Neighbour origin and ploidy level drive impact of an alien invasive plant species in a competitive environment
14
0
0
Texte intégral
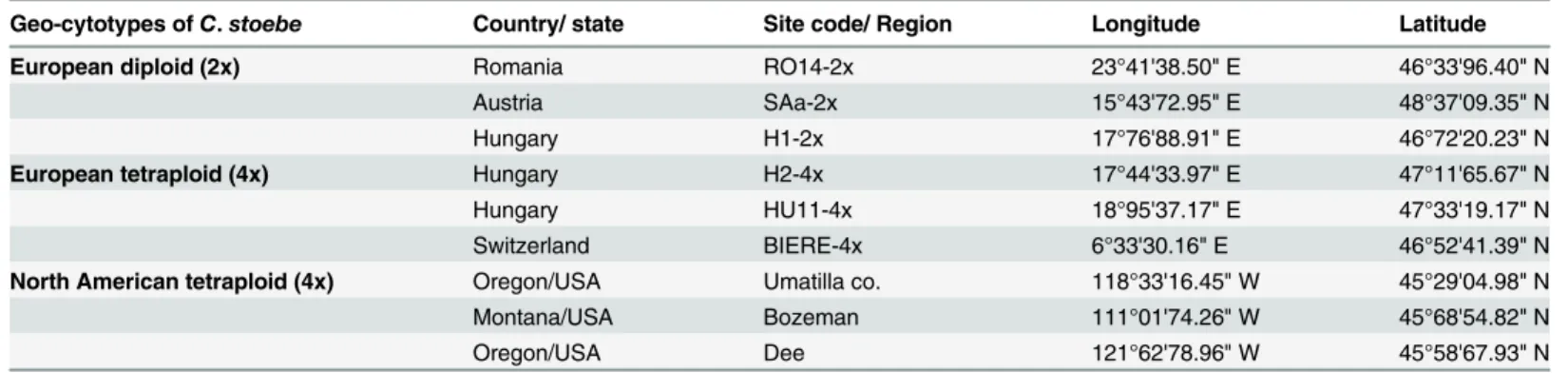
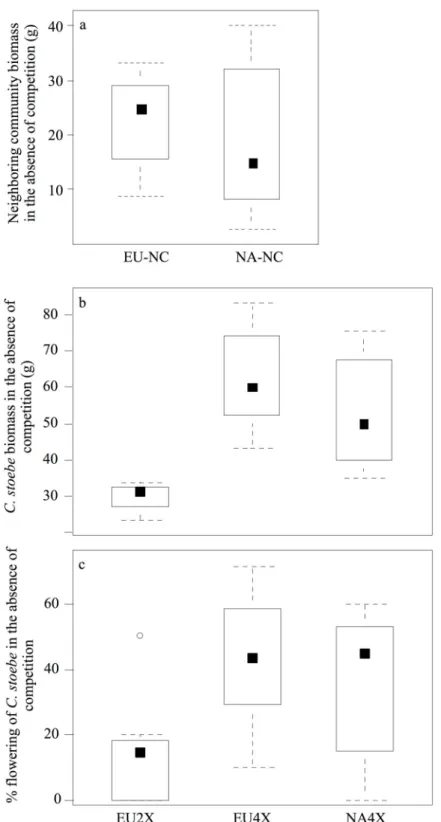
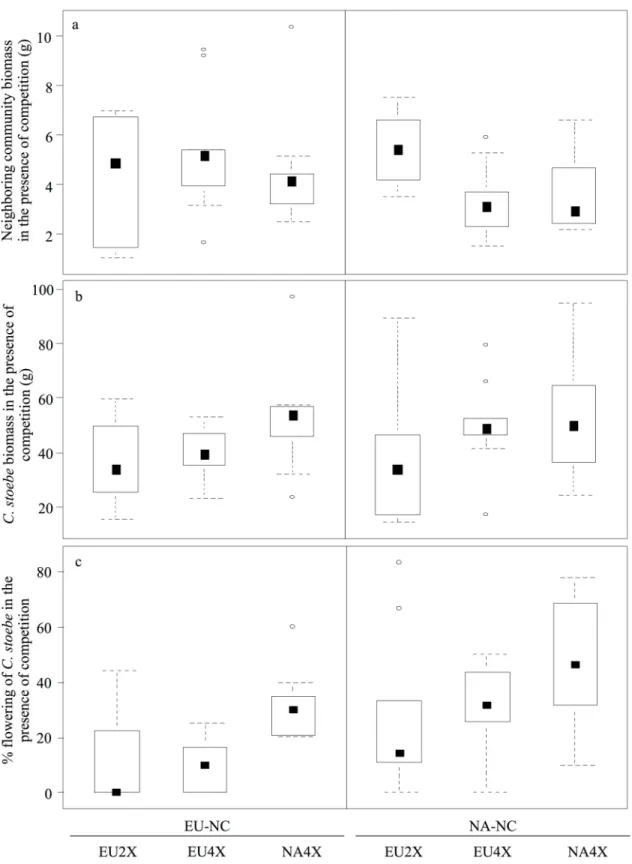
Figure

+2

Documents relatifs