Les effets du préconditionnement ischémique sur
l’oxygénation et la force musculaire
Mémoire
Pénélope Paradis-Deschênes
Maîtrise en kinésiologie
Maître ès sciences (M. Sc.)
Québec, Canada
© Pénélope Paradis-Deschênes, 2015
iii
Résumé
Le préconditionnement ischémique (IPC) consiste à induire des épisodes ischémiques brefs et répétés, entrecoupés d’épisodes de reperfusion. Cette technique initialement testée pour ses effets protecteurs sur le myocarde rendrait différents tissus du corps plus résistants à une réduction d’oxygénation, comme retrouvée durant l’exercice maximal.
Très peu d’études ont étudié les effets de l’IPC sur l’oxygénation musculaire. À l’aide de la spectroscopie proche infrarouge, nous avons examiné les effets du préconditionnement sur l’hémodynamique et l’extraction d’oxygène pendant un protocole de contractions volontaires maximales répétées.
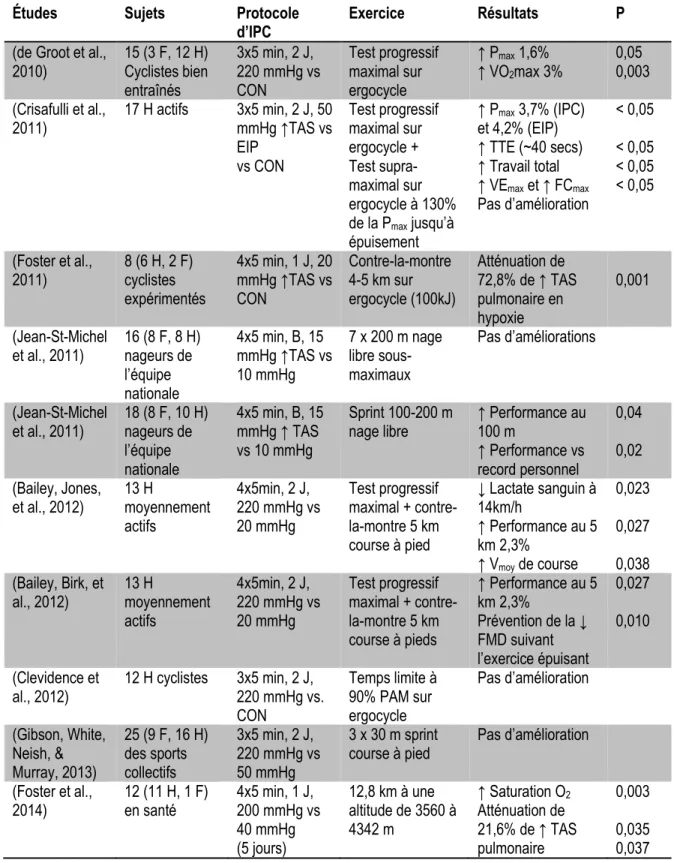
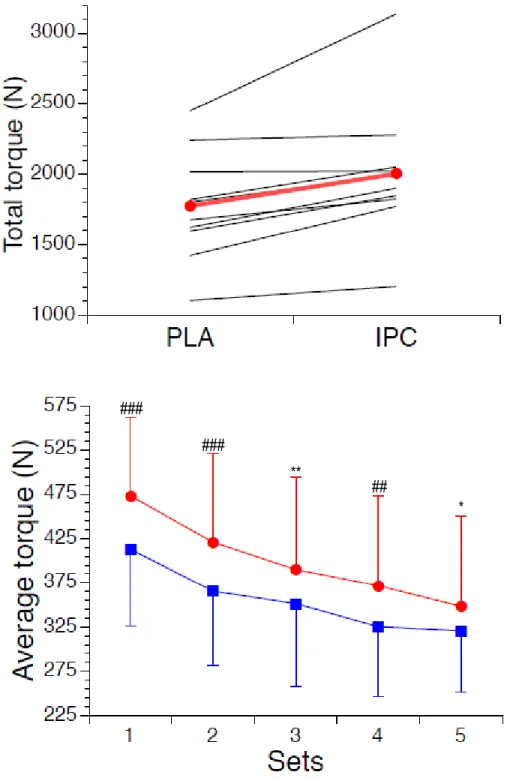
Nos résultats démontrent que ce préconditionnement entraîne une augmentation du volume sanguin local au repos et pendant la récupération après les contractions, ainsi qu’une plus grande extraction d’oxygène en début d’effort. Ces améliorations s’accompagnent d’une augmentation de la force musculaire tout au long du protocole.
L’utilisation de l’IPC comme aide ergogénique pourrait s’avérer utile pour optimiser l’oxygénation et la force musculaire avant une compétition ou un entraînement.
v
Abstract
Repeated episodes of ischemia and reperfusion, commonly known as ischemic preconditioning (IPC), initially tested for its protective effects on the myocardium, renders different tissues more resistant to a subsequent reduction in oxygenation (i.e., tissue O2 saturation), as found during maximal exercise.
Despite its likely effects on the oxidative function, studies examining the effects of IPC on tissue oxygenation are very scarce. We used near-infrared spectroscopy to study the effects of IPC on muscle haemodynamics and peripheral O2 extraction during repeated maximal efforts in resistance-trained participants.
Our results demonstrate that IPC increased muscle blood volume at rest and during recovery between contractions, as well as muscle oxygen uptake during exercise. These acute physiological adaptations were associated with increased muscle strength throughout the protocol.
The use of IPC as an ergogenic aid could be useful to optimize oxygenation and muscle strength during competition and training.
vii
Table des matières
Résumé ... iii
Abstract ... v
Table des matières ... vii
Liste des tableaux ... ix
Liste des illustrations ... xi
Liste des abréviations... xii
Remerciements ... xv
Avant-propos ... xvii
Introduction ... 1
Chapitre 1 : Revue de littérature ... 3
1.1 Historique de l’IPC ... 3
1.2 Protocoles d’application de l’IPC ... 5
1.3 IPC et performance sportive ... 6
1.4 Mécanismes d’actions de l’IPC ... 10
1.4.1 Déclencheurs de l’IPC ... 11
1.4.2 Médiateurs et effecteurs finaux de l’IPC ... 15
1.5 IPC et fonction musculaire ... 17
1.6 IPC et fonction endothéliale ... 18
1.7 IPC et fonction oxydative ... 20
1.8 Spectroscopie proche infrarouge (NIRS) ... 22
Chapitre 2 : Problématique ... 25
Chapitre 3 : Article scientifique ... 27
3.1 ABSTRACT ... 29 3.2 INTRODUCTION ... 30 3.3 METHODS ... 31 3.3.1 Participants ... 31 3.3.2 Experimental design ... 31 3.3.3 Exercise protocol ... 32 3.3.4 Near-infrared spectroscopy ... 33 3.3.5 Statistical analysis ... 34 3.4 RESULTS ... 34
viii
3.4.1 Performance ... 34
3.4.2 Muscle haemodynamics and oxygenation ... 35
3.5 DISCUSSION ... 35 3.6 REFERENCES ... 38 3.7 TABLE ... 43 3.8 FIGURES ... 44 Chapitre 4 : Discussion ... 49 Conclusion ... 53 Bibliographie ... 55
ix
Liste des tableaux
Tableau 1-1 : Effets de l’IPC sur la performance... 7 Tableau 3-1 : Performance, muscle haemodynamics and oxygenation measures for IPC and SHAM conditions during the five sets. ... 43
xi
Liste des illustrations
Figure 1-1 : Représentation schématique des deux phases de protection du préconditionnement ischémique . 3
Figure 1-2 Schéma résumé des mécanismes de l'IPC... 17
Figure 1-3: Détecteur du NIRS et son champ d'émission de rayons infrarouges ... 22
Figure 1-4: Signaux de la [HbO2] en rouge, de la [HHb] en bleu et de la [THb] en vert. ... 23
Figure 3-1 :Total torque and average torque ... 44
Figure 3-2. Change in resting muscle blood volume amplitude (∆[THb]) from baseline to post- SHAM and IPC conditions. ... 46
Figure 3-3 Changes from rest in muscle O2 extraction during exercise and muscle blood volume during recovery. ... 47
xii
Liste des abréviations
ATP : adénosine triphosphateB : bras
Canaux KATP : canaux potassium sensible à l’ATP CON : condition contrôle
CGRP : peptide lié au gène de la calcitonine
EIP : préconditionnement ischémique précédé d’un exercice
eNOS : isoforme endothélial de la synthase d’oxyde nitrique
EPCs : Cellules progénitrices endothéliales
F : femme
FCmax : fréquence cardiaque maximale
FMD : vasodilatation dépendante du flux sanguin
H : homme
[HbO2]: concentrations micromolaires de l’oxyhémoglobine [HHb]: concentrations micromolaires de la désoxyhémoglobine
HIF-1α : facteur de transcription inductible par hypoxie
IPC : préconditionnement ischémique, peut être direct (invasif) ou indirect (non invasif)
J : jambe
Min : minutes
mPTP : pores de perméabilité transitionnelle de la mitochondrie
xiii NO : oxyde nitrique
O2 : oxygène
[O2Hb]: concentrations de l’oxyhémoglobine PAM : puissance aérobie maximale
PCr : phosphocréatine
PKC: protéine kinase C
Pmax : puissance maximale Pmoy : puissance moyenne Ptot : puissance totale
RIPC : préconditionnement ischémique effectué à distance de l’organe cible, peut-être direct (invasif) ou indirect (non invasif)
ROS : dérivés réactifs de l’oxygène
Secs : secondes
TAD : tension artérielle diastolique
TAS : tension artérielle systolique
[THb]: concentrations micromolaires de l’hémoglobine totale
TSI : saturation en oxygène
TTE : temps total d’exercice
VEGF : facteur de croissance vasculaire endothélial
VEmax : ventilation maximale Vmoy : vitesse moyenne
xiv
VO2max : consommation maximale d’oxygène
xv
Remerciements
En tout premier lieu, j’aimerais remercier mon directeur de recherche, François Billaut. C’est grâce à ta passion contagieuse que j’ai fait le grand saut pour la maîtrise avec confiance et je t’en serai toujours reconnaissante. Merci pour l’apprentissage transmis, les occasions offertes, ta confiance et ton support tout au long de la réalisation de ce projet. Merci également pour toutes ces discussions qui m’ont apporté la compréhension, mais surtout des questionnements constants.
Merci à mon codirecteur Denis R. Joanisse. Merci pour ta présence, ton aide et ton expérience qui m’ont permis de mener ce projet à terme. La réalisation de cette maîtrise aurait été beaucoup plus difficile sans ton soutien constant et ta confiance en ce projet.
Un autre merci à l’équipe du GRAME et tout particulièrement à Normand Teasdale et Marcel Kazap. Merci, Normand, de m’avoir donné ce premier contact avec la recherche au baccalauréat et merci pour les questionnements et les connaissances transmises en laboratoire. Marcel, merci pour ta patience infinie et ton aide dans la calibration et le traitement des données.
J’aimerais également remercier Patrice Brassard pour son support dans ce projet et pour sa passion débordante en la recherche.
Merci à mes collègues de bureau qui m’ont accompagné et conseillé tout au long de ce processus.
Enfin, merci à ma famille qui m’a toujours supporté dans mes projets. Merci pour votre amour, vos encouragements et votre appui depuis toujours. Bien sûr, merci à mon amoureux Maxime de toujours être présent et de m’avoir soutenu tout au long de cette aventure.
xvii
Avant-propos
L’article inséré dans ce mémoire a été rédigé en collaboration avec Denis R Joanisse et François Billaut. L’article est présentement en phase finale de rédaction pour être soumis pour publication à la revue indexée
International Journal of Sports Medicine. Cet article est le résultat concret de mon projet de maîtrise. J’ai agi
comme coordonnatrice du projet, accomplissant toutes ses étapes (rédaction du protocole de recherche, obtention du certificat d’éthique, recrutement des participants, expérimentations, analyse des données et rédaction). Je suis donc l’auteure principale de cet article.
François Billaut, Ph. D., professeur-chercheur au Département de kinésiologie de l’Université Laval et chercheur au Centre de recherche de l’Institut universitaire de cardiologie et de pneumologie de Québec, est le directeur de mes travaux de maîtrise. Il a contribué grandement à chacune des étapes du projet, plus particulièrement à l’élaboration du protocole de recherche et à la révision de l’article.
Denis R Joanisse, Ph. D. est professeur-chercheur au Département de kinésiologie de l’Université Laval et chercheur au Centre de recherche de l’Institut universitaire de cardiologie et de pneumologie de Québec. Il a agi comme codirecteur de mes travaux de maîtrise, en participant à l’élaboration du protocole de recherche et à la révision critique de l’article.
1
Introduction
Plusieurs stratégies de conditionnement physique ont été examinées pour augmenter et maintenir l’apport en oxygène (O2) et ainsi parées à l’augmentation considérable des exigences observées à l’exercice physique maximal. Le préconditionnement ischémique, soit l’utilisation d’épisodes brefs d’ischémie (5 minutes) et répétés, entrecoupées d’épisodes de reperfusion sanguine, rendrait différents tissus du corps (incluant le muscle squelettique) plus résistants à une réduction subséquente d’oxygénation, telle que retrouvée au cours de l’exercice (Berger, Macholz, Mairbaurl, & Bartsch, 2015; Tapuria et al., 2008). Les adaptations moléculaires qui découlent de l’IPC (entre autres la libération de métabolites, de dérivés réactifs à l’O2 et d’oxyde nitrique, l’activation de la protéine kinase C et ouverture des canaux potassium sensible à l’adénosine triphosphate) entraîneraient une vasodilatation, une augmentation du flux sanguin, et finalement, une amélioration de l’apport en O2 ainsi qu’une épargne d’adénosine triphosphate (ATP) (Beaven, Cook, Kilduff, Drawer, & Gill, 2012; Lochner, Marais, Du Toit, & Moolman, 2002; Tapuria et al., 2008). Toutefois, aucune étude n'a examiné l'impact de l'IPC sur la réponse oxydative musculaire, par exemple, l'extraction d’O2, durant un exercice d’intensité maximal. Malgré la popularité de l’IPC au sein de la population sportive, l’impact réel de cette technique sur la stimulation de l’extraction d’O2 demeure indéterminé. Pour ce faire, la spectroscopie proche à infrarouge (NIRS) du muscle vaste latéral est prometteuse, car elle permet d’examiner l’activation de la composante périphérique pour l’extraction d’O2.
Plusieurs études se sont intéressées aux impacts de l’IPC sur la performance et ont démontré son potentiel pour améliorer la fonction musculaire et la production de force chez l’humain (Crisafulli et al., 2011; Jean-St-Michel et al., 2011). Toutefois, toutes n’arrivent pas à la même conclusion, la grande variété de protocoles utilisés (c.-à-d. mode d’exercices) pourrait être à l’origine de cette confusion. Trois à quatre cycles d’IPC augmentent la puissance durant un test maximal progressif sur ergocycle (Crisafulli et al., 2011; de Groot, Thijssen, Sanchez, Ellenkamp, & Hopman, 2010), la distance parcourue sous l’eau (Kjeld, Rasmussen, Jattu, Nielsen, & Secher, 2014), ainsi que la performance lors d’un contre-la-montre de 5 km et d’un 100 m sprint en natation (Bailey, Birk, et al., 2012; Jean-St-Michel et al., 2011). À l’inverse, l’IPC n’entraîne pas d’amélioration de la puissance durant un test de Wingate de 30 secondes (Lalonde & Curnier, 2015), et peut même entraîner des effets négatifs lorsque celui-ci est répété (Paixao, da Mota, & Marocolo, 2014). De toute évidence, d'autres expériences sont nécessaires pour comprendre les effets physiologiques de cette méthode de préconditionnement ainsi que l’impact du mode de l'exercice sur son efficacité.
Ce mémoire de maîtrise fait tout d’abord le point sur les connaissances actuelles concernant les effets aigus de l’IPC sur les fonctions physiologiques et les performances lors d’exercices de durée et d’intensité variées,
2
et apporte une contribution aux connaissances dans ce domaine. L’étude présentée ici détermine si l’IPC améliore l’hémodynamique musculaire et l’extraction d’O2 durant des efforts maximaux répétés entrecoupés de périodes de récupération incomplètes. Des contractions isocinétiques à basses vitesses ont été utilisées pour entraver la circulation sanguine et ainsi fournir de nouvelles données sur l’impact réel de l’IPC durant l’effort.
3
Chapitre 1 : Revue de littérature
1.1 Historique de l’IPC
L’IPC a été présenté pour la première fois en 1986 par le docteur Murry et ses collègues (Murry, Jennings, & Reimer, 1986). Ils ont démontré, chez le chien, que l’occlusion à même l’artère coronaire durant quatre cycles de 5 minutes, entrecoupés de 5 minutes de reperfusion, permettait de diminuer la taille de l’infarctus provoqué par une ischémie du myocarde subséquente prolongée (40 minutes) (Murry et al., 1986). Cet effet de « protection » induit par l’IPC contre les atteintes cardiaques d’ischémie/reperfusion a ensuite été reproduit chez différentes espèces puis sur différents organes, incluant le muscle squelettique (Kharbanda, Nielsen, & Redington, 2009; Kocman et al., 2015).Ce conditionnement physiologique serait une réponse adaptative au stress causé par le manque O2, rendant le corps plus tolérant à la fois à l’ischémie, qui provoque la destruction des cellules, et à la reperfusion, qui entraîne autant de dommages en déclenchant la réponse inflammatoire (Blaisdell, 2002). La durée de la protection immédiate engendrée par cette intervention, également nommée préconditionnement classique ou précoce, pourrait s’étendre de quelques minutes suivant l’IPC jusqu’à 5 heures (Berger, Kohne, et al., 2015) (Figure 1). En 1993, une étude de Marber et coll. (Marber, Latchman, Walker, & Yellon, 1993) démontre l’existence d’une deuxième phase de protection de l’IPC, appelée phase de préconditionnement tardif, qui agirait 12 à 24 heures plus tard, et dont les effets perdureraient 3 à 4 jours suivant la manœuvre. À noter que le délai entre les deux phases ne semble procurer aucune protection supplémentaire (Bolli, 2000) et que la deuxième phase aurait des effets moins importants, mais plus longs et plus constants (Baxter, Goma, & Yellon, 1997).
Figure 1-1 : Représentation schématique des deux phases de protection du préconditionnement ischémique (Berger, Macholz, et al., 2015).
4
Dans la même année, Przyklenk et ses collègues (Przyklenk, Bauer, Ovize, Kloner, & Whittaker, 1993) montrent que l’IPC de l’artère circonflexe chez le chien réduit également l’infarctus mesuré à l’artère interventriculaire antérieure. Cette étude démontre alors l’existence du préconditionnement intraorganes et met en perspective la possibilité que l’IPC puisse induire une protection systémique. Ce phénomène physiologique, appelé le « remote ischemic preconditioning » (RIPC) ou préconditionnement à distance, est élégamment corroboré dans une étude rapportant que l’ischémie transitoire aux jambes induit une protection au cœur similaire à celle effectuée par ischémie locale (Birnbaum, Hale, & Kloner, 1997).
Toutefois, malgré ces découvertes, l’application de l’IPC est demeurée limitée puisque l’intervention s’effectuait directement sur les vaisseaux sanguins (intervention invasive). C’est seulement en 2001 que MacAllister et son équipe présentent une étude qui démontre que l’induction de l’IPC de façon non invasive (c.-à-d., exécutée par la compression d’un brassard à tension artérielle autour du bras à raison de 3 cycles de 5 minutes chacun), avant une ischémie prolongée de 20 minutes au bras, atténue le dysfonctionnement endothélial et l’activation systémique des neutrophiles (Kharbanda et al., 2001). La même équipe démontra ensuite que la protection induite par cette technique a elle aussi deux phases temporelles distinctes (Loukogeorgakis et al., 2005). Enfin, en 2006, Cheung et ses collègues (Cheung et al., 2006) publient la première application clinique de l’IPC en appliquant quatre cycles d’IPC aux jambes d’enfants avec un défaut congénital au cœur et devant être opérés. Dans cette étude, l’IPC a eu pour effet de moduler encore une fois la réponse inflammatoire et de réduire l’incidence contre des atteintes cardiaque et pulmonaire d’ischémie/reperfusion suivant l’intervention chirurgicale (Cheung et al., 2006).
Ainsi, l’IPC peut donc s’appliquer selon différentes combinaisons : directement (c.-à-d., méthode invasive) ou indirectement par un brassard à tension artérielle, localement, si exécuté à même l’organe cible, et à distance. Ces combinaisons semblent toutes entraîner deux phases de protection qui produisent des effets locaux et systémiques (Kocman et al., 2015; Loukogeorgakis et al., 2005). Néanmoins, cette revue de littérature se concentre sur l’IPC indirect par l’ischémie transitoire du muscle squelettique en raison de son côté sécuritaire et facilement applicable chez le sportif.
5
1.2 Protocoles d’application de l’IPC
De façon générale, l’IPC consiste à alterner des cycles d’ischémie et de reperfusion d’environ 5 minutes chacun, par la compression et le relâchement d’un brassard à tension artérielle positionné autour d’un ou des membres inférieurs ou supérieurs. Jusqu’à présent, la séquence idéale à utiliser pour optimiser les réponses physiologiques à l’exercice et la contraction musculaire n’a pas encore été identifiée.
Pour entraîner des effets bénéfiques, l’IPC doit permettre l’atteinte d’un certain « seuil » de stimulation physiologique, seuil qui varie entre les espèces et les parties, ou organe, du corps (Goto et al., 1995). Ainsi, on a identifié la durée et le type de préconditionnement comme facteurs influençant l’efficacité de l’IPC. La durée fait référence aux nombres et à la durée de chaque cycle d’ischémie. Les travaux dans ce domaine tendent à montrer que l‘efficacité de l’IPC serait corrélée avec le nombre de cycles de préconditionnement (Birnbaum et al., 1997; Gurke, Marx, Sutter, Frentzel, Salm, et al., 1996). Gurke et ses collègues (Gurke, Marx, Sutter, Frentzel, Salm, et al., 1996) ont d’ailleurs démontré que l’augmentation de la contractilité était plus importante avec l’induction de trois cycles d’ischémie et de reperfusion comparativement à l’induction d’un seul, ou de deux cycles. L’inverse est aussi vrai puisque le terme hyperconditionnement commence à faire son apparition dans la littérature et réfère à l’exécution d’épisodes d’ischémie de manière excessive, nombre encore indéfini, qui pourrait être délétère plutôt que bénéfique (Whittaker & Przyklenk, 2014). Pour la durée des cycles, Mattei et ses collègues (Mattei et al., 2000) ajoutent que l’efficacité de l’IPC est plus dépendante du nombre de cycles que de la durée de ceux-ci. En modulant la durée des épisodes (2,5, 5 et 10 minutes) ils observent que les trois durées améliorent la fonction musculaire des fibres rapides après l’ischémie prolongée, mais que seul le protocole avec trois cycles d’ischémie et de reperfusion de 5 minutes chacun permet d’améliorer la fonction des fibres lentes.
Le type de préconditionnement fait référence à l’endroit d’application de l’IPC. En effet, la pression intramusculaire exercée causant la réduction du débit sanguin varie selon le membre (bras ou jambe) sur lequel le brassard est placé. On estime qu’une pression au bras de 30 mmHg au-dessus de la tension artérielle systolique (TAS) est suffisante pour bloquer l’apport sanguin artériel, alors qu’elle devrait être supérieure à 55 mmHg au-dessus de la TAS pour le membre inférieur, puisqu’une valeur absolue de 180 mmHg de pression peut parfois s’avérer insuffisante (Sharma, Cunniffe, Verma, Cardinale, & Yellon, 2014). La différence de pression efficace (c.-à-d., engendrant une réponse physiologique) peut s’expliquer par la différence de taille entre les membres supérieurs et inférieurs. En effet, des études ont précisé que la pression exercée aux jambes ne devrait pas être modulée en fonction de la TAS brachiale, mais plutôt selon la circonférence de la cuisse qui serait le facteur prédominant dans la détermination de la pression nécessaire (Loenneke et al., 2012; Loenneke et al., 2013). À ce jour, aucune étude n’a modulé la pression exercée en
6
fonction de la circonférence de cuisse pour examiner l’influence de ce facteur méthodologique sur l’efficacité des protocoles IPC.
En somme, même si la pression adéquate à utiliser reste méconnue, et est certainement influencée par la circonférence du membre, une durée totale d’ischémie d’au moins 15 minutes semble être optimale pour maximiser les effets physiologiques de l’IPC. Quoi qu’il en soit, en raison de ses effets sur les fonctions physiologiques, les effets ergogéniques potentiels de cette manœuvre sur la capacité d’exercice et la performance sportive ont été décrits à plusieurs reprises dans la littérature scientifique.
1.3 IPC et performance sportive
Les études s’intéressant aux effets de l’IPC sur les performances physiques et sportives sont décrites dans le Tableau 1. La plupart des études s’intéressant aux tests sous-maximaux (Clevidence, Mowery, & Kushnick, 2012; Jean-St-Michel et al., 2011) n’ont pas trouvé d’amélioration de la performance. Seule l’étude de Barbosa et ses collègues a démontré que l’IPC retardait l’apparition de la fatigue et prolongeait le temps de travail lors d’un protocole de répétitions de contractions sous-maximales jusqu’à épuisement. Les auteurs n’ont pas observé de changement au niveau du flux sanguin, mais dénotaient une plus grande désoxygénation de l’avant-bras (Barbosa et al., 2015). Dans le même ordre d’idées, Kjeld et son équipe rapportaient une augmentation de la distance parcourue sous l’eau (Kjeld et al., 2014), tandis que Foster et coll. rapportaient une atténuation de l’augmentation normale de la TAS pulmonaire d’environ 73% en hypoxie ainsi qu’une augmentation non significative de la performance au contre-la-montre (Foster, Westerdahl, Foster, Hsu, & Anholm, 2011). Foster et son équipe se sont également intéressé à l’effet cumulatif de l’IPC et ont rapporté une atténuation de la TAS pulmonaire, une augmentation de la saturation en O2 et de la performance en course (12,8 km) exécutée en altitude après l’application de l’IPC pendant cinq jours (Foster, Giri, Rogers, Larson, & Anholm, 2014). À l’inverse, Hittinger et coll. n’ont pas trouvé de différence entre l’IPC et son groupe contrôle (aucune intervention), lors de tests progressifs exécutés en normoxie et en hypoxie (Hittinger et al., 2015). Deux études rapportent une augmentation de la puissance maximale aérobie atteinte lors de tests progressifs sur ergocycle (Crisafulli et al., 2011; de Groot et al., 2010) après IPC comparativement à une condition contrôle. Les observations de Bailey et coll. (Bailey, Jones, et al., 2012) corroborent ces données en rapportant également une augmentation de puissance aérobie lors d’un contre-la-montre de 5 km (Bailey, Birk, et al., 2012; Bailey, Jones, et al., 2012). Les auteurs de ces études rapportent une diminution de l’accumulation de lactate sanguin à 14 km/h durant le test progressif précédant le contre-la-montre (Bailey, Jones, et al., 2012). Par contre, ces données ne font pas l’unanimité puisqu’un protocole d’IPC similaire n’a
7 pas permis d’améliorer la performance au contre-la-montre de 5 km sur piste chez des coureurs entraînés (Tocco et al., 2015).
Tableau 1-1 : Effets de l’IPC sur la performance
Études Sujets Protocole
d’IPC Exercice Résultats P
(de Groot et al.,
2010) 15 (3 F, 12 H) Cyclistes bien entraînés 3x5 min, 2 J, 220 mmHg vs CON Test progressif maximal sur ergocycle ↑ Pmax 1,6% ↑ VO2max 3% 0,05 0,003 (Crisafulli et al.,
2011) 17 H actifs 3x5 min, 2 J, 50 mmHg ↑TAS vs EIP vs CON Test progressif maximal sur ergocycle + Test supra-maximal sur ergocycle à 130% de la Pmax jusqu’à épuisement ↑ Pmax 3,7% (IPC) et 4,2% (EIP) ↑ TTE (~40 secs) ↑ Travail total ↑ VEmax et ↑ FCmax Pas d’amélioration < 0,05 < 0,05 < 0,05 < 0,05 (Foster et al., 2011) 8 (6 H, 2 F) cyclistes expérimentés 4x5 min, 1 J, 20 mmHg ↑TAS vs CON Contre-la-montre 4-5 km sur ergocycle (100kJ) Atténuation de 72,8% de ↑ TAS pulmonaire en hypoxie 0,001 (Jean-St-Michel et al., 2011) 16 (8 F, 8 H) nageurs de l’équipe nationale 4x5 min, B, 15 mmHg ↑TAS vs 10 mmHg 7 x 200 m nage libre sous-maximaux Pas d’améliorations (Jean-St-Michel et al., 2011) 18 (8 F, 10 H) nageurs de l’équipe nationale 4x5 min, B, 15 mmHg ↑ TAS vs 10 mmHg Sprint 100-200 m
nage libre ↑ Performance au 100 m ↑ Performance vs record personnel 0,04 0,02 (Bailey, Jones, et al., 2012) 13 H moyennement actifs 4x5min, 2 J, 220 mmHg vs 20 mmHg Test progressif maximal + contre-la-montre 5 km course à pied ↓ Lactate sanguin à 14km/h ↑ Performance au 5 km 2,3% ↑ Vmoy de course 0,023 0,027 0,038 (Bailey, Birk, et al., 2012) 13 H moyennement actifs 4x5min, 2 J, 220 mmHg vs 20 mmHg Test progressif maximal + contre-la-montre 5 km course à pieds ↑ Performance au 5 km 2,3% Prévention de la ↓ FMD suivant l’exercice épuisant 0,027 0,010 (Clevidence et
al., 2012) 12 H cyclistes 3x5 min, 2 J, 220 mmHg vs. CON Temps limite à 90% PAM sur ergocycle Pas d’amélioration (Gibson, White, Neish, & Murray, 2013) 25 (9 F, 16 H) des sports collectifs 3x5 min, 2 J, 220 mmHg vs 50 mmHg 3 x 30 m sprint
course à pied Pas d’amélioration (Foster et al., 2014) 12 (11 H, 1 F) en santé 4x5 min, 1 J, 200 mmHg vs 40 mmHg (5 jours) 12,8 km à une altitude de 3560 à 4342 m ↑ Saturation O2 Atténuation de 21,6% de ↑ TAS pulmonaire 0,003 0,035 0,037
8 ↑ Performance (Paixao et al., 2014) 15 cyclistes 4x5 min, 2 J, 250 mmHg vs CON 3 x 30-sec sprint
sur ergocycle ↓ Pmax au 1 er Wingate ↓Pmoy et Ptot anaérobie au Wingate 1 et 2 <0,05 <0,01 (Gibson, Mahony, Tracey, Fawkner, & Murray, 2015) 16 (9 F, 7 H) des sports collectifs 3x5 min, 2 J, 220 mmHg vs 50 mmHg 5 x 6-sec sprint sur ergocycle avec 30-sec de repos entre les séries
Pas d’amélioration
(Hittinger et al.,
2015) 15 H cyclistes et triathloniens 4x5 min, 2 J, 10-20 mmHg ↑TAS vs. CON 10 min à 55% Pmax + test progressif maximal Pas d’amélioration (Lalonde & Curnier, 2015) 17 (9 F, 8 H) Amateurs triathlons 4x5min, bras droit, 50 mmHg ↑TAS vs. 10 mmHg 6 x 6-sec sur ergocycle avec 2-min de repos entre les séries et test sprint de 30 secs sur ergocycle
Pas d’amélioration
(Tocco et al.,
2015) 11 H coureurs bien entraînés 3x5 min, 2 J, 50 mm Hg ↑TAS vs. 10 mm Hg ↓ TAD
Course de 5 km Pas d’amélioration
B : bras, CON : contrôle, EIP : IPC précédé d’un exercice, F : femmes, FCmax : fréquence cardiaque maximale, FMD : vasodilatation dépendante du flux sanguin, H : Hommes, IPC : préconditionnement ischémique, J : jambes, min : minutes, O2 : oxygène, PAM : puissance aérobie maximale, Pmax : puissance maximale, Pmoy : puissance moyenne, Ptot : puissance totale, Secs : secondes, TAD : Tension artérielle diastolique, TAS : tension artérielle systolique, TTE : temps total d’exercice, VEmax : ventilation maximale, Vmoy : vitesse moyenne, VO2max : consommation maximale d’oxygène
Les études portant sur les efforts courts et supra-maximaux (c.-à-d., d’intensité supérieure à la puissance aérobie maximale) sont moins nombreuses et il est difficile de tirer des conclusions quant à l’efficacité de l’IPC dans ces conditions d’exercice. Lorsqu’on a soumis des nageurs élites à un protocole d’occlusion de 4 cycles de 5 minutes, l’IPC a permis d’améliorer la performance lors de sprints de 100 et 200 m en natation (~60 secondes d’effort) (Jean-St-Michel et al., 2011). Lorsque les efforts sont plus courts, comme dans le cas de test anaérobie de Wingate de 30 secondes sur ergocycle, l’IPC ne semble pas avoir de résultat favorable par rapport à une condition contrôle. En effet, les puissances maximales et moyennes développées étaient similaires dans un cas (Lalonde & Curnier, 2015), et même plus faibles que celles développées lors de la condition contrôle lorsque les tests de Wingate étaient répétés avec 10 minutes de récupération (Paixao et al.,
9 2014). L’IPC a été inefficace pour induire des changements dans deux autres études exécutant des efforts supra-maximaux (Crisafulli et al., 2011; Lalonde & Curnier, 2015). Enfin, il en va de même pour des efforts dynamiques très courts. Par rapport à la condition contrôle, l’IPC n’a pas permis à des athlètes de sports collectifs d’améliorer leur puissance maximale et leur indice de fatigue lors de tests de répétitions de sprints comprenant 3 sprints en course à pied de 30 mètres (Gibson et al., 2013) et 5 sprints de 6 secondes sur ergocycle entrecoupés de 30 secondes de récupération passive (Gibson et al., 2015).
À la lumière de ces résultats, il semble que l’IPC soit ergogénique dans certaines conditions d’exercice. Cette technique de préconditionnement pourrait être plus avantageuse lors d’efforts intenses où l’apport en O2 est limité en raison de l’environnement (ex. : hypoxie, apnée) ou d’une limitation au niveau de la perfusion (Bailey, Jones, et al., 2012; Barbosa et al., 2015; de Groot et al., 2010; Foster et al., 2014; Jean-St-Michel et al., 2011; Kjeld et al., 2014). Toutefois, il est encore difficile de tirer des conclusions à partir des études actuelles. Les divergences pourraient entre autres s’expliquer par la grande variété des protocoles d’IPC utilisés dans les études, dont plusieurs pourraient être inadéquats d’un point de vue méthodologique. En effet, la pression utilisée dans les études varie de 10 mmHg au-dessus de la TAS à une valeur absolue de 250 mmHg (voir Tableau 1). Il est donc probable que certaines études n’aient pas utilisé une pression suffisante pour engendrer une occlusion des tissus et ainsi permettre d’optimiser l’IPC. À titre d’exemple, l’étude de Jean-St-Michel et coll. a utilisé une pression de 15 mmHg au-dessus de la TAS et le brassard de compression était placé sur les membres supérieurs d’athlètes, dont certains étaient mineurs et donc probablement moins corpulents (Jean-St-Michel et al., 2011). En comparaison, une autre étude a utilisé une pression similaire (15 mmHg au-dessus de la TAS), mais en appliquant le brassard sur les membres inférieurs des participants (Hittinger et al., 2015). L’absence de résultat positif dans cette étude pourrait donc s’expliquer par l’inefficacité de la pression choisie pour induire l’IPC (Hittinger et al., 2015). Le choix du protocole placébo peut aussi confondre les résultats. Certaines études utilisent une pression placébo de 10–20 mmHg (Bailey, Birk, et al., 2012; Bailey, Jones, et al., 2012; Jean-St-Michel et al., 2011; Lalonde & Curnier, 2015), alors que d’autres vont induire des pressions ≥50 mm Hg (Gibson et al., 2015; Gibson et al., 2013; Tocco et al., 2015). Or, une pression trop élevée, même si elle s’avère insuffisante pour optimiser l’IPC, peut tout de même induire des effets physiologiques qui viendraient amoindrir la différence entre le groupe IPC et placébo (Sumide, Sakuraba, Sawaki, Ohmura, & Tamura, 2009; Takarada et al., 2000). En plus des problèmes méthodologiques mentionnés ci-dessus, le manque actuel d’études sur ce sujet complique également la généralisation des effets de l’IPC sur la performance sportive. À ce stade, une compréhension des mécanismes physiologiques de ce phénomène devient essentielle pour comprendre ses possibilités d’application dans le sport et ses impacts physiologiques. La section suivante détaille les mécanismes potentiels sous-jacents par lesquels l’IPC modifie les réponses physiologiques et augmente la performance physique.
10
1.4 Mécanismes d’actions de l’IPC
Les mécanismes d’action sous-jacents de l’IPC sont complexes et intègrent une multitude de processus qui comportent eux-mêmes encore plusieurs éléments controversés ou inconnus. La complexité s’accentue davantage lorsqu’il est question d’IPC indirect à distance. En effet, le(s) mécanisme(s) par lequel(s) le préconditionnement offre une protection systémique (c.-à-d., à plusieurs organes du corps) est inconnu. On débat autour de trois hypothèses principales pour expliquer la propagation du signal de l’organe préconditionné à l’organe cible qui est, dans la plupart des études, le myocarde. Les trois voies mécanistiques postulées sont la voie humorale, la voie neuronale et l’activation d’une réponse systémique protectrice. Néanmoins, la réponse serait vraisemblablement la combinaison de ces dernières puisque ces voies peuvent très bien interagir sans que l’une d’elles exclue les autres (Hausenloy & Yellon, 2008; Heusch, Botker, Przyklenk, Redington, & Yellon, 2015).
Brièvement, la voie humorale implique qu’une substance endogène soit relâchée dans la circulation sanguine pour lui permettre d’atteindre l’organe cible. La nature exacte de(s) la molécule(s) responsable(s) est elle aussi inconnue, mais les autocoïdes, hormones libérées par la cellule, telles que l’adénosine (Pell, Baxter, Yellon, & Drew, 1998), la bradykinine (Schoemaker & van Heijningen, 2000), et les opioïdes (Patel, Moore, Hsu, & Gross, 2002) ont été étudiées. Certaines études se sont aussi intéressées au facteur de transcription inductible par hypoxie (HIF)-1α (Cai, Luo, Zhan, & Semenza, 2013; Kalakech et al., 2013). L’hypothèse hormonale est supportée par les études montrant un transfert de la protection d’un animal préconditionné à un autre n’ayant jamais subi la procédure, via une transfusion sanguine (Dickson et al., 2001; Dickson, Reinhardt, et al., 1999), une transplantation d’organe préconditionné (Konstantinov et al., 2005), ou d’effluent coronaire (Dickson, Lorbar, et al., 1999). Le fait que la reperfusion sanguine soit nécessaire pour que l’IPC entraîne des effets suggère que le facteur protecteur doit être mis en circulation pour atteindre l’organe cible et vient aussi supporter l’hypothèse de la voie hormonale (Gho, Schoemaker, van den Doel, Duncker, & Verdouw, 1996; Weinbrenner, Nelles, Herzog, Sarvary, & Strasser, 2002).
L’activation de la voie neuronale pourrait être modulée par les mêmes autocoïdes que ceux de la voie hormonale, mis à part les opioïdes. En effet, l’adénosine (Dong, Liu, Ji, & He, 2004; Liem, Verdouw, Ploeg, Kazim, & Duncker, 2002) et la bradykinine (Schoemaker & van Heijningen, 2000) agiraient comme médiateurs endogènes pour stimuler les nerfs afférents locaux qui activent à leur tour des voies efférentes jusqu’aux différents organes. L’oxyde nitrique (NO) fait également partie des molécules soupçonnées dans le déclenchement de l’IPC (Bolli et al., 1997). Les évidences impliquant le système nerveux autonome proviennent d’études démontrant que l’utilisation de bloqueurs de ganglions, comme l’hexamethonium (Liem et al., 2002; Schoemaker & van Heijningen, 2000) pour la bradykinine, inhibe la réponse protectrice
11 normalement observée après l’IPC. Corroborant ces observations, certaines recherches ont rapporté que le sectionnement de nerfs (Dong et al., 2004) abolissait la protection fournie par l’IPC. Chose intéressante, la résection d’un seul nerf de la jambe, soit le nerf sciatique ou fémoral, bloque que partiellement les effets de l’IPC (Lim, Yellon, & Hausenloy, 2010).
Finalement, la réponse systémique impliquerait la modulation du système immunitaire. Très peu d’études se sont penchées sur ce phénomène et les données à ce propos sont controversées (Berger, Macholz, et al., 2015). La modulation du système immunitaire ne sera donc pas décrite en détail dans ce mémoire. De façon générale, la réponse inflammatoire serait supprimée et la transcription de gènes anti-inflammatoires favorisée (Hausenloy & Yellon, 2008). Kharbanda et coll. ont observé une diminution dans l’expression des neutrophiles chez l’humain (Kharbanda et al., 2001) et Albrecht et coll. une élévation des cytokines lors de la première phase de protection (Albrecht et al., 2013).
Peu importe la ou les voies empruntées, les brefs épisodes d’ischémie/reperfusion entraîneraient la libération de signaux chimiques (déclencheurs) qui activeraient des kinases et plusieurs facteurs de transcription. Ainsi, plutôt que de défendre l’une ou l’autre de ces voies, nous allons aborder les composantes de cette cascade métabolique pour mieux comprendre les impacts physiologiques transitoires de l’IPC; la phase tardive, présentée dans la revue de littérature de Bolli (Bolli, 2000), ne sera pas détaillée ici. Rapidement, la phase tardive est une réponse universelle au stress, qui de par ses effets prolongés, nécessite la fabrication de nouvelles protéines. À l’inverse, la phase classique est quasi immédiate et utilise des médiateurs qui effectuent des modifications de protéines déjà présentes. Néanmoins, ces deux phases ont plusieurs mécanismes en communs puisque c’est cette « fameuse » cascade métabolique qui va déclencher la transcription de gènes et la synthèse de protéines responsables de la phase de protection tardive. Ainsi, peu importe la phase ou l’emplacement visé, la libération initiale de ces signaux chimiques après l’ischémie est essentielle pour déclencher la protection. Les deux prochaines sections s’intéressent aux déclencheurs, aux médiateurs et aux effecteurs finaux de l’IPC. Un schéma résumé (figure 3) sera également présenté à la fin de cette section.
1.4.1 Déclencheurs de l’IPC
Plusieurs substances ont été étudiées pour leur rôle potentiel comme déclencheurs de l’IPC et le consensus actuel semble favoriser l’interaction de plusieurs substances (figure 3). L’importance relative de chaque déclencheur n’est pas encore déterminée et les mécanismes impliqués pourraient varier d’une espèce à l’autre et selon l’organe visé (Hausenloy & Yellon, 2008). Ceci étant dit, la plupart des études se sont intéressées aux effets sur le myocarde et il semblerait que les mécanismes associés à la protection cardiaque
12
soient les mêmes que pour le muscle squelettique (Yellon & Downey, 2003). Voici les déclencheurs les plus souvent discutés dans la littérature scientifique.
1.4.1.1 Adénosine
Les épisodes d’ischémie induits par l’IPC génèrent rapidement de l’adénosine par la dégradation de l’ATP. Celle-ci va s’accumuler dans le tissu privé d’O2 (Cohen, Baines, & Downey, 2000). Cette hormone vasodilatatrice est considérée comme étant l’un des déclencheurs les plus importants et essentiels de l’IPC. L’adénosine agirait par les récepteurs A1 (Liu et al., 1991) et A3 (Liu et al., 1994); l’infusion d’un agoniste de chacun de ces récepteurs imite les effets de l’IPC. Toutefois, l’antagoniste de A1 ou A3, seul, ne bloque que partiellement les effets de l’IPC, suggérant l’implication simultanée des deux récepteurs (Liu et al., 1994). Des études utilisant un antagoniste non sélectif de l’adénosine ont montré une abolition complète de la protection, et ont démontré l’importance de la voie neuronale par sectionnement de nerfs (Dong et al., 2004; Liem et al., 2002). Dans le muscle squelettique, le RIPC augmente les niveaux d’adénosine plasmatique (Bushell et al., 2002). En plus des effets vasodilatateurs, l’activation des récepteurs de l’adénosine permettrait également d’améliorer la fonction mitochondriale en diminuant la production des dérivés réactifs de l’O2 (ROS), des composés qui peuvent endommager les structures cellulaires et créer un stress oxydatif (voir section 1.4.1.4), et en améliorant la respiration cellulaire, soit l’efficience de la mitochondrie (Leung et al., 2014).
1.4.1.2 Opioïdes
L’injection d’un antagoniste non spécifique des récepteurs opioïdes, le naloxone, abolit la protection sur le myocarde des rats induit par un seul cycle d’ischémie/reperfusion (Patel et al., 2002), mais ne permet pas de bloquer la protection induite par plusieurs cycles (Miki, Cohen, & Downey, 1998). Contrairement à l’adénosine, les opioïdes agiraient par la voie hormonale seulement. Cette hypothèse est supportée par l’étude d’Addison et coll. qui a montré que le blocage des ganglions autonomes par un antagoniste (hexamethonium) des opioïdes n’abolit pas ses effets protecteurs sur le muscle squelettique (Addison et al., 2003). Les effets sur le cœur seraient bloqués par un antagoniste spécifique du récepteur κ1 (Zhang et al., 2006), et ceux sur le muscle squelettique par celui du récepteur δ1 (Addison et al., 2003). Le préconditionnement induit par les opioïdes avant une ischémie du muscle squelettique entraînerait une diminution de la déplétion d’ATP, de l’accumulation de lactate et de l’infiltration des neutrophiles dans le muscle (Addison et al., 2003). L’activation des récepteurs δ1 mimait aussi les effets protecteurs induits par le RIPC (Weinbrenner et al., 2002). La contribution individuelle de chaque sous-type des récepteurs requiert encore des investigations (Hausenloy & Yellon, 2008).
13
1.4.1.3 Bradykinine
La bradykinine est une autre hormone vasodilatatrice libérée lors de l’ischémie. Goto et son équipe (Goto et al., 1995) ont démontré que l’injection d’un antagoniste du récepteur B2 bloque complètement la protection offerte par un cycle d’ischémie/reperfusion et que l’infusion de bradykinine diminue l’infarctus au cœur du lapin. Avec la répétition des cycles, il n’est plus possible de bloquer la protection avec un antagoniste de la bradykinine seulement (Cohen et al., 2000). McNeil et coll. (McNeil, Allen, Olympico, Shoemaker, & Rice, 2015) ont aussi montré des augmentations des niveaux de bradykinines et leurs rôles dans la stimulation des nerfs sensoriels afférents chez les rats. L’adénosine, les opioïdes et la bradykinine seraient tous relâchés lors de l’ischémie et participeraient de façon conjointe à l’atteinte du seuil métabolique en se fixant aux protéines G de la membrane cellulaire qui activent les kinases intracellulaires, notamment la protéine kinase C (PKC) qui sera abordée dans la prochaine section.
1.4.1.4 Dérivés réactifs de l’O2 (ROS)
Les ROS pourraient agir à la fois comme déclencheur et médiateur de la voie de signalisation de l’IPC (Berger, Macholz, et al., 2015). Leur rôle de déclencheur est supporté par des études qui démontrent que l’injection de piégeurs des radicaux abolit la protection fournie par l’IPC (Baines, Goto, & Downey, 1997; Weinbrenner et al., 2002) et que l’infusion de substances permettant de former un système générateur de ROS peut mimer les effets du préconditionnement (Baines et al., 1997). Ce déclencheur n’aurait pas besoin de se fixer aux récepteurs, mais participerait de façon parallèle aux autres hormones pour atteindre le seuil métabolique nécessaire au préconditionnement (Cohen et al., 2000). Encore une fois, la protection ne peut pas être bloquée uniquement par l’inhibition des ROS lorsque plusieurs cycles sont effectués (Baines et al., 1997; Weinbrenner et al., 2002). Aspect intéressant, les ROS activeraient directement les kinases intracellulaires pour la transcription de protéines protectrices et convergeraient aussi vers la PKC (Baines et al., 1997; von Ruecker, Han-Jeon, Wild, & Bidlingmaier, 1989). En effet, la protection fournit par l’adénosine (Sakamoto, Miura, Goto, & Iimura, 1995), la bradykinine (Goto et al., 1995), les opioïdes (Miki et al., 1998) ou les radicaux libres (Baines et al., 1997) peuvent être abolis lors de l’inhibition de PKC. Les radicaux libres pourraient atteindre l’organe cible à la fois par la circulation sanguine et la voie neuronale (Chen, Tu, Connolly, & Ronnett, 2005).
1.4.1.5 Oxyde nitrique (NO)
Les épisodes d’ischémie/reperfusion causent la génération rapide de NO qui est un puissant vasodilatateur. La conversion de L-arginine en NO requiert de l’O2 et n’est possible que par trois synthases différentes (neuronale, inductible, endothéliale) (Daff, 2010). Dans la phase de préconditionnement classique, la formation de NO serait plus attribuable à l’isoforme neuronale , exprimée dans les cellules nerveuses,
14
cardiaques et musculaires, ainsi qu’à l’isoforme endothéliale (eNOS) exprimée dans les cellules des vaisseaux sanguins (Daff, 2010). La génération de NO par eNOS est dépendante des forces de cisaillement induites, de façon aiguë ou chronique (Miller & Vanhoutte, 1988), par les changements de flux sanguin qui activent la guanylylcyclase et augmente la formation de guanosine monophosphate cyclique (Lochner et al., 2002). La modulation de la formation de guanosine monophosphate cyclique par NO pourrait être bénéfique en diminuant la déplétion d’ATP (Ebihara & Karmazyn, 1996). De plus, NO pourrait moduler l’activité des canaux ioniques dans la mitochondrie (Lochner et al., 2002). En plus d’être un puissant vasodilatateur, NO atténuerait la consommation d’O2 par la mitochondrie, autrement dit l’efficience de celle-ci, ce qui pourrait influencer la capacité à consommer de l’O2 durant l’exercice (Cooper & Brown, 2008; Prime et al., 2009). Ces modifications au niveau de la respiration mitochondriale seraient encore plus évidentes lorsque la concentration en O2 est réduite (c.-à-d. hypoxie) (Shiva et al., 2007). La stimulation d'eNOS via les manipulations du flux sanguin pourrait expliquer l’augmentation du NO durant la manœuvre d’IPC (Bolli et al., 1997; Heusch et al., 2015). D’ailleurs, les effets protecteurs de l’IPC peuvent être abolis avec l’administration d’inhibiteurs de NO (Bolli et al., 1997; Lochner et al., 2002) ou de la guanylylcyclase (Lochner et al., 2002) et par l’ablation génétique d'eNOS (Przyklenk, Darling, Dickson, & Whittaker, 2003). L’administration de NO peut aussi mimer les effets de l’IPC (Lochner et al., 2002). La formation de NO stimulerait les nerfs sensoriels sensibles à la capsaïcine ce qui entraînerait la libération de peptide lié au gène de la calcitonine (CGRP) dans la circulation et activerait PKC (Xiao, Lu, Hu, Deng, & Li, 2001). Le NO pourrait aussi diminuer le seuil métabolique nécessaire pour observer des effets positifs (Bell & Yellon, 2001).
1.4.1.6 Facteur de transcription inductible par hypoxie (HIF)-1α
Ce facteur de transcription est induit en réponse aux changements de disponibilité d’O2 dans l’environnement et serait en partie responsable des adaptations observées en conditions d’ischémie (Randhawa, Bali, & Jaggi, 2015). Le rôle de cette molécule dans la cascade de l’IPC est encore controversé (Heusch et al., 2015). L’étude de Cai et son équipe (Cai et al., 2013) démontre que la protection cardiaque chez la souris est dépendante de l’expression du gène HIF-1α, et celle de Davidson (Davidson et al., 2013) que les niveaux circulants de HIF-1α sont augmentés et la protection au cœur partiellement annulée avec l’injection de l’inhibiteur. Albrecht et collègues (Albrecht et al., 2013) ont aussi observé une augmentation de l’expression des protéines HIF-1α dans le cœur de sujets ayant été soumis à l’IPC aux jambes, mais son implication directe demeure incertaine. Tout comme les substances précédentes, le facteur HIF-1α ne serait pas le seul à être impliqué dans cette réaction complexe (Heusch et al., 2015).
15 En résumé, l’adénosine, les opioïdes et la bradykinine se fixeraient aux protéines G, les ROS agiraient à la fois comme déclencheur et médiateur, et le NO activerait la synthèse de peptide lié au gène de la calcitonine. Dans tous les cas, ils déclencheraient l’IPC en activant la protéine kinase C. Le facteur de transcription inductible par hypoxie (HIF)-1α pourrait aussi agir en tant que déclencheur, mais la séquence est toujours méconnue. La prochaine section abordera les principaux médiateurs et effecteurs finaux connus de ces voies de signalisation.
1.4.2 Médiateurs et effecteurs finaux de l’IPC
Les médiateurs impliqués dans les voies de signalisation de l’IPC sont très peu connus et les incertitudes entourant les effecteurs finaux sont encore plus grandes. Étant donné la multitude de voies possibles dans l’IPC, seule la séquence mécanistique la plus acceptée et ses composés prédominants seront abordés dans ce mémoire (figure 3).
Les CGRP sont des neurotransmetteurs libérés grâce à la génération de NO qui stimuleraient les nerfs sensoriels sensibles à la capsaïcine situés dans les intestins. Les CGRP agiraient alors comme médiateur hormonal en étant transportés dans la circulation sanguine jusqu’à l’organe cible où ils activeraient PKC (Li, Xiao, Peng, & Deng, 1996; Wolfrum et al., 2005; Xiao et al., 2001). En effet, l’inhibiteur de PKC n’a aucune influence sur la libération de CGRP, mais abolit la protection induite par son activation (Wolfrum et al., 2005).
Cet élément vient s’ajouter aux résultats de précédentes études qui ont démontré le rôle clé de PKC comme médiateur. Rappelons que les ROS (Baines et al., 1997), ainsi que la fixation de l’adénosine (Sakamoto et al., 1995), la bradykinine (Goto et al., 1995), et les opioïdes (Miki et al., 1998) aux protéines G, impliquent tous l’activation de PKC dans leur voie de signalisation. C’est pourquoi, malgré les incertitudes associées aux différents déclencheurs actifs, l’implication de PKC comme médiateur clé dans la signalisation de l’IPC est considérée comme incontournable (Hausenloy & Yellon, 2008; Shimizu et al., 2009). Les premières études (Mitchell et al., 1995; Ytrehus, Liu, & Downey, 1994) sur le rôle central de PKC ont démontré que ses inhibiteurs bloquaient la protection et que l’infusion de deux activateurs de cette protéine kinase entraînait les mêmes effets que l’IPC.
L’étape dépendante et suivant l’activation de PKC est l’ouverture des canaux potassium sensible à l’ATP (canaux KATP) de la mitochondrie. Gross et ses collègues (Gross & Auchampach, 1992) ont été les premiers à démontrer que l’ouverture des canaux KATP mimait les effets de l’IPC et que le blocage de ces derniers avait l’effet inverse. Ces résultats ont ensuite étaient confirmés dans plusieurs études (D'Alonzo, Darbenzio, Parham, & Grover, 1992; Schulz, Rose, & Heusch, 1994; Van Winkle et al., 1994). L’intervention de PKC, et possiblement d’autres kinases, pour ouvrir les canaux KATP, ainsi que son rôle clé dans l’IPC est maintenant
16
bien accepté (Yellon & Downey, 2003). De plus, Kirsch et collègues (Kirsch, Codina, Birnbaumer, & Brown, 1990) ont rapporté que la stimulation des récepteurs A1 par l’adénosine agit directement sur les canaux KATP et provoque leur ouverture. L’ouverture de ces canaux dépolarise la membrane mitochondriale et diminue la consommation de Ca2+ (Murphy & Steenbergen, 2008). Les canaux KATP ont aussi été associés à la sympatholyse fonctionnelle, une réponse intégrative importante durant l’exercice, qui entraîne une distribution du flux sanguin de manière à répondre aux besoins métaboliques (Joyner & Thomas, 2003; Keller, Ogoh, Greene, Olivencia-Yurvati, & Raven, 2004). Cette ouverture entraînerait également la génération de ROS qui activeraient des kinases et modulerait l’effecteur final permettant le maintien de l’intégrité de la mitochondrie (Pain et al., 2000).
Ceci explique en partie le fait que les ROS pourraient agir à la fois comme déclencheur en activant PKC, et comme médiateur en étant libérés suite à l’ouverture des canaux KATP (Hausenloy & Yellon, 2008). En effet, l’activation de PKC, suivi de l’ouverture des canaux KATP de la mitochondrie entraînerait une production supplémentaire de ROS qui pourrait alors activer des kinases prosurvie (Yellon & Downey, 2003) ou inhiber l’ouverture des pores de perméabilité transitionnelle de la mitochondrie (mPTP) (Costa et al., 2006). Des études utilisant l’IPC à distance ont montré que l’inhibition de l’ouverture des pores de la mitochondrie était abolie avec l’utilisation d’antagonistes des canaux KATP (c.-à-d. glibenclamide et 5-hydroxydecanoate) (Konstantinov et al., 2005; Pell et al., 1998). Les mPTP sont des canaux non spécifiques dans la membrane interne de la mitochondrie qui s’ouvrent lors de la reperfusion et peuvent causer la mort cellulaire en entraînant la déplétion d’ATP et en induisant le gonflement de la mitochondrie (Hausenloy & Yellon, 2003). La prévention de l’ouverture de ces pores pourrait être la dernière étape et pourrait être à l’origine de la protection induite par l’IPC, mais d’autres investigations sont nécessaires pour confirmer ce mécanisme (Hausenloy & Yellon, 2008).
En somme, l’IPC libère des molécules, pour la plupart ayant des effets vasodilatateurs, qui vont activer des voies de signalisation permettant de préserver et d’améliorer la fonction mitochondriale lors d’une ischémie subséquente. Ainsi, cette technique pourrait influencer à la fois la fonction endothéliale, oxydative et musculaire squelettique. Les trois prochaines sections s’intéresseront aux effets de l’IPC sur ces trois fonctions qui peuvent influencer les performances.
17
Figure 1-2 Schéma résumé des mécanismes de l'IPC (Source personnelle)
1.5 IPC et fonction musculaire
En 1990, Murry et ses collègues (Murry, Richard, Reimer, & Jennings, 1990) ont montré que l’utilisation en ATP et la production de lactate étaient similaires dans les 5 premières minutes d’ischémie, puis ralenties dans les cœurs de chien préconditionnés après 5 à 20 minutes d’ischémie. La glycolyse anaérobie et la dégradation du glycogène étaient également plus lentes étant donné l’accumulation réduite de glucose-1-phosphate, de glucose-6-phosphate et de lactate. La glycogénolyse était probablement ralentie en raison d’une diminution de l’activité de l’enzyme phosphorylase (Murry et al., 1990). Des résultats similaires ont été observés sur d’autres animaux. Selon Murry et ses collègues (Murry et al., 1990), le ralentissement de la déplétion totale d’ATP en condition d’ischémie serait davantage associé à une préservation des niveaux d’énergie, par une diminution de l’utilisation d’ATP, plutôt qu’à une augmentation de la production d’ATP. L’utilisation moindre de l’ATP pourrait être causée par une réduction de la demande énergétique durant l’ischémie, ou l’inhibition de certaines ATPases (Murry et al., 1990). Cette épargne d’énergie pourrait aussi passer par un meilleur couplage excitation/contraction, et une diminution du pompage ionique (Pang et al., 1995). D’ailleurs, le NO modulerait à la baisse l’activité des canaux ioniques dépendants du voltage dans la mitochondrie, ce qui diminuerait l’hydrolyse et préserverait l’ATP (Lochner et al., 2002). Le ralentissement de la glycolyse anaérobie souvent observé dans la littérature (Murry et al., 1990; Pang et al., 1995) serait aussi impliqué dans l’amélioration de la fonction contractile en diminuant l’accumulation de produits métaboliques. Fleet et ses
18
collègues (Fleet, Johnson, Graebner, & Gettes, 1985) ont observé une diminution progressive de l’acidose du myocarde avec la répétition d’épisodes ischémiques et Bailey et coll. (Bailey, Jones, et al., 2012), une atténuation de l’accumulation de lactate dans le sang lors d’un exercice progressif maximal précédé de quatre cycles de compression (220 mmHg) de 5 minutes aux deux jambes, et ce malgré une intensité identique entre les deux tests. La performance au contre-la-montre de 5 km suivant ce test était améliorée, mais aucune mesure de lactate n’était effectuée à ce moment.
L’amélioration de l’efficience énergétique s’observe également à la suite d’une période d’ischémie. On rapporte en effet qu’après trois cycles d’ischémie de 10 minutes, entrecoupés de 10 minutes de reperfusion, la force et la résistance à la fatigue du muscle squelettique chez le rat sont augmentées, en dépit de niveaux d’ATP et de phosphocréatine (PCr) inchangés (Gurke, Kuhrmeier, et al., 1996; Gurke, Marx, Sutter, Frentzel, Harder, et al., 1996). L’IPC favoriserait donc l’épargne de l’ATP pendant l’exercice, mais pourrait également accélérer sa resynthèse. En effet, Reimer et son équipe (Reimer, 1996) sont les premiers à rapporter une resynthèse plus rapide des phosphagènes durant la phase de reperfusion de l’IPC. Cette accélération de la resynthèse de l’ATP et de la PCr après une ischémie précédée de l’IPC est rapportée par d’autres études (Moses et al., 2005; Takaoka et al., 1999). Toujours à propos de la récupération, si les observations d’acidose réduite et de resynthèse accélérée des phosphagènes mettent en valeur les effets de l’IPC sur le métabolisme énergétique, elles soulignent également l’influence potentielle de la perfusion sanguine. Une amélioration de la perfusion pourrait être à l’origine des changements métaboliques observés dans le muscle squelettique, et un consensus semble apparaitre dans la littérature quant aux effets de l’IPC sur la fonction endothéliale.
1.6 IPC et fonction endothéliale
Tel que mentionné à la section 1.4, les épisodes d’ischémie et de reperfusion entraînent la libération de plusieurs déclencheurs, dont notamment l’adénosine, la bradykinine et le NO qui sont des substances vasodilatatrices importantes. L’adénosine, connue pour son action sur le tonus vasculaire du muscle squelettique (U. Gustafsson, Gidlof, Lewis, & Sollevi, 1994), et le NO, puissant vasodilatateur (Lochner et al., 2002), régule la fonction endothéliale et influence ainsi directement la microcirculation et le débit sanguin (Lochner et al., 2002; Tapuria et al., 2008). Dans les dernières années, un bon nombre d’études se sont intéressées aux modifications de la fonction endothéliale chez l’humain engendrées par l’IPC, dont l’un des paramètres les plus évoqués est la vasodilatation dépendante du flux sanguin (FMD). Ceci nous permet donc de nous concentrer presque uniquement sur les études effectuées sur les humains in vivo.
19 En 2001, Kharbanda et ses collègues (Kharbanda et al., 2001) s’intéressent aux effets de l’IPC sur la circulation périphérique en soumettant 31 volontaires à une ischémie brachiale de 20 minutes, suivant ou non, un protocole de préconditionnement de trois cycles de compression du bras de 5 minutes à 200 mmHg. Ils démontrent que les détériorations de la FMD à l’artère brachiale et de la réponse vasodilatatrice à l’acétylcholine, normalement observées après une ischémie prolongée, sont prévenues lorsque l’ischémie est précédée de l’IPC. Ils constatent également une inhibition de l’activation des neutrophiles en circulation, suggérant que l’IPC pourrait protéger à la fois les conduits artériels et la microcirculation de façon locale en prévenant le dysfonctionnement endothélial et la réponse inflammatoire (Kharbanda et al., 2001). Ces résultats d’une protection de la fonction endothéliale (c.-à-d., prévention de la détérioration de la FMD) induite par l’IPC ont été confirmés lors d’un protocole quasi identique avec pression de 220 mmHg (van den Munckhof et al., 2013). Bien qu’intéressantes, ces études ont essentiellement observé des bénéfices périphériques locaux, et ne permettent pas de savoir si la libération des déclencheurs induite par l’IPC était suffisante pour induire une vasodilatation systémique et une protection généralisée.
Une vasodilatation systémique (et non seulement sur l’organe affecté par l’IPC) a été observée par Shimizu et ses collègues (Shimizu, Konstantinov, Kharbanda, Cheung, & Redington, 2007). Les auteurs rapportent qu’un protocole d’IPC sur les jambes de cochons entraîne une diminution de la résistance de vaisseaux coronaires et une augmentation du flux sanguin coronaire. En 2011, Kraemer et son équipe (Kraemer et al., 2011) démontrent, par des analyses effectuées avec un laser-doppler et spectrophotométrie, que l’IPC au bras améliore le flux sanguin capillaire (35%) de la jambe ainsi que sa saturation en O2 de 29%. Cette amélioration de la microcirculation cutanée, à distance de l’organe préconditionné, était immédiate. Finalement, on rapporte que trois cycles de compression à 200 mmHg au bras gauche génèrent une vasodilatation des vaisseaux du bras droit ainsi qu’une élévation de l’activité parasympathique cardiaque (Enko et al., 2011). Lorsqu’un protocole d’IPC est réalisé de façon chronique durant 4 semaines, on observe aussi une vasodilatation dépendante de l’endothélium par une augmentation de la production de NO, des niveaux de facteur de croissance vasculaire endothélial (VEGF) et des cellules progénitrices endothéliales (EPCs) (Kimura et al., 2007). En effet, en condition d’hypoxie, la production de HIF-1α active l’expression génique de VEGF (T. Gustafsson & Kraus, 2001) entraînant une augmentation du nombre d’EPCs circulantes qui sont associées à la fonction endothéliale chez l’humain (Hill et al., 2003). En utilisant un préconditionnement comprenant quatre cycles de 5 minutes de compression sur le bras non dominant de volontaires sains durant 7 jours consécutifs, Jones et collègues (Jones et al., 2014) constatent que l’IPC améliore la FMD et la microcirculation, évaluée par conductance vasculaire cutanée, et ce dans les deux bras. Aspect intéressant, ces adaptations vasculaires ont persisté jusqu’à 8 jours après la fin de l’intervention. Ces études chroniques corroborent parfaitement les résultats d’études utilisant l’IPC de façon transitoire, mais concernent davantage la phase tardive de l’IPC et la synthèse protéique qui s’ensuit.
20
Une fonction endothéliale plus efficace et une vasodilatation périphérique induite par un protocole IPC pourraient être déterminantes dans les sports qui requièrent de maintenir une intensité élevée pendant un certain temps ou de façon répétée. Néanmoins, l’amélioration de la perfusion ne s’accompagne pas automatiquement d’une meilleure oxygénation des tissus musculaires. La prochaine section s’intéresse aux effets observés de l’IPC sur la fonction oxydative du muscle squelettique.
1.7 IPC et fonction oxydative
Compte tenu des perturbations physiologiques mentionnées aux sections précédentes, l’IPC pourrait influencer la fonction oxydative de plusieurs façons. Il existe très peu d’études ayant examiné de façon robuste l’effet ergogénique de l’IPC sur la consommation d’O2 musculaire. Néanmoins, plusieurs arguments scientifiques permettent de penser qu’un protocole d’IPC efficace pourrait engendrer une augmentation de l’extraction musculaire d’O2. Tout d’abord, en entraînant une vasodilatation locale et systémique ainsi qu’une amélioration de la fonction endothéliale, on peut supposer d’une amélioration de l’oxygénation durant l’effort. En 2004, Saito et collègues (Saito, Komiyama, Aramoto, Miyata, & Shigematsu, 2004) ont observé une augmentation de l’oxygénation du muscle chez les rats durant l’exercice. Puis en 2012, l’équipe de Kanoria (Kanoria et al., 2012) a rapporté que l’IPC aux jambes améliorait la fonction mitochondriale et réduisait l’acidose au foie chez les lapins. Chez l’humain, l’IPC prolonge le temps en apnée (Kjeld et al., 2014), et selon une étude de Barbosa et coll. (Barbosa et al., 2015), l’IPC exécuté avant un protocole de contractions sous-maximales répétées, augmente la désoxygénation et retarde l’apparition de la fatigue. Cette désoxygénation pourrait aussi être associée à la prolongation du temps d’effort et demande donc des validations supplémentaires. À notre connaissance, ces études sont les seules à avoir étudié la fonction oxydative après l’IPC, et parmi celles faites chez l’humain, aucune ne s’est intéressée directement aux modifications dans l’extraction d’O2. Parmi les méthodes utilisées pour investiguer sur le métabolisme oxydatif du muscle squelettique, l’une d’elles est invasive, la biopsie, et les deux autres sont non-invasives; la spectroscopie par résonance magnétique et la spectroscopie proche infrarouge (NIRS). La spectroscopie par résonance magnétique est considérée comme la méthode étalon pour évaluer la capacité mitochondriale en mesurant les cinétiques de resynthèse de la PCr et le pH intramusculaire. En effet, la resynthèse de PCr est directement liée à la présence et la quantité d’O2, ainsi qu’au potentiel du muscle donc à l’oxygénation des tissus musculaires (Glaister, 2005). L’IPC précédant une ischémie prolongée entraîne une amélioration dans la resynthèse de l’ATP et de PCr chez l’animal (Dealbuquerque, Gerstenblith, & Weiss, 1994; Takaoka et al., 1999), et chez l’humain (Andreas et al., 2011). Andreas et collègues ont été les premiers à montrer que l’induction de trois cycles de compression à 200 mmHg (5 minutes) entraînait une augmentation de la
21 consommation d’O2 et de la production de PCr durant la reperfusion (Andreas et al., 2011). La littérature actuelle sur l’IPC est encore peu élaborée à ce propos et ne permet pas de tirer de conclusion sur les bénéfices ou non de cette technique sur la réoxygénation, amélioration qui pourrait pourtant entraîner des effets significatifs sur la performance, notamment lors d’efforts maximaux répétés entrecoupés de périodes de récupération. La production de NO lors de l’IPC pourrait, sans augmenter l’extraction, atténuait la consommation d’O2 par la mitochondrie, autrement dit l’efficience de celle-ci (Cooper & Brown, 2008; Prime et al., 2009). De plus, ces modifications au niveau de la respiration mitochondriale seraient encore plus évidentes lorsque la concentration en O2 est réduite (c.-à-d. exercice maximal et hypoxie) (Shiva et al., 2007). Tous ces arguments physiologiques jouent en faveur de l’IPC comme moyen d’augmenter l’extraction d’O2 à l’exercice. Si le métabolisme aérobie et la fonction mitochondriale peuvent être étudiés au moyen de méthodes telles que la biopsie musculaire ou l’imagerie par résonance magnétique, ces techniques sont invasives, dispendieuses et peu pratiques dans le contexte de l’entraînement sportif. La technique du NIRS est non invasive et permet l’analyse continue de l’hémodynamique et de l’oxygénation du muscle squelettique (M. C. P. van Beekvelt, Colier, Wevers, & van Engelen, 2001). Cette technique se prête donc parfaitement à l’étude des changements physiologiques induits par l’IPC.


![Figure 1-4: Signaux de la [HbO 2 ] en rouge, de la [HHb] en bleu et de la [THb] en vert](https://thumb-eu.123doks.com/thumbv2/123doknet/6575646.177999/41.918.147.800.518.749/figure-signaux-hbo-rouge-hhb-bleu-thb-vert.webp)

![Figure 3-2. Change in resting muscle blood volume amplitude (∆[THb]) from baseline to post- SHAM and IPC conditions](https://thumb-eu.123doks.com/thumbv2/123doknet/6575646.177999/64.918.200.685.221.583/figure-change-resting-muscle-volume-amplitude-baseline-conditions.webp)