Review
Contribution of connexins to the function of the vascular wall
Jacques-Antoine Haefliger
a,*, Pascal Nicod
a, Paolo Meda
ba
Laboratory of Molecular Biology, Department of Internal Medicine, University Hospital, 19-135S Centre Hospitalier Universitaire Vaudois, CHUV-1011, Lausanne, Switzerland
b
Department of Morphology, University of Geneva CMU, 1211 Gene`ve 4, Geneva, Switzerland Received 30 September 2003; received in revised form 31 October 2003; accepted 17 November 2003
Time for primary review 26 days
Abstract
Gap junction channels provide an enclosed conduit for direct exchanges of signalling molecules, including ions and small metabolites between cells. This system of communication allows cells to monitor the functional state of their neighbours, and is rapidly modulated to continuously adapt to the immediate needs of groups of coupled cells. In the major arteries, endothelial cells may express three connexins isotypes, namely Connexin37 (Cx37), Cx40 and Cx43, whereas the underlying smooth muscle cells may express Cx37, Cx40, Cx43 and Cx45. Moreover, myoendothelial gap junctions have also been shown to be involved in the regulation of vascular tone. This review highlights the regulation of vessel connexins in response to injury, as observed during experimental hypertension or wound repair, as well as the consequences of loss of one connexin in different transgenic null mice. In view of the major endocrine role of the kidney in the control of blood pressure, we also discuss the distribution of connexins in the kidney vasculature. Cx40 is present between endothelial cells of vessels and glomeruli, as well as between renin-secreting cells, the modified smooth muscle cells which form the wall of the terminal part of afferent arterioles. Modulation of Cx40 expression in a model of renin-dependent hypertension suggests that this connexin may be implicated in the function of renin-secreting cells. Finally, to address the possible regulation of connexin expression by fluid pressure, we summarize the effects of elevated transmural urine pressure on bladder Cx43 expression.
D 2003 European Society of Cardiology. Published by Elsevier B.V. All rights reserved.
Keywords: Cell – cell communication; Gap junctions; Connexins; Vasculature; Vessels; Aorta; Smooth muscle cells; Endothelium; Blood pressure; Hypertension
1. Diversity of connexin expression in the arterial wall The wall of arteries undergoes both physiological and pathological changes in response to mechanical stress and on exposure to hormonal stimuli. This wall consists of two communication departments, the smooth muscle and the endothelium, both of which are connected by gap junctions. In the vascular wall, four connexins have been detected in a pattern that varies depending on the cell type and the compartment.
2. The intima layer
The vascular endothelium consists of a continuous monolayer of cells, lining the luminal surface of the entire vascular system, and which provides a structural and met-abolic barrier between the blood and the underlying tissues. Endothelial cells are induced to migrate during the process of new capillary blood formation and during repair of the endothelial lining which result from injury of large vessels. Moreover, the endothelium plays a central role in the regulation of the vascular tone [1].
Communication between endothelial cells, as assessed by diffusion of fluorescent dyes, has been demonstrated in both large and micro-vessels [2 – 4]. Due to the necessity of establishing a continous, non-thrombogenic covering during the formation and renewal of blood vessels, endothelial cells often move as sheets, in which groups of coupled cells 0008-6363/$ - see front matterD 2003 European Society of Cardiology. Published by Elsevier B.V. All rights reserved.
doi:10.1016/j.cardiores.2003.11.015
* Corresponding author. Tel.: 26; fax: +41-21-314-09-68.
E-mail address: [email protected] (J.-A. Haefliger).
migrate in a coordinate manner. The contribution of con-nexins to this coordination has been tested by mechanically removing endothelial cells within confluent monolayers, which induces the cells bordering the experimental wound to proliferate and to migrate into the denuded area. Using cells derived from micro-vessels, it has been found that this in vitro migration is locally associated within an increased expression of Cx43 and coupling, which is not observed in the quiescent cells distant from the wound [2,3,5]. Con-ditions blocking migration, but not cell division, prevented the coupling increase, and, conversely, a pharmacological as well as a genetic inhibition of gap junctions channels altered the movement of endothelial sheets, without affecting the migration of individual cells, thus delaying the closure after wound [3,5]. Connexins expression is not uniform in the endothelium of all vessels and may vary depending on the species. Thus, native endothelial gap junctions consist of Cx37, Cx40 and Cx43, depending on the type of vessel and its position in the vascular tree [6 – 16]. In large arteries, Cx40 and Cx37 are widely distributed within the vascular endothelium whereas Cx43 shows a more heterogeneous expression pattern[6,12] (Figs. 1 and 2). The occurrence of extensive endothelial cell coupling in situ[17]indicates that connexins may coordinate the migration of endothelial cells under in vivo conditions, as suggested by the in vitro experiments [3,5]. To test this hypothesis, the spatial and temporal expression pattern of endothelial gap junctions has been investigated during healing of rat carotid artery after a
denudation injury [11]. Regeneration of the injured endo-thelium was found to be first associated with a decreased number of gap junctions, possibly as a result of the initial cell injury [11], which, subsequently, returned to levels found in uninjured controls. This process involved Cx43, Cx40 and Cx37, that appeared colocalized to the same gap junction plaque early in the regeneration process [10,11]. Within the 28-day duration of the study, the levels of Cx40 returned to the control levels observed in the uninjured vessel, whereas those of Cx37 and Cx43 were higher[11]. This behaviour suggests that the three main vascular con-nexins are differentially regulated to ensure specific patterns of communications between cells, which vary during the healing process. Co-expression of multiple connexins in a single cell also makes possible the formation of heteromeric connexons, providing a way to form many more channels types that permitted in a homomeric configuration[18 – 21]. The existence of heteromeric connexons made of Cx43 and Cx40 has been reported in vascular smooth muscle cells
[22]and channels made of Cx43 and either Cx45 or Cx37 may form as well [20,23]. The abundance of various heteromeric channels may therefore be altered by changes in the ratio of the connexins co-expressed by vascular cells, providing a suitable mechanism for fine tuning of cell – cell communication in the vascular wall.
Cx37 and Cx40 are the only connexins expressed by the mouse aortic endothelium and, recently, the contribution of these proteins has been studied in transgenic mice lacking
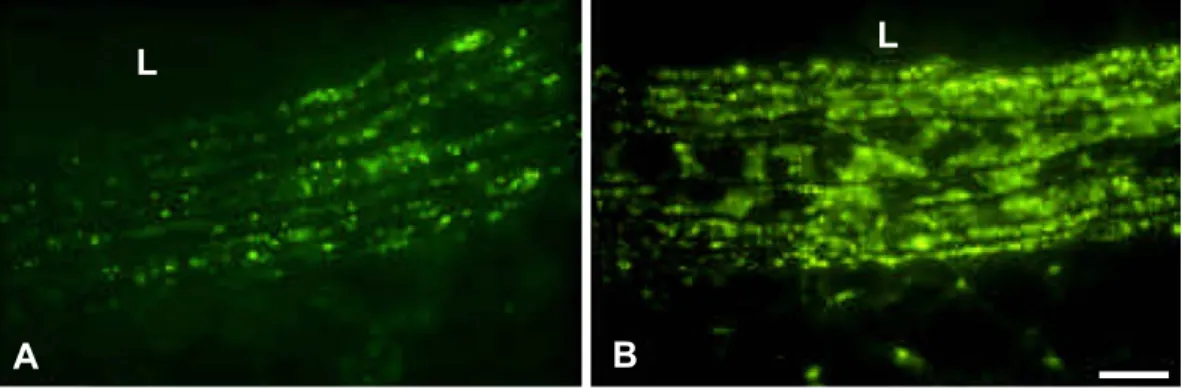
Fig. 1. Different connexins link the cells of large arteries. Immunofluorescence labeling reveals that the endothelial cells of large arteries express abundant levels of Cx40 (A), lower amounts of Cx37 (B) and, in some restricted areas, even lower levels of Cx43 (C). The latter connexin, as well as Cx45 (D), is also detected, as sparse spots in the vessel media, presumably corresponding to contacts between the smooth muscle cells of this pig coronary. L = lumen of vessel. Bar, 50 Am.
Cx40 (Cx40 / ) and/or Cx37 (Cx37 / ) in the aortic endothelium [15,24]. The intercellular transfer of injected dyes was found to be altered in Cx40-deficient aortic endothelium, under conditions leading to upregulation of Cx37[15]. Other dye transfer experiments, testing interen-dothelial cell communication in aortic segment of wild-type, Cx37 / , Cx40 / , Cx37+/ Cx40 / , and Cx37 / Cx40 / mice showed that both Cx37 and Cx40 are crucial for endothelial cell communication. The two connexins are mutually dependent on each other for optimal expression in
vascular endothelium, inasmuch as elimination of either Cx40 or Cx37 resulted in a decrease of the non-ablated connexin, whereas both Cx37 and Cx43 increased in the media layer of Cx40 / mice[24]. Thus, it appears that, in response to connexin deletion, a decrease in another, co-expressed connexins occurs preferentially rather than a compensatory upregulation, indicating that intercellular communication is determined not only by the properties of individual connexins, but also by the interactions of con-nexins within heteromeric channels with novel properties
[20,21]. In an other study, however, lack of Cx40 in endothelium was associated with an increase of Cx37
[15]. Mice lacking both Cx37 and Cx40 are not viable beyond the first postnatal day and exhibit severe vascular abnormalities [25]. They show localized hemorrhages in different tissues, as well as blood vessel dilation and congestion, suggesting that endothelial communication is required for the normal development and/or functional maintenance of portions of the mouse vasculature [25].
3. The media layer
Gap junction proteins may also coordinate the mechan-ical contractions of smooth muscle cells, possibly to insure a proper modulation of the vasomotor tone of the vessel wall. Vasoconstriction and vasodilation travel rapidly along the vessel network, due to the conduction of signals between endothelial and/or smooth muscle cells [26] through gap junctions. Vascular smooth muscle cells of both resistance and conduit arteries express predominantly Cx43 and Cx45, albeit Cx37 and Cx40 have also been reported in certain vessels [8,14,27 – 32]. The space constant of the spreading electrical signals within the media, as measured with a partition chamber, demonstrates that vascular smooth mus-cle cells behave as an electrical syncitium[26,33].
Within vessels, the endothelial and the smooth muscle cells are separated by connective tissue and the internal elastic membrane (Figs. 1 and 2). However, these two cell types also establish close contacts with each other, via myo-endothelial bridges that cross fenestration of the internal elastic lamina[34,35]. This arrangement provides the basis of different cell – cell communication pathways in hamster arterioles. The effect of these connections varies in different vessels, depending on the number of layers of smooth muscle cells. In arterioles, in which a single layer of smooth muscle surrounds the monolayer of endothelial cells, a bidirectional electrical communication has been demonstrat-ed [36,37]. Thus, a change in the membrane potential of endothelial cells is electronically transmitted to smooth muscle and, conversely, the membrane potential of endo-thelial cells is influenced by that of smooth muscle cells. This reciprocal influence serves to amplify the spreading of cytosolic free calcium [38]and the transmission of hyper-polarization induced by bradykinin from the endothelial cells to smooth muscle cells [39]. The consequences of Fig. 2. Cx40 and Cx37 form gap junction plaques between endothelial cells
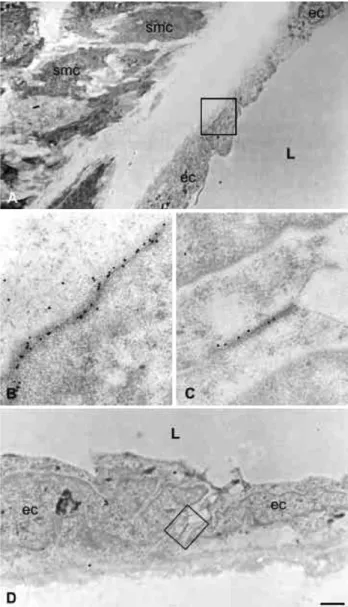
of large arteries. Immunocytochemical staining using specific antibodies against either Cx40 (A and B) or Cx37 (C and D) and protein A-coated gold particles, reveal that the two connexins are concentrated in minute areas of apposition (boxed in A and D) between the membranes of adjacent endothelial cells (ec) of rat aorta. Note the absence of staining over nearby smooth muscle cells (smc). At larger magnifications of the areas boxed in A and D (B and C, respectively), the immunostained regions of the membrane feature an appearance typical of gap junctions. smc = smooth muscle cell; ec = endothelial cell, L = lumen of vessel. Bar, 1 Am in A, 0.5 Am in D and 150 nm in B and C.
such a coupling is that the arteriolar wall works as func-tional unit, with unique properties resulting from the inter-play between excitable and nonexcitable components.
In muscular arteries, signal conduction is also efficient from smooth muscle to endothelial cells [40], whereas signals passing in the opposite direction are rapidly dissi-pated in the multiple layers of the media [41]. Thus, symmetrical electrical coupling can not be evidenced in arteries, and heterocellular communication is unidirectional due to an asymmetry in input resistance between the coupled cells of media and intima. Calcium fluxes have been shown to play a central regulatory role in vasomotion and it has been demonstrated that exchange of cations between smooth muscle and endothelial cells involves heterocellular gap junction channels[42].
Sites of electrical communication between endothelial and smooth muscle cells have been demonstrated in the mesenteric vascular bed [34], as well as in the resistance microvessels that control the blood flow to the hamster retractor muscle [35]. This bidirectional transmission of electrical and/or chemical signals ensures that both cell types display equivalent membrane potential, thus contributing to control blood flow and vasomotor responses along large distances. Myoendothelial gap junctions may also account, at least in part, for the response evoked by the endothelium-derived hyperpolarization factor (EDHF) [43 – 47] in the mesenteric vascular bed [34,48]. The relationship between the existence of myo-endothelial gap junctions[8,34,35,49 – 52], endothelial cell hyperpolarization and EDHF-induced hyperpolarization of smooth muscle cells has been recently studied in the rat mesenteric artery [48]. In this system, EDHF-mediated vasodilation is prominent and stimulation of the endothelium by acetylcholine (ACh) evokes a hyper-polarization of both endothelial and smooth muscle cells, which are connected by gap junctions. In contrast, in the femoral artery, where there is no EDHF-dependent relaxa-tion, ACh-evoked hyperpolarization was exclusively ob-served in endothelial cells, consistent with the lack of detectable myo-endothelial gap junctions. These data dem-onstrate that transfer of endothelial hyperpolarization and/or of small molecules from endothelial to smooth muscle cells connected by gap junctions is essential and sufficient to explain the EDHF induced effects in small vessels[48].
Typically, endothelial cells are more extensively inter-linked by gap junctions than the smooth muscle cells of the vascular wall, and gap junctional contacts between endo-thelial cells and superficial smooth muscle cells of small arteries and arterioles are established through discontinuities of the internal elastic lamina. Thus, homocellular coupling exists within both the endothelium layer and within the smooth muscle cell layer of these vessels, and heterocellular coupling also occurs between endothelial and smooth mus-cle cell, mostly in the intima to media direction[8]. These communications may be important during both the patho-physiological and the experimental repair of a damaged vessel.
4. Chronic hypertension alters the expression of connexins in aorta
Blood pressure, blood flow and shear stress modulate vascular diameter [39] and the integration of these forces may be achieved by gap junctions since coupling of vascular cells enables the rapid conduction of the electrical signals generating vasomotor responses [35,53 – 56]. Sev-eral studies have related the expression of endothelial Cx43 to shear stress and to mechanical load [12,50,57 – 59]. The regulation of blood flow requires the coordinated response of resistance and feeding arteries and, in turn, the proper function of these large vessels requires the coordi-nation of the smooth muscle cells of the vascular media. The aorta, which is a sparsely innervated and electrically quiescent vessel, is likely to be particulary dependent on gap junctional communications for coordinating the responses of smooth muscle cells to diverse neural and endothelial signals[54]. Conditions perturbing the function of the aortic wall, as observed during chronic hyperten-sion, are therefore expected to be associated to connexins alterations. To test this hypothesis, we evaluated the effects of chronic hypertension in two rat models resulting in a similar increase in blood pressure, thickening of the aortic wall and increase in the isobaric distensibility of the carotid [60,61]. In the mineralocorticoid-salt model (DOCA-salt), hypertension resulted from increased reten-tion of sodium chloride in the presence of suppressed renin secretion, whereas in the two kidney, one-clip model (2K, 1C), the development of hypertension resulted from the constriction of one renal artery and the ensuing activation of the renin-angiotensin system [62]. In a third rat model, in which a degree of hypertension similar to that observed in both 2K, 1C and DOCA-salt models was induced by inhibiting nitric oxide synthase (L-NAME
model), hypertension was associated with limited cardio-vascular hypertrophy, but not with increased isobaric carotid distensibility. Comparison of the three models, showed a comparable increase of the Cx43 expressed by the smooth muscle cells of the aortic media In the 2K, 1C and the DOCA-salt hypertensive rats (Fig. 3). In contrast, decreased levels of Cx43 were found in the smooth muscle cells of L-NAME-treated rats, which failed to show
in-creased arterial distensibility.
When exposed to chronic hypertension, conduit arteries show a preserved isobaric luminal diameter in spite of an outward hypertrophic remodelling, which results from the hypertrophy of smooth muscle cells and alterations of extracellular matrix, and which modifies the mechanical properties the vessels. Previous studies have shown that the distensibility and compliance of various arteries are in-creased under isobaric conditions in the 2K, 1C hyperten-sive animals, but not after inhibition of nitric oxide syn-thase, despite a thickening of the arterial wall which is similar in both hypertensive models. The different changes in Cx43 provide a clue that cell-to-cell communication
mediated by connexins may contribute to regulate the elasticity of the vascular wall.
Antibodies against Cx40 decorate punctate regions of large arteries, where the membranes of adjacent endothelial cells are closely apposed and feature the pentalaminar appearance typical of gap junctions (Figs. 1 and 2). In contrast, no labeling is detected in the smooth muscle cells and connective fibers of the same vessels (Figs. 1 and 2). The transcript of Cx40 was found increased in the aorta of 2K, 1C animals [63], as well as after balloon injury [64]. The molecular mechanism whereby an increase in blood pressure enhances the expression of both Cx40[63,65]and Cx43 genes in the aorta, while selectively changing only the former connexin species in the hypoperfused kidneys[63], is still undetermined. Cx43 is expressed by endothelial cells of large arteries[66], but almost exclusively at sites where branching or flow division is expected to create high turbulence [12]. Induction of a flow disturbance in a segment of abdominal aorta which was experimentally reduced by clipping, further demonstrated that Cx43 ex-pression is increased by hemodynamic forces in vivo[12]. Together, these studies indicate that Cx40 and Cx43 may represent suitable, tissue-specific markers to monitor the changes induced by hypertension in the vasculature. The altered expression of Cx43 in the aortae of hypertensive rats, which is predominantly attributed to changes occuring in the smooth muscle cells of the media, further raises the possi-bility that this protein may contribute to lack of autoregu-lation of arterial distensibility.
Studies in a genetic model of hypertension have demon-strated similar levels of Cx43 in the mesenteric arteries of spontaneously hypertensive and of control Wistar – Kyoto (WKY) rats[67], whereas the endothelium of tail arteries of spontaneously hypertensive rats (SHR) showed a significant decrease in Cx43, Cx40, and Cx37, as judged by immuno-histochemistry[68]. In contrast, Cx45[30]has been shown to increase in cerebral arteries of SHR animals, resulting in alterations of electrical connectivity between smooth muscle cells of brain vessels, but not of aorta and femoral artery. In
rats made hypertensive by treatment with L-NAME or
angiotensin II, no change in Cx45 was observed, suggesting that neither increased blood pressure nor angiotensin II are directly implicated in the increase in Cx45 observed in SHR rats[69].
5. Deleting vascular connexins in transgenic mice To investigate the function of Cx43 in endothelial cells, independently from that the connexin may play in smooth muscle cells, an endothelial-specific Cx43 deletion was studied in mice. In one study, loss of Cx43 in the endothelium was found to cause hypotension and brady-cardia [70]. This hypotension was associated with a marked elevation in the plasma levels of nitrate (an indicator of nitric oxide production), plasma angiotensin I and angiotensin II [70]. In another study, however, lack of Cx43 in endothelium appeared to have no effect on resting blood pressure [71]. The difference between the two transgenic models may be explained by a significant decrease in the levels of Cx43 of aortic smooth muscle cells [70], which would then indicate that lowered expres-sion of Cx43 between smooth muscle cells is critical for the regulation of vasomotor tone.
Loss of Cx45 after replacement of the coding region of the Cx45 gene by the bacterial h-galactosidase reporter gene, has shown that Cx45 is expressed in the smooth muscle layer of vessels and, to a much less extent, also in the endothelium at least at early stages of development[72]. Thus, full disruption of the Cx45 blocks vascular develop-ment after the normal initiation of vessel formation, and impairs the differentiation of smooth muscle cells. As a result, Cx45 / mice showed a narrowing or the absence of the dorsal aorta and rare blood vessels in the yolk sac, due to reduced levels of hematopoiesis, possibly caused by defec-tive TGFh signaling[72].
In mice lacking Cx40 [73,74], loss of this protein was shown to impair the conduction of dilatory signals along Fig. 3. Cx43 expression is upregulated in the smooth muscle cells of aorta of chronically hypertensive rats. Immunofluorescence labeling of rat aorta using specific antibodies, located Cx43 at discrete spots dispersed throughout the media layer, presumably at homotypic smooth muscle cell contacts (A). A sizable increase in the number and size of these spots was observed in the thickened aortic media of rats made hypertensive for four weeks after partial obstruction of one renal artery (B). Due to the focus plane, the limited amount of Cx43 sparsely expressed by some endothelial cells is not detectable in these figures. L = lumen of vessel. Bar, 50 Am.
arterioles, indicating that Cx40 plays a role in the propaga-tion of the vasodilapropaga-tions which are initiated by the local administration of acetylcholine (Ach) or bradykinin [75]. These results demonstrate a crucial role of Cx40 in mouse microcirculation. The Cx40-deficient mice were also hyper-tensive, indicating also a role of Cx40 in the control of blood pressure, possibly as a result of impaired endothelium-dependent dilation in response to Ach, and of lack of coordination between down-stream and up-stream vessels
[75]. Moreover, a spontaneous and irregular vasomotion was observed in a few Cx40-deficient mice that lead to arteriolar constriction, thus contributing to increase vascular resistance
[75]. Further studies have recently shown that impaired action or release of endothelial nitric oxide, as well as alterations in the efficiency of other endothelial dilators,
were not responsible for the hypertension of Cx40 / mice
[76]. Also, this hypertension does not seem to be caused by alterations in the renin-angiotensin system, in spite of the fact that Cx40 is strongly expressed in the renin-secreting cells[76].
The generation of mice lacking either Cx37 and Cx40
[24,25,73,74] or Cx45 [72] has provided evidence for the biological function of gap junctions in the development and/ or functional maintenance of mouse vasculature. There are similarities in the phenotype of Cx45 / and Cx37 / Cx40 / mice as both animals types show abnormally enlarged blood vessels and endothelium-lined cavernous vascular spaces[25,72]. However, these animals also differ from each other, inasmuch as Cx37 / Cx40 / mice do not show obvious anomalies of yolk sac vasculature or
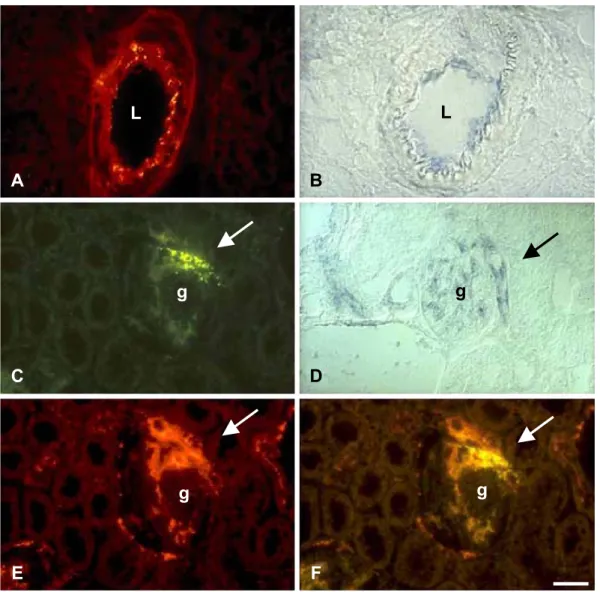
Fig. 4. Cx40 is expressed in the interlobular and afferent arterioles of rat kidney. Immunostaining with specific antibodies (A) and in situ hybridization with selective antisense cDNA probes (B) show that Cx40 is expressed by the endothelial cells of interlobular arteries of rat kidney. The two approaches also detect Cx40 in glomerular capillaries (C and D) and in the portion of the afferent arteriole (arrows) which is juxtapposed to a glomerulus (C and D). Immunostaining reveals that this region corresponds to the portion of the afferent arteriole that contains renin-producing cells (E). Double exposure of the same area showed that most of the Cx40 (green signal) actually colocalised with that of renin (red signal), resulting in a yellow staining at the distal part of the afferent arteriole (arrow) (F). g = glomerulus; L = lumen of vessel. Bar, 25 Am.
failure of smooth muscle cell formation. Moreover, unlike Cx37 / Cx40 / animals, Cx45 / display narrowing of some vessels and die much earlier, due to critical defects in all three embryonics blood vessel systems (vasculature of yolk sac, placenta and embryo proper). These distinct phenotypes may be due to cell-specific differences in the expression of various connexins.
6. Connexins of kidney vessels
In the kidney the juxtaglomerular region accomodates smooth muscle cells, endothelial cells, mesangial cells, macula densa cells and the renin-producing cells that are located in the distal part of the afferent arterioles. All these cell types are connected by gap junctions which also connect the endothelial and the renin-producing cells of the afferent arteriole[77 – 80]. Accordingly, mRNAs for nine connexins
species, referred to as Cx26, Cx32, Cx30.3, Cx31, Cx37, Cx40, Cx43, Cx45 and Cx46, have been found in the kidney
[77,81]. With regard to vessels, Cx43 is expressed by the endothelial cells of medium size and small kidney arterioles, including the afferent arterioles[63], but not by the media cells of the latter vessel and, barely, by glomerular capillaries
[63]. Immunolabeling of kidney sections further showed that Cx40 is expressed by most endothelial cells of vessels and glomeruli, as well as by the renin-secreting cells[63,82,83] (Figs. 4 and 5).
Ryan et al. [84] have recently demonstrated that cells of the renin-expressing line AS4.1 form gap junctions with both endothelial cells and smooth muscle cells, and express RNAs coding for Cx40, Cx37, Cx43 and Cx45. In vivo, the predominant connexin of the preglomerular rat vasculature is Cx40, present in the media of the distal parts of the afferent arterioles, whereas Cx40 and Cx37 connect the endothelial cells of the proximal parts of
Fig. 5. Cx40 links the cells of the juxta-glomerular apparatus, which control blood pressure. Immunostaining of the juxta-glomerular apparatus of a rat kidney, locates Cx40 at minute areas of apposition between the membranes of renin-containining cells (box 1), as well as between those of endothelial cells (box 2). Higher magnification views of the areas boxed in panel A are shown in panels B (box 2) and C (box 1). Immunostaining for renin, using gold particles of 9 nm diameter, showed the hormone within characteristic secretory granules (D). Further immunostaining of the same section, using particles of 15 nm diameter, shows that Cx40 (arrow in D) is also found at contacts between renin-producing (rpc) and enthothelial cells (ec). g = glomerulus; rbc = red blood cell; ec = endothelial cell; rpc = renin-producing cell; L = lumen of afferent arteriole. Bar, 1 Am in A, 150 nm in in (B – D).
these vessels [83]. Only a weak staining for Cx43 has been observed in the media of the afferent arteriole [82]. Together, these data indicate that cells of the preglomer-ular vessels are, presumably, well coupled. Accordingly, the afferent arteriole admits vascular conducted responses
[85,86], and the tubulo-glomerular feedback response elicited by the macula densa may propagate more than a millimeter upstream the glomerulus, along preglomeru-lar vessels [86]. Coupling within preglomerular vessels is likely provided by Cx40, a connexin that could form both homo-(endothelial-endothelial or smooth muscle cell-smooth muscle cell) or hetero-cellular (endothelial-renin) connections (Figs. 4 and 5).
In view of the major endocrine role of the kidney in the control of blood pressure, we have investigated connexin expression in a renin-dependent model of hypertension after clipping one renal artery of rat kidney (two kidney, one-clip-2K, 1C-model). Northern blot analysis of polyA+ RNA demonstrated that, compared to sham-operated ani-mals, the hypertensive 2K, 1C animals featured an increase in Cx40 mRNA expression in both left (clipped) and right (unclipped) kidneys. In contrast, Cx43 mRNA expression was only increased in the latter organ. Antibodies con-firmed that the levels of Cx40 were actually increased in the kidneys of hypertensive animals due, at least in part, to enhanced expression of this protein in the renin-secreting cells. These data indicate that cell-to-cell communication mediated by Cx40 may be implicated in the function of renin-secreting cells, hence participating in the control of blood pressure [63]. Accordingly, Cx40-deficient mice were found to be hypertensive when compared to wild-type controls [75]. To test whether enhanced release of renin may be the cause of the observed hypertension, the vasoconstriction induced by angiotensin II was blocked using candesartan, a AT1 receptor antagonist. Under these conditions, arterial blood pressure was reduced in both wild type animals and in mice lacking Cx40. However, Cx40 / mice still exhibited a higher blood pressure than controls, suggesting that the acute pressure effects of angiotensin II were not responsible for the observed hyper-tension[76]. We are currently developing a model to eval-uate the consequence of the loss of Cx40 between renin-secreting cells (Haefliger et al., unpublished) in chronically hypertensive mice featuring increased levels of circulating renin.
7. Cx43 of smooth muscle cells is increased by stretching Due to the rapid turnover of connexins, the plasticity of their tissue-specific expression in response to various stim-uli, and the fast dynamics of the channels these proteins form, signaling via connexin channels can ensure a moment-to-moment, as well as a chronic remodeling of intercellular circuits [87]. The bladder is a sparsely innervated and electrically quiescent tissue, in which intercellular signaling
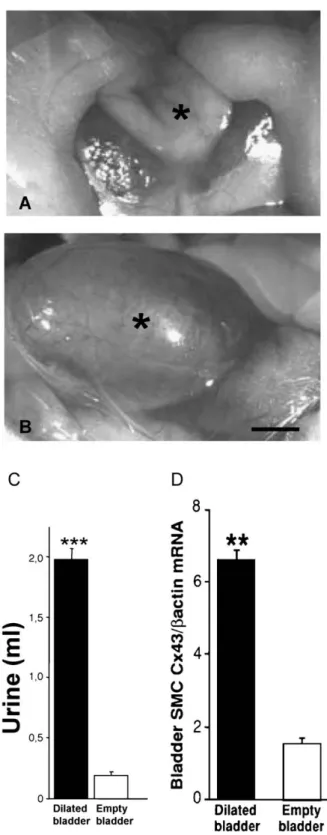
Fig. 6. Stretching of smooth muscle cells increases Cx43 expression. In control rats, the bladder (asterisk) contains at rest a minimal volume of urine (A). Eight hours after the surgical obstruction of the low urinary tract, the bladder dilates (B), due to a tenfold increase in the volume of retained urine (C). This change resulted in a stretching of smooth muscle cells that was associated to increased expression of Cx43 (D). ***p < 0.001. Bar, 3 mm.
is likely to be particulary dependent on gap junctional communication. Intercellular communication must ensure a coordinated response of the cells forming the detrusor muscle to ensure normal bladder function. To test this hypothesis, we evaluated the effects of elevated transmural urine pressure on the expression of Cx43 by bladder smooth muscle cells [88,89]. Using a model of bladder outlet obstruction in the rat (Fig. 6), a progressive increase of Cx43 mRNA and protein was detected as a function of bladder dilation [88]. Maximum transcript expression occured 7 – 8 h after obstruction, whereas removal of the urethral ligature decreased Cx43 mRNA expression within 4 h[88]. In cell culture, smooth muscle cells of guinea-pig and human detrusor muscle are coupled by gap junction channels containing at least Cx43 [90,91]. Recently, the role of intercellular communication mediated by Cx43 was evalu-ated during bladder overactivity, following 6 weeks of partial urethral outflow obstruction [89]. Under these conditions, Cx43 was also increased at the transcript and protein levels, in parallel with an increase in spontaneous bladder contrac-tions, which were reduced in the presence of heptanol. Moreover, carbachol-precontracted bladder strips from obstructed animals were more sensitive to the heptanol-induced relaxation than strips from empty bladders [89]. Outlet obstruction is a common medical disorder which leads to a rearrangement of smooth muscle cells within the bladder wall and to alterations of their function. The mechanisms underlying these changes remain mostly unknown. The available data indicate that, by integrating signals during urine storage, and the associated mass action on the detrusor smooth muscle cells, Cx43 may be relevant in this context. Measurements of electrical impedance have shown that detrusor cells are electrically coupled [92]. At least in the guinea-pig, the propagation of spontaneous excitation, and associated calcium waves propagate between smooth muscle cells through gap junctions[93]. Thus, the increase in Cx43 observed after stretch may be important to regulate bladder tone and contractility, by changing the asynchronous con-tractile activity which is typical of control, adult bladder into a more coordinated contraction.
8. Conclusions and perspectives
The over-active dilated bladders, as well as the hyper-trophied aortae of hypertensive rats, are examples of quies-cent organs. In vessels, an increase in connexin expression may be implicated in the structural and functional alterations that take place after mechanical stretch, and could represent a compensatory response to the numerous insults associated with increased fluid pressure, possibly to control the elas-ticity of the bladder, or vessel wall. Thus, Cx43 appears to be an early-activated sensor of mechanical changes, which is presumably required for proper adaptation of the elasticity and contractibility of the smooth muscle cells to increased transmural pressure. The mechanism underlying the
stretch-induced changes in connexin expression remains to be elucidated. The finding that these changes take place within hours and are reversible within the same time course, indicate that connexin modulation is an early event associ-ated with stretch and the ensuing increase in transmural pression, as well as with the return of these two parameters to control values. The time course of these changes is also significantly faster than that required to observe structural wall changes, such as smooth muscle cell hypertrophy and hyperplasia, suggesting that alterations of the normal con-nexin pattern may induce the subsequent structural and functional changes in muscle. In this perspective, it is worth mentioning that AP-1, a transcription factor implicated in the MAPK-mediated gene regulation induced by osmotic stress, and AP-1 binding activity, are both increased by shear stress in endothelial cells [94 – 96]. It is therefore possible that the increased levels of Cx43 mRNA observed in the muscle layers of stretched bladders, may also be induced by the higher AP-1 transcription factor activity which is observed under such conditions[97]. An involve-ment of AP-1 activity in the induction of Cx43 expression has been demonstrated in smooth muscle cells of human myometrium at term[98], and may be also involved in the upregulation of Cx43 expression in other types of smooth muscle cells. This consideration does not exclude the participation of other factors. Thus, angiotensin II has been shown to upregulate Cx43 expression in other types of cultured neonatal rat myocytes[99]and mechanical stretch induces the receptors for this hormone in the bladder, similarly to multiple other receptors, transcription factors, growth factors [100]. These findings show that connexins, and noticeably Cx43 are modulated in vivo, under condi-tions of mechanical stress.
Acknowledgements
The Haefliger team is supported by grants from the Swiss National Science Foundation (31-68036.02), the Novartis Foundation, the Juvenile Diabetes Foundation International (1-2001-555), the Placide Nicod and the Octav and Marcella Botnar Foundations. The Meda team is supported by grants from the Swiss National Science Foundation (31-67788.02), the Juvenile Diabetes Foundation International (1-2001-622), the European Union (QLRT-2001-01777), the Fonda-tion Romande pour la Recherche sur le Diabe`te and the National Institute of Health (1RO1 DK-63443-01).
References
[1] Furchgott RF, Zawadzki JV. The obligatory role of endothelial cells in the relaxation of arterial smooth muscle by acetylcholine. Nature 1980;288:373 – 6.
[2] Pepper MS, Meda P. Basic fibroblast growth factor increases junc-tional communication and connexin 43 expression in microvascular endothelial cells. J Cell Physiol 1992;153:196 – 205.
[3] Pepper MS, Montesano R, el Aoumari A, et al. Coupling and con-nexin 43 expression in microvascular and large vessel endothelial cells. Am J Physiol 1992;262:C1246 – 57.
[4] Little TL, Xia J, Duling BR. Dye tracers define differential endothe-lial and smooth muscle coupling patterns within the arteriolar wall. Circ Res 1995;76:498 – 504.
[5] Kwak BR, Pepper MS, Gros DB, et al. Inhibition of endothelial wound repair by dominant negative connexin inhibitors. Mol Biol Cell 2001;12:831 – 45.
[6] Bruzzone R, Haefliger JA, Gimlich RL, et al. Connexin 40, a com-ponent of gap junctions in vascular endothelium, is restricted in its ability to interact with other connexins. Mol Biol Cell 1993;4:7 – 20. [7] Reed KE, Westphale EM, Larson DM, et al. Molecular cloning and functional expression of human connexin 37, an endothelial cell gap junction protein. J Clin Invest 1993;91:997 – 1004.
[8] Little TL, Beyer EC, Duling BR. Connexin 43 and connexin 40 gap junctional proteins are present in arteriolar smooth muscle and en-dothelium in vivo. Am J Physiol 1995;268:H729 – 39.
[9] Yeh HI, Dupont E, Coppen S, et al. Gap junction localization and connexin expression in cytochemically identified endothelial cells of arterial tissue. J Histochem Cytochem 1997;45:539 – 50.
[10] Yeh HI, Rothery S, Dupont E, et al. Individual gap junction plaques contain multiple connexins in arterial endothelium. Circ Res 1998; 83:1248 – 63.
[11] Yeh HI, Lai YJ, Chang HM, et al. Multiple connexin expression in regenerating arterial endothelial gap junctions. Arterioscler Thromb Vasc Biol 2000;20:1753 – 62.
[12] Gabriels JE, Paul DL. Connexin 43 is highly localized to sites of disturbed flow in rat aortic endothelium but connexin 37 and con-nexin 40 are more uniformly distributed. Circ Res 1998;83:636 – 43. [13] Traub O, Hertlein B, Kasper M, et al. Characterization of the gap junction protein connexin 37 in murine endothelium, respiratory epithelium, and after transfection in human HeLa cells. Eur J Cell Biol 1998;77:313 – 22.
[14] van Kempen MJ, Jongsma HJ. Distribution of connexin 37, connex-in 40 and connexconnex-in 43 connex-in the aorta and coronary artery of several mammals. Histochem Cell Biol 1999;112:479 – 86.
[15] Kruger O, Beny JL, Chabaud F, et al. Altered dye diffusion and upregulation of connexin 37 in mouse aortic endothelium deficient in connexin 40. J Vasc Res 2002;39:160 – 72.
[16] Ko YS, Yeh HI, Rothery S, et al. Connexin make-up of endothelial gap junctions in the rat pulmonary artery as revealed by immuno-confocal microscopy and triple-label immunogold electron micro-scopy. J Histochem Cytochem 1999;47:683 – 92.
[17] Meda P, Spray D. Gap junctions function. Adv Mol Cell Biol 2000;30:263 – 322.
[18] Jiang JX, Goodenough DA. Heteromeric connexons in lens gap junction channels. Proc Natl Acad Sci U S A 1996;3:1287 – 91. [19] Bevans CG, Kordel M, Rhee SK, et al. Isoform composition of
connexin channels determines selectivity among second messengers and uncharged molecules. J Biol Chem 1998;273:2808 – 16. [20] Valiunas V, Gemel J, Brink PR, et al. Gap junction channels formed
by coexpressed connexin 40 and connexin 43. Am J Physiol 2001; 281:H1675 – 89.
[21] Cottrell GT, Wu Y, Burt JM. Cx40 and Cx43 expression ratio influ-ences heteromeric/heterotypic gap junction channel properties. Am J Physiol 2002;282:C1469 – 82.
[22] He DS, Jiang JX, Taffet SM, et al. Formation of heteromeric gap junction channels by connexins 40 and 43 in vascular smooth muscle cells. Proc Natl Acad Sci U S A 1999;96:6495 – 500. [23] Martinez AD, Hayrapetyan V, Moreno AP, et al. Connexin 43 and
connexin 45 form heteromeric gap junction channels in which indi-vidual components determine permeability and regulation. Circ Res 2002;90:1100 – 7.
[24] Simon AM, McWhorter AR. Decreased intercellular dye-transfer and downregulation of non-ablated connexins in aortic endothelium de-ficient in connexin 37 or connexin 40. J Cell Sci 2003;116:2223 – 36.
[25] Simon AM, McWhorter AR. Vascular abnormalities in mice lacking the endothelial gap junction proteins connexin 37 and connexin 40. Dev Biol 2002;251:206 – 20.
[26] Beny JL, Connat JL. An electron-microscopic study of smooth muscle cell dye coupling in the pig coronary arteries. Role of gap junctions. Circ Res 1992;70:49 – 55.
[27] Li X, Simard JM. Multiple connexins form gap junction channels in rat basilar artery smooth muscle cells. Circ Res 1999;84:1277 – 84. [28] Nakamura K, Inai T, Shibata Y. Distribution of gap junction protein connexin 37 in smooth muscle cells of the rat trachea and pulmonary artery. Arch Histol Cytol 1999;62:27 – 37.
[29] Cai WJ, Koltai S, Kocsis E, et al. Connexin 37, not Cx40 and Cx43, is induced in vascular smooth muscle cells during coronary arterio-genesis. J Mol Cell Cardiol 2001;33:957 – 67.
[30] Li X, Simard JM. Connexin 45 gap junction channels in rat cerebral vascular smooth muscle cells. Am J Physiol 2001;281:H1890 – 8. [31] Rummery NM, Hickey H, McGurk G, et al. Connexin 37 is the
major connexin expressed in the media of caudal artery. Arterioscler Thromb Vasc Biol 2002;22:1427 – 32.
[32] Ko YS, Coppen SR, Dupont E, et al. Regional differentiation of desmin, connexin 43, and connexin 45 expression patterns in rat aortic smooth muscle. Arterioscler Thromb Vasc Biol 2001;21: 355 – 64.
[33] Pacicca C, Schaad O, Beny JL. Electrotonic propagation of kinin-induced, endothelium-dependent hyperpolarizations in pig coronary smooth muscles. J Vasc Res 1996;33:380 – 5.
[34] Sandow SL, Hill CE. Incidence of myoendothelial gap junctions in the proximal and distal mesenteric arteries of the rat is suggestive of a role in endothelium-derived hyperpolarizing factor-mediated re-sponses. Circ Res 2000;86:341 – 6.
[35] Emerson GG, Segal SS. Electrical coupling between endothelial cells and smooth muscle cells in hamster feed arteries: role in vaso-motor control. Circ Res 2000;87:474 – 9.
[36] Xia J, Little TL, Duling BR. Cellular pathways of the conducted electrical response in arterioles of hamster cheek pouch in vitro. Am J Physiol 1995;269:H2031 – 8.
[37] Welsh DG, Segal SS. Endothelial and smooth muscle cell conduc-tion in arterioles controlling blood flow. Am J Physiol 1998;274: H178 – 86.
[38] Dora KA. Cell – cell communication in the vessel wall. Vasc Med 2001;6:43 – 50.
[39] Beny JL. Information networks in the arterial wall. News Physiol Sci 1999;14:68 – 73.
[40] von der Weid PY, Beny JL. Simultaneous oscillations in the mem-brane potential of pig coronary artery endothelial and smooth muscle cells. J Physiol 1993;471:13 – 24.
[41] Beny JL, Zhu P, Haefliger IO. Lack of bradykinin-induced smooth muscle cell hyperpolarization despite heterocellular dye coupling and endothelial cell hyperpolarization in porcine ciliary artery. J Vasc Res 1997;34:344 – 50.
[42] Schuster A, Oishi H, Beny JL, et al. Simultaneous arterial calcium dynamics and diameter measurements: application to myoendothe-lial communication. Am J Physiol, Heart Circ Physiol 2001;280: H1088 – 96.
[43] Edwards G, Weston AH. Endothelium-derived hyperpolarizing fac-tor—a critical appraisal. Prog Drug Res 1998;50:107 – 33. [44] Feletou M, Vanhoutte PM. The third pathway:
endothelium-depen-dent hyperpolarization. J Physiol Pharmacol 1999;50:525 – 34. [45] Campbell WB, Harder DR. Prologue: EDHF—what is it? Am J
Physiol, Heart Circ Physiol 2001;280:H2413 – 6.
[46] Campbell WB, Gauthier KM. What is new in endothelium-derived hyperpolarizing factors? Curr Opin Nephrol Hypertens 2002;11: 177 – 83.
[47] McGuire JJ, Ding H, Triggle CR. Endothelium-derived relaxing factors: a focus on endothelium-derived hyperpolarizing factor(s). Can J Physiol Pharmacol 2001;79:443 – 70.
myoendo-thelial gap junctions in the actions of endothelium-derived hyper-polarizing factor. Circ Res 2002;90:1108 – 13.
[49] Chaytor AT, Martin PE, Edwards DH, et al. Gap junctional commu-nication underpins EDHF-type relaxations evoked by ACh in the rat hepatic artery. Am J Physiol 2001;280:H2441 – 50.
[50] Hill CE, Phillips JK, Sandow SL. Heterogeneous control of blood flow amongst different vascular beds. Med Res Rev 2001; 21:1 – 60.
[51] Yamamoto Y, Imaeda K, Suzuki H. Endothelium-dependent hyper-polarization and intercellular electrical coupling in guinea-pig mes-enteric arterioles. J Physiol 1999;514(Pt 2):505 – 13.
[52] Beny J. Electrical coupling between smooth muscle cells and endo-thelial cells in pig coronary arteries. Pflugers Arch 1997;433:364 – 7. [53] Davies PF. Flow-mediated endothelial mechanotransduction. Physiol
Rev 1995;75:519 – 60.
[54] Christ GJ, Spray DC, el-Sabban M, et al. Gap junctions in vascular tissues. Evaluating the role of intercellular communication in the modulation of vasomotor tone. Circ Res 1996;79:631 – 46. [55] Bartlett IS, Crane GJ, Neild TO, et al. Electrophysiological basis of
arteriolar vasomotion in vivo. J Vasc Res 2000;37:568 – 75. [56] Bartlett IS, Segal SS. Resolution of smooth muscle and endothelial
pathways for conduction along hamster cheek pouch arterioles. Am J Physiol Heart Circ Physiol 2000;278:H604 – 12.
[57] Cowan DB, Lye SJ, Langille BL. Regulation of vascular connexin 43 gene expression by mechanical loads. Circ Res 1998;82:786 – 93. [58] DePaola N, Davies PF, Pritchard Jr WF, et al. Spatial and temporal regulation of gap junction connexin 43 in vascular endothelial cells exposed to controlled disturbed flows in vitro. Proc Natl Acad Sci 1999;96:3154 – 9.
[59] Severs NJ, Rothery S, Dupont E, et al. Immunocytochemical anal-ysis of connexin expression in the healthy and diseased cardiovas-cular system. Microsc Res Tech 2001;52:301 – 22.
[60] Haefliger JA, Castillo E, Waeber G, et al. Hypertension increases connexin 43 in a tissue-specific manner. Circulation 1997;95: 1007 – 14.
[61] Haefliger JA, Meda P, Formenton A, et al. Aortic connexin 43 is decreased during hypertension induced by inhibition of nitric oxide synthase. Arterioscler Thromb Vasc Biol 1999;19:1615 – 22. [62] Haefliger JA, Bergonzelli G, Waeber G, et al. Renin and angiotensin
II receptor gene expression in kidneys of renal hypertensive rats. Hypertension 1995;26:733 – 7.
[63] Haefliger JA, Demotz S, Braissant O, et al. Connexins 40 and 43 are differentially regulated within the kidneys of rats with renovascular hypertension. Kidney Int 2001;60:190 – 201.
[64] Yeh HI, Lupu F, Dupont E, et al. Upregulation of connexin 43 gap junctions between smooth muscle cells after balloon catheter injury in the rat carotid artery. Arterioscler Thromb Vasc Biol 1997;17: 3174 – 84.
[65] Bastide B, Neyses L, Ganten D, et al. Gap junction protein connexin 40 is preferentially expressed in vascular endothelium and conduc-tive bundles of rat myocardium and is increased under hypertensive conditions. Circ Res 1993;73:1138 – 49.
[66] Hong T, Hill CE. Restricted expression of the gap junctional protein connexin 43 in the arterial system of the rat. J Anat 1998;192(Pt 4): 583 – 93.
[67] Tsai ML, Watts SW, Loch-Caruso R, et al. The role of gap junctional communication in contractile oscillations in arteries from normoten-sive and hypertennormoten-sive rats. J Hypertens 1995;13:1123 – 33. [68] Rummery NM, McKenzie KU, Whitworth JA, et al. Decreased
en-dothelial size and connexin expression in rat caudal arteries during hypertension. J Hypertens 2002;20:247 – 53.
[69] Li X, Simard JM. Increase in Cx45 gap junction channels in cerebral smooth muscle cells from SHR. Hypertension 2002;40:940 – 6. [70] Liao Y, Day KH, Damon DN, et al. Endothelial cell-specific
knock-out of connexin 43 causes hypotension and bradycardia in mice. Proc Natl Acad Sci U S A 2001;98:9989 – 94.
[71] Theis M, de Wit C, Schlaeger TM, et al. Endothelium-specific
re-placement of the connexin 43 coding region by a lacZ reporter gene. Genesis 2001;29:1 – 13.
[72] Kruger O, Plum A, Kim JS, et al. Defective vascular development in connexin 45-deficient mice. Development 2000;127:4179 – 93. [73] Kirchhoff S, Nelles E, Hagendorff A, et al. Reduced cardiac
con-duction velocity and predisposition to arrhythmias in connexin 40-deficient mice. Curr Biol 1998;8:299 – 302.
[74] Simon AM, Goodenough DA, Paul DL. Mice lacking connexin 40 have cardiac conduction abnormalities characteristic of atrio-ventricular block and bundle branch block. Curr Biol 1998;8: 295 – 8.
[75] de Wit C, Roos F, Bolz SS, et al. Impaired conduction of vaso-dilation along arterioles in connexin 40-deficient mice. Circ Res 2000;86:649 – 55.
[76] de Wit C, Roos F, Bolz SS, et al. Lack of vascular connexin 40 is associated with hypertension and irregular arteriolar vasomotion. Physiol Genomics 2003;13:169 – 77.
[77] White TW, Bruzzone R, Paul DL. The connexin family of intercel-lular channel forming proteins. Kidney Int 1995;48:1148 – 57. [78] Boll HU, Forssmann WG, Taugner R. Studies on the
juxtaglomeru-lar apparatus: IV. Freeze-fracturing of membrane surfaces. Cell Tis-sue Res 1975;161:459 – 69.
[79] Mink D, Schiller A, Kriz W, et al. Interendothelial junctions in kidney vessels. Cell Tissue Res 1984;236:567 – 76.
[80] Barajas L, Liu L, Tucker M. Localization of connexin 43 in rat kidney. Kidney Int 1994;46:621 – 6.
[81] Guo R, Liu L, Barajas L. RT-PCR study of the distribution of con-nexin 43 mRNA in the glomerulus and renal tubular segments. Am J Physiol 1998;275:R439 – 47.
[82] Seul H, Beyer EC. Heterogeneous localization of connexin 40 in the renal vasculature. Microvasc Res 2000;59:140 – 8.
[83] Arensbak B, Mikkelsen HB, Gustafsson F, et al. Expression of con-nexin 37, 40, and 43 mRNA and protein in renal preglomerular arterioles. Histochem Cell Biol 2001;115:479 – 87.
[84] Ryan MJ, Liu B, Herbowy MT, et al. Intercellular communication between renin expressing As4.1 cells, endothelial cells and smooth muscle cells. Life Sci 2003;72:1289 – 301.
[85] Steinhausen M, Endlich K, Nobiling R, et al. Electrically induced vasomotor responses and their propagation in rat renal vessels in vivo. J Physiol 1997;505(Pt 2):493 – 501.
[86] Wagner AJ, Holstein-Rathlou NH, Marsh DJ. Internephron coupling by conducted vasomotor responses in normotensive and spontane-ously hypertensive rats. Am J Physiol 1997;272:F372 – 9. [87] Spray DC. Gap junction proteins: where they live and how they die.
Circ Res 1998;83:679 – 81.
[88] Haefliger JA, Tissieres P, Tawadros T, et al. Connexins 43 and 26 are differentially increased after rat bladder outlet obstruction. Exp Cell Res 2002;274:216 – 25.
[89] Christ GJ, Day NS, Day M, et al. Increased connexin 43-mediated intercellular communication in a rat model of bladder overactivity in vivo. Am J Physiol 2003;284:R1241 – 8.
[90] Neuhaus J, Wolburg H, Hermsdorf T, et al. Detrusor smooth muscle cells of the guinea-pig are functionally coupled via gap junctions in situ and in cell culture. Cell Tissue Res 2002;309:301 – 11. [91] Neuhaus J, Weimann A, Stolzenburg JU, et al. Smooth muscle cells
from human urinary bladder express connexin 43 in vivo and in vitro. World J Urol 2002;20:250 – 4.
[92] Fry CH, Cooklin M, Birns J, et al. Measurement of intercellular electrical coupling in guinea-pig detrusor smooth muscle. J Urol 1999;161:660 – 4.
[93] Hashitani H, Fukuta H, Takano H, et al. Origin and propagation of spontaneous excitation in smooth muscle of the guinea-pig urinary bladder. J Physiol 2001;530:273 – 86.
[94] Chien S, Li S, Shyy YJ. Effects of mechanical forces on signal transduction and gene expression in endothelial cells. Hypertension 1998;31:162 – 9.
p60src-Ras-MAPK signaling pathways in vascular endothelial cells. Arterioscler Thromb Vasc Biol 1998;18:227 – 34.
[96] Riser BL, Cortes P, Yee J. Modelling the effects of vascular stress in mesangial cells. Curr Opin Nephrol Hypertens 2000;9:43 – 7. [97] Persson K, Dean-Mckinney T, Steers WD, et al. Activation of the
transcription factors nuclear factor-kappaB and activator protein-1 in bladder smooth muscle exposed to outlet obstruction and mechanical stretching. J Urol 2001;165:633 – 9.
[98] Wu JJ, Geimonen E, Andersen J. Increased expression of estrogen
receptor beta in human uterine smooth muscle at term. Eur J Endo-crinol 2000;142:92 – 9.
[99] Dodge SM, Beardslee MA, Darrow BJ, et al. Effects of angio-tensin II on expression of the gap junction channel protein con-nexin 43 in neonatal rat ventricular myocytes. J Am Coll Cardiol 1998;32:800 – 7.
[100] Nguyen HT, Adam RM, Bride SH, et al. Cyclic stretch activates p38 SAPK2-, ErbB2-, and AT1-dependent signaling in bladder smooth muscle cells. Am J Physiol 2000;279:C1155 – 67.