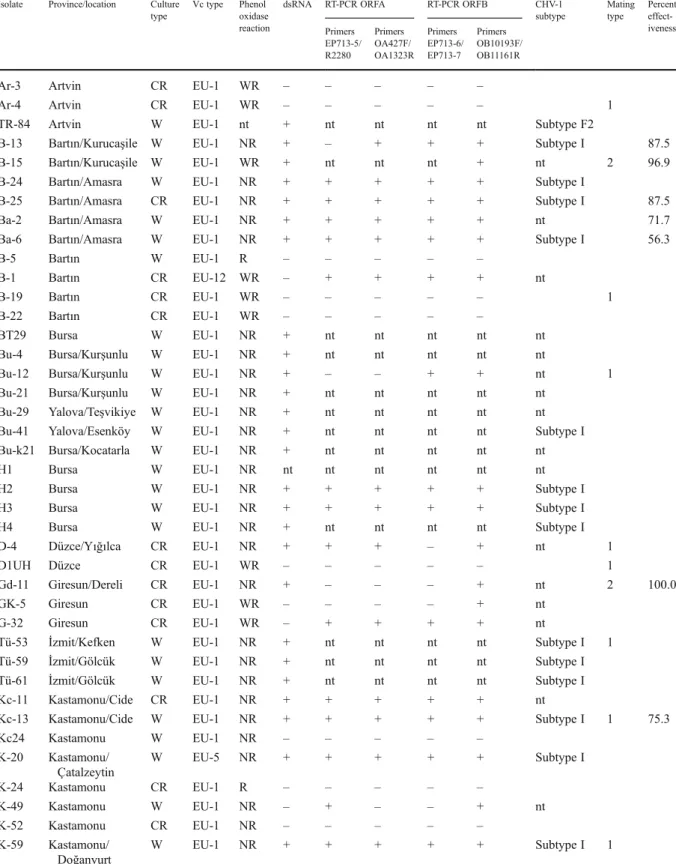
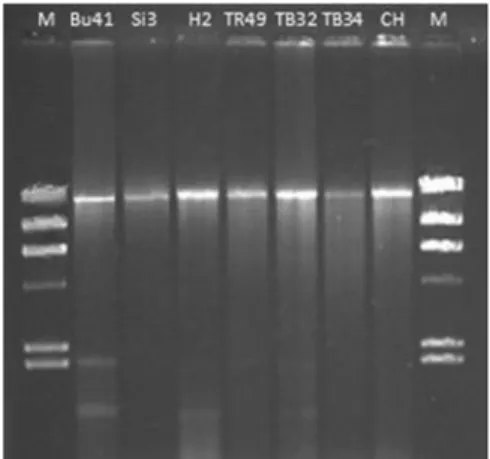
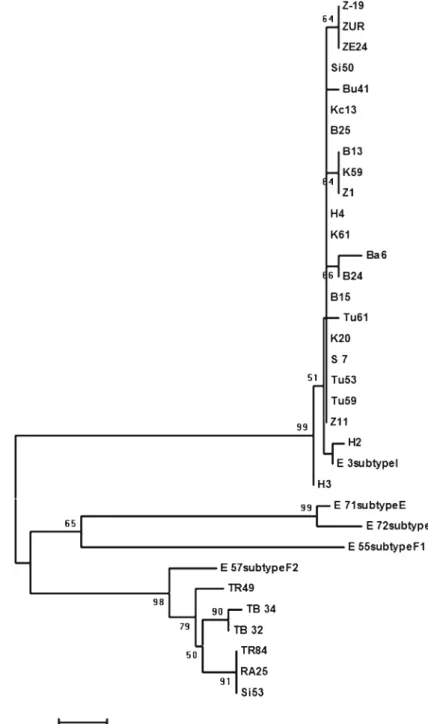
Characterization of hypovirulent isolates of the chestnut blight fungus, Cryphonectria parasitica from the Marmara and Black Sea regions of Turkey
12
0
0
Texte intégral
Figure
Documents relatifs