filles inscrites au contrôle laitier et du pointage sur la production laitière et la morphologie mammaire, il est possible d’indexer les mâles. Les mâles sont ainsi indexés sur descendance avec un minimum de 30 filles contrôlées par mâle. Sur les 70 mâles testés tous les ans, seulement une quarantaine sont choisis pour la diffusion de la semence (CAPGENES, 2017).
1.2.2 Les caractères en sélection
Depuis les années 70, les principaux critères de sélection sont la production laitière, les quantités de matières grasses et protéiques produite par chèvre et par lactation, mais aussi les taux protéiques et butyreux du lait. Une sélection sur des caractères morphologiques a vu le jour depuis les années 2000 afin d’améliorer l’aptitude des chèvres à la traite. Depuis 2005, un index de synthèse morphologique (IMC) a été mis en place et inclut 4 postes descriptifs de la morphologie mammaire : profil de la mamelle, hauteur du plancher, qualité de l’attache-arrière et forme de l’arrière-pis (CLEMENT
et al., 2006).
Cela fait maintenant quelques années que des caractères autres que la production laitière ont été intégrés dans les schémas de sélection des caprins laitiers. En 2010, l’INRA a mis en place une évaluation génétique sur le comptage cellulaire des races Alpine et Saanen comme critère de sélection pour la résistance aux mammites (CLEMENT et al., 2016).
Figure 3 : Schéma de sélection pour les caprins laitiers de race Alpine et Saanen (Capgenes, 2017)
Page | 14 En 2016, l’INRA démarre des recherches pour sélectionner les caprins sur leur longévité grâce à une base de presque 1 138 000 chèvres Saanen et Alpine (PALHIERE et al., 2018). Le projet CASDAR
RUSTIC a pour but l’indexation des ovins et caprins sur la longévité fonctionnelle. Aujourd’hui à l’état d’expérimentation, ce caractère pourrait devenir un point important pour la sélection.
1.2.3 La longévité fonctionnelle, nouveau critère de sélection
La longévité est un trait permettant de quantifier la durée de vie productive d’un animal de rente et d’accroître la rentabilité d’un élevage. Dans les élevages laitiers, la longévité peut se traduire par :
La longévité réelle : durée de vie productive de l’animal qui comprend toutes les causes de réforme y compris la productivité.
La longévité fonctionnelle : durée de vie productive de l’animal qui comprend toutes les causes de réforme sauf la productivité.
La longévité fonctionnelle est donc ainsi corrigée pour le niveau de production, qui est l’une des premières causes de réforme (SASAKI, 2013). Elle reflète l’aptitude de l’animal à surmonter des challenges alimentaires ou sanitaires. Au cours des 20 dernières années, la durée de vie productive a diminué de 320 jours en moyenne pour les races Alpine et Saanen (cf. Annexe B, fig. n°28). Cette diminution est notamment liée à des taux de renouvellement plus importants allant de 20% dans les années 90 à 30% aujourd’hui. Chez la chèvre Alpine, il a été montré que la longévité fonctionnelle est corrélée négativement avec le score de cellule somatique (rg = -0,35 ± 0,04) et positivement avec la hauteur de la mamelle (rg = 0,26 ± 0,04) (cf. Annexe C, tab. n°37) (PALHIERE et al., 2018). Ces résultats
coïncident avec d’autres études réalisées chez la vache et chez la chèvre (SASAKI, 2013; CASTAÑEDA -BUSTOS et al., 2017) indiquant qu’une mamelle en bonne santé et une bonne morphologie contribuent
à une longévité fonctionnelle plus élevée. L’héritabilité de la durée de vie productive reste cependant faible avec h²=0,08 (±0,02) chez la chèvre Alpine. Chez la chèvre Saanen, un QTL très significatif (p=1,94e-09) pour la longévité fonctionnelle a été trouvé et pourrait offrir la possibilité d’améliorer la sélection par la génomique et de mieux comprendre les gènes et mécanismes sous-jacents à ce caractère complexe (PALHIERE et al., 2018).
La longévité d’un animal est due à des mécanismes adaptatifs opérant à différents niveaux d’organisation physiologique. Ils sont en partie sous le contrôle des conditions d’élevage et de nutrition, notamment dans le jeune âge (BATESON, GLUCKMAN, 2011). La dégradation de la durée de vie productive a eu des conséquences négatives sur la santé, la reproduction ainsi que la longévité des animaux. Par conséquent, cela a impacté l’efficacité de production et donc la rentabilité des élevages (BEAUMONT et al., 2002). Le début de carrière des animaux d’élevage est une partie importante puisqu’il
s’agit d’une période non productive. Ainsi, il est essentiel de pouvoir jouer sur la survie de l’animal et éviter la mortalité pour ne pas engendrer de perte économique alors que l’animal n’a pas encore commencé sa carrière productive. En moyenne, le taux de mortalité périnatale est de 14% et varie généralement entre 8 et 17%. En système extensif, le taux de mortalité peut atteindre les 60% (FERNANDEZ, 2014;O’BRIEN, SHERMAN, 1993). De plus, 50% de la mortalité entre la naissance et le sevrage
se déroule dans les 24 premières heures de vie de l’animal (CHEMINEAU et al., 1985; DWYER et al., 2016).
1.3 L’immunité et la santé du jeune
La santé et le bien-être des chevreaux sont les premiers déterminants de la carrière de l’animal et sont sous l’influence d’une ingestion précoce et efficace de colostrum. L’immunité acquise de façon passive par cette ingestion de colostrum est un élément important pour la survie du jeune. C’est pourquoi, dans un premier temps, il est important d’étudier ce qu’est le colostrum et la colostrogénèse. Une seconde partie se focalisera sur le transfert d’immunité passive, avant d’enchainer sur la santé et le bien-être des chevreaux. Les méthodes d’évaluation de ces différents paramètres seront présentées dans chacune des parties.
Page | 15
1.3.1 Le colostrum
1.3.1.1 Définition et composition
D’un point de vue biologique, le colostrum est le premier lait après la mise-bas. Il possède un aspect jaunâtre, plus dense, plus épais et plus visqueux que le lait (ABDOU et al., 2012). Il s’agit d’un
mélange de constituants du sérum sanguin et de sécrétions lactées s’accumulant dans la glande mammaire lors de la période sèche (JACQUES, 2012). Le colostrum joue un rôle dans la protection contre les agressions extérieures et dans le développement des nouveau-nés par l’apport de nutriments et d’immunoglobulines (BOUDRY, THEWIS, 2009). Chez les nouveau-nés des espèces ayant un placenta
épithéliochorial (ou syndesmochoriale), pour les caprins et ovins, le transfert d’immunoglobulines s’effectue par l’ingestion du colostrum car celles-ci ne peuvent traverser la barrière placentaire (NOWAK, POINDRON, 2006).
Le colostrum possède une composition chimique différente du lait (cf.tab. n°2). Il est une source importante de nutriments protéiques, glucidiques, lipidiques, vitaminiques et minéraux (ABDOU et al.,
2012). Le colostrum présente des concentrations plus élevées en protéines, lipides et minéraux mais plus faibles en lactose.
Tableau 2 : Composants chimiques majeurs du colostrum et du lait de chèvre (YANG et al., 2009; SANCHEZ-MACIAS et al., 2014)
Composants Colostrum Lait
Densité (g/mL) 1,048 1,028 Matière sèche (%) 21,32 ± 0,07 16,20 ± 0,07 Protéines (%) 10,24 ± 2,15 5,91 ± 2,17 Lipides (%) 7,73 ± 0,44 4,30 ± 0,43 Lactose (%) 1,93 ± 0,12 3,99 ± 0,07 Minéraux (%) 1,57 ± 0,06 0,89 ± 0,08
Il contient des concentrations très élevées en calcium, phosphore et magnésium ainsi que des oligo-éléments essentiels tels que du cuivre, du magnésium, du molybdène, du nickel, du sélénium, de l’étain, du chrome, de l’aluminium, du lithium et du cobalt qui sont des éléments importants dans le système immunitaire (YANG et al., 2009). La quantité importante de protéines apportée par le
colostrum permet au jeune animal de synthétiser ses propres protéines. La stimulation du métabolisme des protéines après la mise-bas requiert en effet une grande quantité d’acides aminés (QUIGLEY, DREWRY, 1998).
De plus, le colostrum apporte des effecteurs immunitaires comme les immunoglobulines et les leucocytes mais également d’autres composés antimicrobiens : cytokines, lysozymes, lactoferrines et lactoperoxydases. La lactoferrine est une glycoprotéine qui joue un rôle dans l’absorption du fer dans l’intestin et dans l’activation des phagocytes et de la réponse immunitaire. La lactoperoxydase est, quant à elle, une enzyme antibactérienne qui inhibe le métabolisme des bactéries par oxydation. Les lysozymes sont également des enzymes bactériennes intervenant sur la paroi bactérienne en hydrolysant les peptidoglycanes qu’elle contient. Les immunoglobulines, IgG, IgA et IgM (cf. tab. n°3), participent au transfert d’immunité passive chez les jeunes animaux. Ces immunoglobulines
permettent de protéger le nouveau-né contre les infections (PAKKANEN, AALTO, 1997).
Tableau 3 : Quantité d'immunoglobulines IgG, IgM et IgA dans le colostrum et le lait de chèvre de race Majorera (SANCHEZ-MACIAS et al., 2014)
Concentration en
Immunoglobulines Colostrum Lait
IgG (mg/ml) 32,99 0,88
IgA (mg/ml) 0,86 0,07
IgM (mg/ml) 3,84 0,2
Le colostrum possède également des facteurs de croissance et des hormones, notamment les
Insulin-like Growth Factor (IGF) I et II, l’insuline, les Beta Transforming Growth Factors (TGF) β1 et β2
et les Epidermal Growth Factor (EGF). Les IGF sont des médiateurs de la croissance, du développement et de la différenciation cellulaire. Ils stimulent l’absorption du glucose, ainsi que la synthèse de
Page | 16 glycogène, de protéines, d’ARN, d’ADN et de lipides. Ils agissent également comme des hormones endocrines via le sang. Les TGF vont, quant à eux, permettre une stimulation de la prolifération de certaines cellules dans les tissus conjonctifs mais agissent également comme inhibiteurs d’autres cellules comme les lymphocytes et les cellules épithéliales. Les TGF sont aussi utiles pour l’embryogénèse, la réparation des tissus, la formation des os et du cartilage ainsi que le contrôle du système immunitaire. Cependant, la fonction des TGF dans le colostrum et le lait reste inconnue. Certaines hypothèses ont été émises proposant que les TGF puissent être des médiateurs de l’immunité de la muqueuse et/ou de la différentiation des cellules épithéliales intestinales chez le nouveau-né. Les EGF jouent un rôle dans le développement de l’épiderme, de la glande mammaire et de l’intestin et agissent comme des facteurs angiogéniques, c’est-à-dire permettant la croissance de nouveaux vaisseaux sanguins en utilisant des vaisseaux préexistants (PAKKANEN, AALTO, 1997).
Cependant, le colostrum peut également être un vecteur de maladies. En effet, chez les caprins, il est notamment vecteur de l’Arthrite-Encéphalite Virale Caprine, communément appelée le CAEV. Ce virus fait partie de la famille des lentivirus et s’intègre directement dans le génome des cellules du système immunitaire (monocytes et macrophages). L’évolution de cette maladie est lente et irréversible car le système immunitaire est incapable de l’éliminer. Les cellules cibles se retrouvent ensuite dans le sang et dans différents organes : gaine synoviale des articulations, poumons, glande mammaire et cerveau. Suite à l’infection et réplication du virus, il y a réaction inflammatoire dans les tissus infectés qui entraine leur épaississement. Ainsi, les différents signes cliniques observables sont : une atteinte des articulations (arthrite chronique), une réduction de la tolérance à l’exercice, une détresse respiratoire (pneumonie) et une induration de la glande mammaire (mammite). Des symptômes neurologiques peuvent également être observés chez les chevreaux. Outre le colostrum et le lait qui sont les principales voies de transmission, le CAEV peut également se propager par les sécrétions nasales et le sang (VANDIEST, 2004). En revanche, seulement 30% des animaux infectés présentent des signes cliniques. En élevage, ce virus entraîne donc des diminutions des performances de reproduction et de production (détérioration qualitative mais aussi quantitative du lait) en plus d’une dégradation de l’état sanitaire de l’animal (CREMOUX, 2012). Des moyens de prévention ont été
mis en place chez le jeune et l’adulte (GDSCENTRE, 2017):
Pour les jeunes animaux :
Séparation de la mère immédiatement après mise-bas : ne pas faire lécher, téter, favoriser le bouchonnage et séchage, désinfection du cordon…
Thermisation du colostrum et du lait : inactivation du virus lorsque le colostrum/lait est chauffé à 56°C (± 2°C) pendant 1h, avec agitateur et thermostat.
Eviter les traumatismes des membres : ne pas attraper/porter les animaux par les pattes. Pour les adultes :
Elevage en processus d’éradication ou qualifié d’indemne : introduction exclusive d’animaux qualifiés d’indemnes CAEV. Pour les autres élevages : faire attention aux symptômes cliniques lors des introductions.
Respecter les mesures d’hygiènes au niveau du matériel sanitaire et de la traite : faire passer les non contaminés avant les autres, éviter les entrées d’air dans les manchons de traite…
Limiter l’infection des nouveaux animaux et l’apparition des signes cliniques chez les animaux infectés : entretien de la machine à traire, changement des manchons une fois par an au minimum, lavage complet matin et soir, bonnes pratiques de traite, utilisation de matériel d’injection à usage unique, limiter les fatigues articulaires (logement des animaux).
1.3.1.2 La colostrogénèse
Page | 17
Figure 4 : Les 5 étapes du développement mammaire
Contrairement au lait qui est produit par sécrétion de la mamelle, le colostrum est quant à lui produit à la fois par transsudation et par sécrétion mammaire (BOURNE, CURTIS, 1973).
La colostrogénèse est le transfert prepartum des immunoglobulines de la circulation sanguine vers les sécrétions mammaires. Chez les espèces domestiques, ce transfert commence quelques semaines avant la mise-bas et s’arrête immédiatement au moment de celle-ci (BARRINGTON et al., 2001).
Le sérum maternel possède de nombreuses protéines et d’immunoglobulines de classes différentes. Seules les IgG sont transférées sélectivement en quantité importantes vers le colostrum. Les IgA et IgM sont également présentes dans le colostrum en moindre quantité (LARSON et al., 1980).
Ce transport sélectif des IgG vers le colostrum comprend deux fonctions distinctes. Tout d’abord, des récepteurs spécifiques aux IgG, les « Fc Receptors of the neonate » (BAUMRUCKER, BRUCKMAIER,
2014), doivent être présent sur la membrane basale des lactocytes. Une fois les IgG fixées sur les récepteurs Fc, la transcytose peut se dérouler. Les cellules épithéliales mammaires vont internaliser les IgG dans des vacuoles de transport puis les délivrer dans la lumière des acini mammaires lorsque le pH est acide (BARRINGTON et al., 2001). Les IgA et IgM sont synthétisées localement dans la glande
mammaire par les plasmocytes du parenchyme mammaire (LARSON et al., 1980) (cf. fig. n°5, gauche).
Ces immunoglobulines sont reconnues par un récepteur des immunoglobulines polymériques nommé « pIgR ». Une fois les IgA et IgM reconnues, les complexes IgA/IgM et pIgR sont transportés dans des vésicules à travers le cytoplasme vers la lumière par transcytose. Lorsque le complexe atteint la surface luminale de la cellule épithéliale, il subit un clivage protéolytique, libérant ainsi les IgA de la partie extracellulaire du récepteur ou composante sécrétoire (SALMON, 2011) (cf. fig. n°5, droite).
Figure 5 : Mécanismes de transport des immunoglobulines maternelles vers le colostrum (SALMON, 2011)
La régulation de la colostrogénèse se réalise grâce à des contrôles endocriniens puisque différentes hormones interviennent dans cette régulation. En effet, la colostrogénèse commencerait lorsque la concentration en progestérones chute et que celle en œstrogènes augmente, cette dernière initie le transfert des IgG. Les hormones nécessaires à la lactogénèse, telle que la prolactine, jouent un rôle important dans l’arrêt de la colostrogénèse. Il a été montré que la prolactine provoque l’arrêt de la colostrogénèse en augmentant l’activité lactogénique, et en diminuant la concentration des sécrétions d’IgG et l’activité des récepteurs des IgG (BAUMRUCKER, BRUCKMAIER, 2014).
1.3.1.3 Facteurs de variation
La qualité du colostrum est caractérisée par sa concentration et/ou sa masse en immunoglobulines. Plusieurs facteurs peuvent influencer cette teneur en immunoglobulines dans le colostrum.
Page | 18 Tout d’abord, il faut noter que la concentration en immunoglobulines du colostrum diminue rapidement après parturition (cf. tab. n°4).
Tableau 4 : Evolution des concentrations en IgG, IgA et IgM de 0 à 90 jours après mise-bas sur la race Majorera (SANCHEZ-MACIAS et al., 2014)
Concentration en immunoglobulines
Jours après mise-bas
0 1 2 3 4 5 15 30 60 90
IgG (mg/ml) 32,99 20,13 8,23 6,05 2,16 1,87 1,02 1,09 0,80 0,88
IgA (mg/ml) 0,86 0,40 0,24 0,19 0,17 0,11 0,11 0,12 0,10 0,07
IgM (mg/ml) 3,84 1,20 0,70 0,59 0,38 0,38 0,22 0,16 0,18 0,20
Cela s’explique par des facteurs affectant la colostrogénèse chez les petits ruminants. Grâce à l’inactivation des récepteurs spécifiques aux IgG sur les cellules épithéliales alvéolaires, le transfert des immunoglobulines est alors inhibé par une augmentation de la prolactine durant la lactogénèse.
Une étude a également montré que la taille de la portée a un effet sur la qualité immunitaire du colostrum. En effet, il a été observé que le colostrum contenait 20,2 mg/ml d’IgG de plus chez les chèvres ayant 2 chevreaux que chez celles n’en ayant qu’un (CSAPO et al., 1994).
D’après G. CAJA et al, la période de tarissement impacte la concentration en immunoglobulines. Ils ont en effet montré que les chèvres sans tarissement possédaient une concentration en IgG de 5,6 mg/ml contre 42,4 mg/ml pour les chèvres ayant 56 jours de tarissement avant mise-bas. Ce résultat peut s’expliquer par le fait que la traite continue affecte la colostrogénèse. En effet, à l’approche de la mise-bas, il y a un transfert sélectif massif des IgG du sérum vers le colostrum. Ce transfert d’IgG est localement contrôlé par les cellules épithéliales mammaires. Lorsque la sécrétion lactée est maintenue, ce transfert diminue. Une traite continue maintient ainsi l’activité des cellules épithéliales mammaires et empêche l’accumulation des IgG dans le colostrum (CAJA et al., 2006).
En 2017, une étude a été menée pour connaitre les facteurs influençant la qualité du colostrum chez les vaches de race Holstein et leurs liens avec la concentration postpartum du calcium dans le sang et le colostrum. Il en ressort que le facteur influençant le plus la concentration en immunoglobulines du colostrum est d’origine génétique puisqu’il s’agit du bagage génétique du père, suivi par la quantité de colostrum produit et la durée du tarissement. Ainsi, une sélection des taureaux, pour lesquels leurs filles produisent un colostrum riche en immunoglobulines, pourrait améliorer la qualité du colostrum du troupeau (KARL, STAUFENBIEL, 2017).
1.3.1.4 Méthodes d’évaluation
Comme il a été vu précédemment, le colostrum possède de nombreux facteurs immunitaires. En revanche, le dosage des immunoglobulines G est généralement le seul indicateur utilisé pour l’évaluation de la qualité immunitaire du colostrum. Il existe 2 types de méthodes pour doser les IgG : les méthodes directes et les méthodes indirectes.
Méthodes directes :
Il s’agit d’un dosage quantitatif des IgG, basé sur le principe de réaction des anticorps face aux antigènes. Deux techniques sont utilisées, l’immunodiffusion radiale et le test ELISA (JACQUES, 2012).
La technique d’immunodiffusion radiale est considérée comme la méthode de choix pour quantifier les IgG. La concentration en immunoglobulines peut être déterminée dans le lait, le lactosérum, le colostrum ainsi que dans le sérum et le plasma des jeunes animaux de moins de 30 jours. L’échantillon est déposé sur une plaque constituée d’un gel d’agar et contenant un antisérum dirigé contre les IgG. Pendant l’étape de diffusion, les IgG réagissent avec l’antisérum et forment un disque de précipitation autour du puits de dépôt. La surface de chaque précipité est proportionnelle à la concentration en IgG de l’échantillon. A partir des diamètres mesurés pour les points de gamme dont la concentration en IgG est connue, une droite de régression linéaire est établie et utilisée pour
Page | 19 calculer la concentration en IgG de chaque échantillon (JACQUES, 2012). Il est considéré qu’un colostrum
de chèvre est de mauvaise qualité lorsque celui-ci contient moins de 40 g/L d’IgG (EHRHARDT et al.,
2014).
Pour le test ELISA, l’échantillon de colostrum doit être dilué dans une solution saline puis déposé sur une plaque de microtitration possédant des puits qui contiennent des anticorps dirigés contre les immunoglobulines à tester. Il y a ensuite incubation à 37°C pendant une heure et rinçage des puits. Des anticorps marqués par une enzyme sont ensuite rajoutés aux complexes immunoglobulines-anticorps anti-immunoglobulines et dirigés contre les immunoglobulines à tester. Un substrat chromogène de l’enzyme est ajouté et l’absorbance des échantillons est évaluée par spectrophotométrie. L’intensité de la coloration obtenue est proportionnelle à la concentration en immunoglobulines de l’échantillon (JACQUES, 2012).
Méthodes indirectes :
Les méthodes indirectes comprennent également 2 techniques. Une mesure de la densité du colostrum grâce à un colostromètre et une mesure de la teneur totale en protéines via un réfractomètre.
Le colostromètre estime la richesse en immunoglobulines par la mesure de la densité du colostrum. La densité du colostrum est corrélée positivement avec la concentration en IgG mais peut être influencée par certains facteurs comme la température (JACQUES, 2012). Le seuil d’un colostrum de qualité moyenne chez les caprins est de 75g/L d’immunoglobulines (IDELE, 2014).
Le réfractomètre permet une évaluation de la teneur totale en protéines du colostrum (corrélée positivement avec la teneur en IgG) par mesure de son indice de réfraction (JACQUES, 2012). Chez les caprins, la limite entre les « bon » et « mauvais » colostrums est fixée à 24 % BRIX (IDELE, 2014).
1.3.2 Le transfert d’immunité passive
1.3.2.1 Définition et mécanismes
Le transfert d’immunité passive est défini comme le transfert d’immunoglobulines produites par la mère et présentes dans sa circulation sanguine vers sa progéniture. Cette transmission de la mère au jeune peut avoir lieu avant la naissance, après la naissance ou les deux (BRAMBELL, 1958).
Les petits ruminants possèdent un placenta dit syndesmochorial ou synépithéliochorial. Pour ce type de placenta, l’épithélium utérin disparait plus ou moins, laissant le chorion en contact avec le tissu conjonctif de l’endomètre (AMOROSO, 1961). Le placenta possède une activité métabolique permettant d’assurer un transport actif et sélectif des substances indispensables au développement du fœtus. Il intervient également dans la protection vis-à-vis de certaines molécules toxiques et des agressions virales et bactériennes (ZARROUK et al., 1998). Néanmoins, le placenta syndesmochorial inhibe le
passage des immunoglobulines et autres facteurs immunologiques vers le fœtus (BORGHESI et al., 2014),
les nouveau-nés sont alors considérés comme agammaglobulinémiques (THIRY et al., 2002). Le système
immunitaire des jeunes ruminants est donc reçu, de façon passive, par l’ingestion du colostrum contenant les immunoglobulines qui sont alors absorbées au niveau de l’intestin (BORGHESI et al., 2014).
Une fois les immunoglobulines du colostrum absorbées dans l’intestin, celles-ci gagnent la circulation sanguine de l’animal, empêchant une propagation systémique des infections. Cette protection fait intervenir les IgG mais également les IgM (JACQUES, 2012). Il faut noter que cette protection, apportée par les immunoglobulines maternelles, diminue pendant les premières semaines de vie du fait de leur utilisation et leur catabolisme (PISARSKA et al., 2002).
Page | 20 Le transfert de l’immunité passive ne peut se faire que par une accumulation de très grandes quantités d’immunoglobulines intactes et fonctionnelles dans le colostrum et par une prise efficace de celles-ci par le nouveau-né (THIRY et al., 2002). L’efficacité de l’absorption est la plus élevée durant les
premières heures de vie puis diminue rapidement (PISARSKA et al., 2002). Les protéines colostrales que
l’animal a ingéré se retrouvent dans la lumière de l’intestin et sont alors absorbées par transcytose dans les entérocytes immatures, sous la forme de vésicules. Ce mode de transport est non spécifique et une grande partie des protéines se retrouve dans le sang en passant par le canal thoracique après être passée par la circulation lymphatique via exocytose (SALMON, 2011). Cette absorption des
protéines colostrales se déroule dans l’intestin grêle et plus particulièrement dans le jéjunum et dans l’iléon. Ainsi, le profil en immunoglobulines du sérum est similaire à celui du colostrum (JACQUES, 2012). L’intégrité des immunoglobulines est maintenue par différents systèmes. Il existe tout d’abord un facteur anti-trypsique ainsi qu’une activité protéolytique se réalisant dans l’intestin et permettant de limiter la dégradation des immunoglobulines par les enzymes. De plus, les immunoglobulines sont protégées grâce à une enveloppe glucidique (JACQUES, 2012).
Il a été observé chez les veaux que les cellules épithéliales de l’intestin perdaient leur capacité à absorber les macromolécules, dont les immunoglobulines, après plus ou moins 24 heures. L’efficacité d’absorption des IgG, chez les veaux, diminue de 0,3%/h entre la naissance et les 12 heures suivantes et jusqu’à 2,5%/h entre 12 heures et au moins 18 heures après la naissance (OSAKA et al., 2014).
Après consommation de colostrum, la concentration en immunoglobulines du sérum augmente (JACQUES, 2012) et atteint son maximum entre 24 et 48h après ingestion (CHEN et al., 1999; CASTRO et al., 2009). Il y a « Failure of passive transfer » (ou échec de transfert d’immunité passive) lorsque la
concentration en IgG du sérum est inférieure à 8 ou 10g/L (EHRHARDT et al., 2014;PISARSKA et al., 2002).
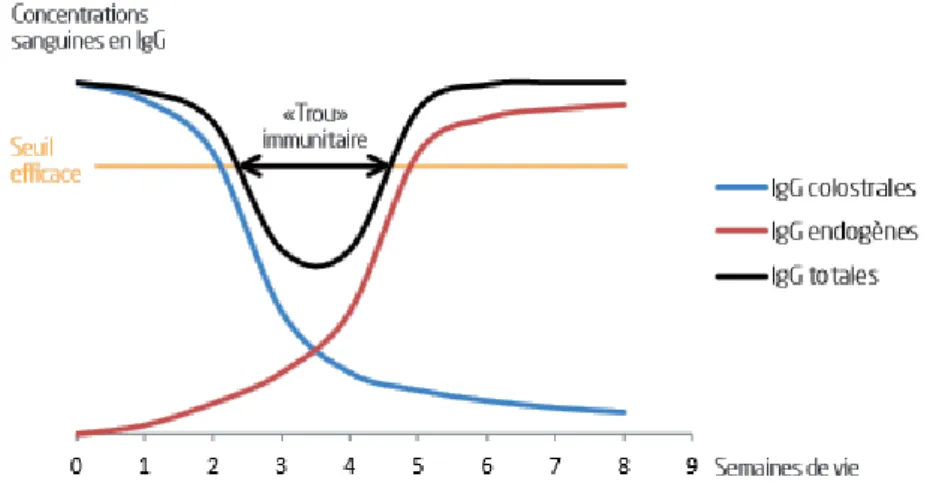
48 heures après la naissance, il est observé un déclin des immunoglobulines d’origine maternelle. Le nouveau-né va synthétiser des IgG de manière endogène, de l’ordre de 1g/j chez le veau (DEVERY et al.,
1979). Un « trou immunitaire » est présent vers 3-4 semaines d’âge (cf. fig. n°6), c’est-à-dire que la concentration en immunoglobulines sériques, d’origine colostrale ou endogène, est inférieure au seuil efficace de transfert d’immunité passive (OMACAPet al., 2015).
Figure 6 : Représentation schématique du "trou immunitaire" chez le chevreau (OMACAPet al., 2015) 1.3.2.2 Facteurs de variation
Différents facteurs peuvent influencer le transfert et l’absorption des immunoglobulines chez le nouveau-né :
L’âge à la 1ère buvée de colostrum
La masse d’immunoglobulines colostrales ingérées
Présence de la mère : il a été montré que les veaux ayant la présence de la mère avait une concentration en immunoglobulines plus élevée que les veaux sans (WEAVER et al., 2000).
Page | 21
La prolifération microbienne dans la lumière de l’intestin avant ingestion de colostrum (JACQUES, 2012).
La vigueur de l’animal : qui impacte l’âge à la 1ère buvée et le volume de colostrum ingéré.
La génétique.
La vigueur est une caractéristique vitale des nouveau-nés car elle permet des comportements nécessaires à la survie de ceux-ci : se lever, trouver la mamelle, localiser le pis et boire le colostrum (NOWAK, POINDRON, 2006; HOMEROSKY et al., 2017). Il s’agit d’un des critères les plus importants, avec le
poids de naissance, la durée de gestation, la taille de la portée, le comportement de la mère, l’hypothermie et les conditions climatiques, qui affectent la mortalité des jeunes animaux (O’BRIEN,
SHERMAN, 1993). La vigueur à la naissance pourrait influencer la capacité de l’animal à consommer le
colostrum. Une vigueur faible peut augmenter le temps que l’animal met à se lever, retarder la consommation de colostrum, diminuer les quantités de colostrum ingérées et donc augmenter les chances de mortalité liées à un transfert d’immunité passive insuffisant (VASSEUR et al., 2009). D’après
NASH et al., la vigueur possède un impact plus important sur la survie du nouveau-né que le poids de naissance, la mortalité diminuant lorsque que la vigueur à la naissance augmente (NASH et al., 1996). Il
a par ailleurs été montré, chez les veaux, que le poids de naissance n’a pas d’impact sur la concentration en immunoglobulines du sérum, contrairement à la vigueur à la naissance (ROBISON et
al., 1988). Il a également été observé que les agneaux en bonne condition survivaient mieux quelle
que soit la qualité du lait distribué, contrairement aux agneaux faibles pour lesquels la mortalité est forte, même avec un accès à un lait de bonne qualité (NASH et al., 1996).
D’un point de vue bien-être, un animal peu vigoureux à la naissance peut ressentir la faim à cause d’une inhabilité à ingérer suffisamment de colostrum ou de lait et peut donc entrainer une production insuffisante de chaleur et par conséquent une hypothermie.
La vigueur à la naissance d’un animal peut être influencée par son poids de naissance (NASH et al.,
1996) mais également par la difficulté de mise bas de la mère. En effet, une vigueur plus faible chez les animaux provenant d’une mise bas difficile peut être expliquée par la présence de lésions traumatiques, de durées d’interventions plus longues et/ou d’acidoses métaboliques résultant d’hypoxies ou d’asphyxies.
Chez les agneaux, un protocole a été mis en place afin de mesurer la vigueur sur une échelle de 5 scores avec une évaluation comportementale des animaux, cinq minutes après leur naissance (cf. tab. n°5) (MATHESON et al., 2011) :
Tableau 5 : Définitions des scores de vigueur chez les agneaux (MATHESON et al., 2011)
Score Description
0 Agneau extrêmement actif et vigoureux, debout sur ses 4 pattes
1 Agneau très actif et vigoureux, en appui sur ses pattes arrières et ses genoux
2 Agneau actif et vigoureux, sur le ventre avec la tête relevée
3 Agneau faible, couché mais capable de lever la tête
4 Agneau très faible, incapable de lever la tête, fait peu de mouvement
Suite à une étude menée sur le génome d’un croisement des races bovines Holstein et Jersey, il a été observé une part de génétique en lien avec le transfert d’immunité passive. En effet, il semble que la concentration en IgG du sérum des veaux possède une héritabilité h² modérée de 0,18. De plus, une région chromosomique en lien avec la concentration en IgG sérique a été identifiée sur le QTL nommé BTA20 (MALTECCA et al., 2009).
1.3.2.3 Méthodes d’évaluation
Comme pour l’évaluation du colostrum, il existe de nombreuses méthodes pour évaluer le transfert d’immunité passive chez les ruminants. Ces méthodes peuvent être directes ou indirectes. Méthodes directes :
Page | 22 La technique d’immunodiffusion radiale et le test ELISA, décrits précédemment, peuvent également être utilisés pour le sérum.
En plus de ces deux méthodes, il existe également l’électrophorèse des protéines plasmatiques ou sériques. Cela consiste en la séparation des protéines, dont les gammaglobulines (IgG, IgA et IgM), en fonction de leur poids. Des agents réducteurs vont être sous l’influence d’un champ électrique et vont charger négativement les immunoglobulines, changeant ainsi leur poids moléculaire. Il y a ensuite migration des protéines vers le pôle positif, la migration se réalisant plus vite lorsque le poids moléculaire est plus faible. Les molécules sont par la suite placées sur une membrane de nitrocellulose pour la détection des différentes gammaglobulines via des anticorps spécifiques (JACQUES, 2012). Méthodes indirectes :
La réfractométrie, expliquée plus haut, peut aussi permettre l’évaluation de la teneur totale en protéines du sérum.
Il existe également 2 tests de précipitation :
Le test de précipitation au sulfite de sodium se réalise par l’utilisation de trois solutions de sulfite de sodium à concentration 14, 16 et 18%. Ces solutions causent la précipitation sélective des protéines à poids moléculaire élevé. Une précipitation en utilisant la solution à 14% est indicatrice d’une concentration en immunoglobulines plus élevée qu’une précipitation avec la solution à 16% et 18%. Il y a absence de transfert d’immunité lorsqu’il n’y a pas de précipitation avec la solution à 18% (WEAVER et al., 2000).
Le test de précipitation au sulfate de zinc fonctionne sur le même principe que le test précédent. 0,1 ml de sérum est ajouté à 6 ml de sulfate de zinc de concentration 208 mg/L. La solution est incubée 30 minutes à température ambiante. La lecture se réalise ensuite par spectrophotométrie en évaluant le degré de précipitation (WEAVER et al., 2000).
Le test de coagulation au glutaraldéhyde fonctionne par le fait que les amines non chargées des protéines vont former des liens avec les groupements aldéhydes donnant ainsi des caillots. La formation de ces caillots est en relation avec les gammaglobulines. Pour ce test, 1,5ml de sérum est ajouté à une solution de glutaraldéhyde. Si un caillot se forme en moins de 5 minutes, cela signifie qu’il y a bien transfert d’immunité passive (WEAVER et al., 2000).
La dernière méthode est la mesure de l’activité de la GGT (Gamma-Glutamyl-Transférase). La GGT est une enzyme produite par les cellules ductiles de la glande mammaire et se retrouvant dans le colostrum. Il a été montré que l’activité de la GGT augmente rapidement après ingestion puis diminue brusquement durant les 24 prochaines heures avant de diminuer progressivement pendant les 2 prochains mois. L’activité de la GGT peut confirmer une ingestion de colostrum mais ne peut cependant pas permettre de connaitre la concentration en IgG du sérum. Il est toutefois possible d’interpréter l’activité de la GGT pour savoir si le transfert d’immunité passive s’est bien réalisé. Chez le veau, il est recommandé d’avoir une activité de la GGT supérieure à 200 UI/L à 1 jour, supérieure à 100 UI/L à 4 jour et supérieure à 75 UI/L à 1 semaine. Si l’activité de la GGT est inférieure à 50 UI/L durant les 2 semaines de vie du veau, il est alors considéré qu’il y a eu échec de transfert d’immunité passive (WEAVER et al., 2000).
1.3.3 Santé et bien-être du chevreau
1.3.3.1 Définition du bien-être
Le bien-être animal est défini par l’ANSES comme « […] l’état mental et physique positif lié à la satisfaction de ses besoins physiologiques et comportementaux, ainsi que de ses attentes. Cet état varie en fonction de la perception de la situation par l’animal. » (ANSES, 2018).