ETUDE DE L'ACTIVITE DES PROTEINES PRECOCES DU VIRUS DU POLYOME DANS LE PROCESSUS D'ONCOGENESE
IN VIVO ET IN VITRO
par
Louise Bouchard
Département de Microbiologie
Thèse présentée à la Faculté de Médecine en vue de l'obtention du grade de
Philosophiae Doctor CPh.D.>
TABLE DES MATIERES
TABLE DES MATIERES. • • • • • • • • • • • • • • • • • • • • • • • • • • • • . • • • • • • • ii
LISTE DES FIGURES... vi
LISTE DES TABLEAUX ••••••••••••••••••••••••••••••••••••• viii LISTE DES ABREVIATIONS... ix
RESUME • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • ., • • • x i INTRODUCTION • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • 1 MATERIEL ET METHODES... 15
I. Souche virale... 15
II. Plasmides recombinants... 15
III. Induction de tumeurs par injection de plasmides recombinants... 18
IV. Etablissement des lignées tumorales... 18
V. Analyse pathologique des tumeurs... 19
VI. Transfection de cellules en culture... 20
VII. Clonage en agar noble •••••••••• 22 VIII. Evaluation du potentiel tumorigène des cellules transformées... 23
IX. Mesure du taux d'apparition des variants dans les populations cellulaires... 23
1. Taux de conversion des cellules non-transformées au phénotype transformé... 24
2. Taux de conversion des cellules transformées au phénotype normal... 25
3. Taux de conversion des cellules sensibles au 6418 en cellules résistantes... 25
4. Evaluation de la valeur statistique du test
de fluctuation... 26
X. Mise en évidence des antigènes T dans les
lignées transformées... 27
1. Immunofluorescence... 27
2. Détection de l'activité kinasique associée
à l'antigène T moyen <in vitro)... 28
i. Immunoprécipitation des antigènes T,
marquage et électrophorèse... 28
ii. Traitement des gels pour
autoradiographie... 31
XI. Caractérisation du patron d'intégration de
l'ADN viral dans l'ADN des lignées cellulaires 32
1. Extraction de l'ADN cellulaire... 32
2. Transfert sur filtre de nitrocellulose... 33
3. Préhybridation et hybridation des filtres
à une sonde radioactive... 34
4. Préparation d'une sonde radioactive... 35
RESULTATS. • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • 38
Première partie: Caractérisation du système d'injection
d'ADN recombinant à des rats nouveaux-nés
I. Evaluation du potentiel tumorigène de l'ADN
viral cloné... 38
II. Propriétés des lignées tumorales... 44
III. Analyse des séquences virales intégrées dans
l'ADN des cellules tumorales... 47
IV. Effet de la coinjection d'un ADN
sur le potentiel tumorigène de l'ADN viral.... 50
V. Analyse pathologique des tumeurs... 53
Deuxième partie: Activité des protéines précoces de
polyome dans la transformation in vitro I. Etude des propriétés des cellules contenant le
phénotype transformé... 59 1. Morphologie des cellules sélectionnées pour
leur résistance au 6418... 62
2. Caractéristiques de croissance des lignées
résistantes au 6418... 66
3. Propriétés des lignées sélectionnées pour la
formation de foyers sur plastique... 68
4. Détection de l'antigène T moyen dans les
lignées cellulaires... 69
II. Etude de la dynamique des populations
cellulaires... 72
1. Conversion des cellules non transformées
au phénotype transformé... 73
2. Réversion des cellules transformées au
phénotype normal... 76
3. Influence de l'activité d'un gène sur
l'activité du gène adjacent... 84
i. Caractéristiques de croissance des
révertants et de leurs retransformants.. 85
ii. Comportement des révertants et de leurs
retransformants en présence de 6418.... 88
III. Effet du grand T de polyome dans les cellules normales contenant des copies silencieuses
du gène T moyen... 94
1. Description de l'essai utilisé... 94
2. Retransformation de la lignée 8-2 par le
gène
Tmoyen...
963. Effet du grand T de polyome dans la lignée E3--:2. • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • 4. Effet de certains grand T mutants dans la
lignée 8-2 •••••••••••••••••••••••••••••••• 103 i. Effet du mutant LT97... 103 ii. Effet du grand T portant la
mutation tsa... 103
6. Détection des protéines virales produites par les cellules 8-2 transformées sous
l'action de grand T ••••••••••••••••••••••• 106 i. Immunofluorescence... 106
ii. Essai d'activité kinasique associée à
T moyen... 107 7. Analyse des séquences virales intégrées
dans les cellules 8-2 transformées sous
l'action de grand T ••••••••••••••••••••••• 111 8. Stabilité de l'insert du gène T moyen dans
la lignée 8-2 et dans ses retransformants
spontanés... 114 9. Effet du grand T de polyome dans les
lignées portant le recombinant pMT97A8t... 120 10. Analyse des séquences virales intégrées
dans les lignées transformées sous
l'action de grand T ••••••••••••••••••••••• 129
DISCUSSION. • • • • • • • • • • • • • • • . • • • • • . • . • • • • • • • • • • • • • • • • • • • • 132
I. Caractérisation du système d'injection d'ADN
recombinant à des rats nouveaux-nés... 132
II. Activité des protéines précoces du virus du
polyome in vitro... 137
REMERCIEMENTS.... • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • 149
LISTE DES FIGURES
Figure 1. Carte physique du génome du virus du
pol yome.. . . . • . . . . . . . . 3
Figure 2. Carte physique des plasmides recombinants
pPAS et pPB21 •••••••••••••••••••••••••••••••• 41 Figure 3. Morphologie des lignées tumorales •••••••••••• 45
Figure 4. Croissance des lignées tumorales à forte
et à faible concentration de sérum ••••••••••• 46
Figure 5. Analyse par Southern blot des séquences
virales intégrées dans les lignées tumorales. 49
Figure 6. Effet de la présence d'un ADN sur
l'incidence des tumeurs •••••••••••••••••••••• 52 Figure 7. Coupes histologiques de tumeurs induites par
le recombinant pPAS en présence ou en absence
d'ADN ••••••••••••••••••••••••••••• 57
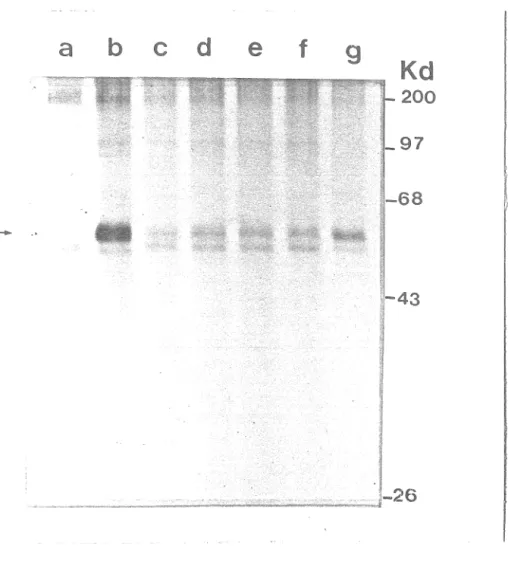
Figure 8. Activité kinasique associée à T moyen dans
les complexes immuns de cellules FR3T3
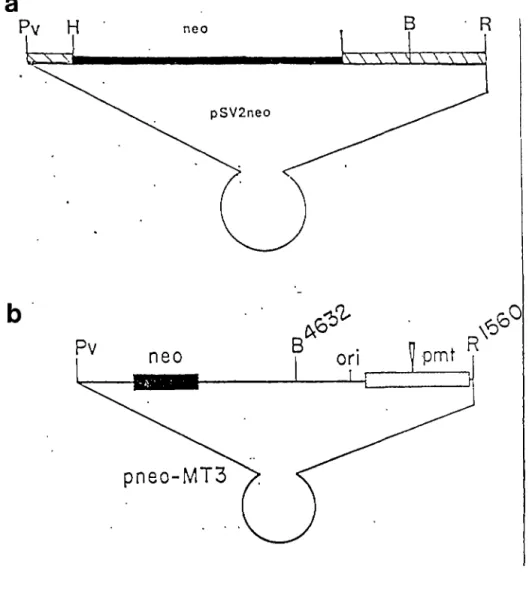
transformées par pPAS •••••••••••••••••••••••• 61 Figure 9. Structure des plasmides recombinants pSV2neo
et pneoMT3 •.••••••••••••••••••••••••••••••••• 63 Figure 10. Morphologie des colonies résistantes au 6418
après transfection de cellules FR3T3 avec
pneoMT3. • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • 65
Figure 11. Analyse des différents T moyen produits dans les lignées transformées par pdl8MT3 ••••••••• 77 Figure 12. Morphologie de quelques lignées dérivées de
la lignée FRneoMT-8 •••••••••••••••••••••••••• 86 Figure 13. Expression ·de l'antigène T moyen dans les
révertants et dans leurs retransformants ••••• 89 Figure 14. Détection par un essai d'activité kinasique
des formes de T moyen produites dans les cellules 8-2 retransformées avec le mutant
dl
a. . .
98Figure 15. Structure des plasmides recombinants
contenant le gène nec et différents grand T
de polyome ••••••••••••••••••••••••••••••••••• 100 Figure 16. Représentation graphique de l'effet des
différents grand T dans la lignée 8-2 •••••••• 102 Figure 17. Représentation graphique de l'effet du grand
T thermosensible dans la lignée 8-2 •••••••••• 105 Figure 18. Morphologie et immunofluorescence pour T
moyen des lignées 8-2 retransformées sous
l'action de grand T •••••••••••••••••••••••••• 108
Figure 19. Essai d'activité kinasique associée à T moyen
dans les lignées 8-2 transformées sous
l'action de grand T •••••••••••••••••••••••••• 109 Figure 20. Analyse des séquences virales intégrées dans
les lignées 8-2 transformées sous l'action
de grand T ••••••••••••••••••••••••••••••••••• 113
Figure 21. Détection des séquences spécifiques à grand T
dans les lignées 8-2 transformées sous son
action ••••••••••••••••••••••••••••••••••••••• 115 Figure 22. Analyse des séquences virales intégrées dans
18 sous-clones indépendants de la
lignée 8-2 ••••••••••••••••••••••••••••••••••• 117 Figure 23. Analyse des séquences virales intégrées dans
les transformants spontanés de la
lignée 8-2 ••••••••••••••••••••••••••••••••••• 119 Figure 24. Carte du plasmide recombinant pMT97A8t ••••••• 121 Figure 25. Activité de grand T dans la lignée
FRHyMT97ABt-2 ...•...••.•....•.•.•..•..• 125 Figure 26. Activité du grand T thermosensible dans la
lignée FRHyMT97A8t-3 ••••••••••••••••••••••••• 127
Figure 27. Activité kinasique associée à T moyen dans
les lignées FRHyMT97A8t transformées sous
l'action de grand T •••••••••••••••••••••••••• 128 Figure 28. Analyse des séquences virales intégrées
dans les lignées FRHyMT97A8t transformées
LISTE DES TABLEAUX
Tableau 1. Potentiel tumorigène des plasmides
recombinants ••••.••••••.•..••.•...•...••... 43 Tableau 2. Analyse pathologique des tumeurs induites
par pol yome. . . • . . . . . . . . • . . . . . . . . . . . . 55
Tableau 3. Propriétés des lignées sélectionnées pour
la résistance au 6418 •••••••••••••••••••••••• 67 Tableau 4. Phénotype des lignées contenant le plasmide
pneoMT3 •••••••••••••••••••••••••••••••••••••• 70 Tableau 5. Taux d'apparition des variants transformés
dans les lignées non transformées •••••••••••• 75 Tableau 6. Taux de réversion des cellules transformées •• 79 Tableau 7. Taux de réversion des lignées 3-lOneoMT •••••• 83 Tableau 8. Propriétés des révertants FRneoMT-8 et
de leurs retransformants ••••••••••••••••••••• 87 Tableau 9. Taux de conversion au phénotype transformé
des révertants de la lignée FRneoMT-8 •••••••• 90 Tableau 10. Taux de réacquisition de la résistance au
6418 des révertants sensibles •••••••••••••••• 92 Tableau 11. Activité des plasmides recombinants dans
la lignée 8-2 ••.••••.••••••••••.••••••••••••• 97 Tableau 12. Taux de conversion spontanée au phénotype
transformé des lignées FRHyMT97A8t ••••••••••• 123 Tableau 13. Activité du gène grand T dans les cellules
a. a. ATP cm cpm dATP dCTP dGTP DMSO EDTA Hep es Kb Kd M ml mm mM ng NP40 pb plv rpm SDS Tris TTP
LISTE DES ABREVIATIONS Acide aminé
Adénosine 5' triphosphate Degré celsius
Centimètre
Coups par minute
déoxy-adénosine 5' triphosphate
déoxy-cytidine 5' triphosphate
deoxy-guanosine 5' triphosphate Diméthylsulfoxide
Ethylène diamine tétraacétate Acide N-2-hydroxyethylpiperazine-N'-2-ethanesulfonique
Kilo paires de bases Kilodalton Molaire Millilitre Millimètre Millimolaire nanogramme Nonidet-P40 Paire de bases Poids/volume
Révolution par minute Sodium dodécyl sulfate Hydroxyméthyl-aminométhane Thymidine 5' triphosphate
uCi ug ul uM v/v Microcurie Microgramme Microlitre Micromolaire Volume/volume
RESUME
Nous avons étudié la capacité du virus du polyome
à induire le développement de tumeurs chez le rat. Pour
être tumorigène l'ADN viral doit être injecté à des animaux
nouveaux-nés sous forme linéaire. La coinjection d'un en-d'origine eucaryote n'a que peu d'effet sur
l'inci-dence des tumeurs. Par contre, des d'origine
procaryote manifestent un effet inhibiteur très marqué. De plus, la majorité des tumeurs ainsi obtenues sont bénignes alors que normalement, les tumeurs induites par polyome sont malignes.
Nous avons également étudié l'activité du gène T moyen dans un essai de transfection de lignées établies. Le gène
T moyen peut à lui seul induire le phénotype transformé.
Lorsque l'ADN transfecté contient le gène T moyen lié au
marqueur de sélection ngQ, la moitié des lignées résistantes
affichent un phénotype transformé. Ce dernier reflète l'ex-pression de T moyen. Les cellules présentant un phénotype normal et n'exprimant pas T moyen génèrent des variants
transformés à un taux de l'ordre de
to-e.
Cetteconver-sion s'accompagne de l'activation du gène T moyen et de _la production
leurs, les
d'une grande quantité de cet antigène.- Par ail-cellules transformées exprimant l'antigène T
moyen génèrent à un taux de l'ordre de des
réver-tants dans lesquels la protéine n'est plus produite.
re-produire l'effet de conversion au phénotype transformé lorsqu'il est introduit dans une lignée non transformée con-tenant plusieurs copies silencieuses du gène T moyen. La transformation est concomitante à l'activation du gène T moyen.
tants
Elle s'accompagne parfois de réarrangements imper-de l'insert imper-des gènes T moyen. Dans le but imper-de véri-fier si grand T agit en provoquant de la recombinaison homo-logue, nous avons produit des lignées contenant un tandem de deux gènes T moyen non transformants affectés par des lé-sions différentes. Grand T peut en effet provoquer la transformation de ces cellules, et celle-ci s'accompagne de l'apparition d'un T moyen fonctionnel. Ceci suggère que grand T agit directement ou indirectement en provoquant de la recombinaison homologue entre les copies inactives du gène T moyen. Ce processus pourrait rendre certaines copies du gène accessibles à la machinerie transcriptionnelle de la cellule, favorisant par le fait même l'expression de T moyen et la transformation.
INTRODUCTION
L'oncogenèse est un processus qui nécessite plusieurs étapes distinctes et pour lequel le virus du polyome cons-titue un excellent modèle d'étude. Ce virus a d'abord été identifié comme agent responsable du développement d'adéno-carcinomes des glandes salivaires chez la souris, son hOte naturel <Gross, 1959a,b). Toutefois, dans la majorité des cas, il provoque une infection silencieuse qui affecte peu l'animal. Par contre, il a la capacité d'induire une grande variété de tumeurs chez d'autres rongeurs comme le rat et le
hamster. C'est d'ailleurs cette polyvalence qui lui a valu
son nom <Stewart et Eddy, 1959).
Tout comme les virus SV40, BK et JC, le virus du polya-me est un polya-membre de la famille des papovavirus CTooze,
1981). Il s'agit d'un virus non-enveloppé composé d'une
capside protéique icosahédrique. Cette capside protège une molécule d'ADN bicaténaire circulaire de 5292 paires de
ba-ses, associée à des histones cellulaires pour former la
chromatine virale CSoeda et al., 1980). L'origine de répli-cation est située dans la partie non-codante du génome vi-ral. Cette même région comprend également les signaux
néces-saires à l'initiation de la transcription. De part et
d'au-tre de la région non-codante et sur les brins opposés s'étendent deux régions codantes, chacune occupant environ
la moitié du génome viral <Soeda et al, 1980) (figure la). La première, la région précoce, est exprimée au tout début du cycle lytique, avant la réplication du génome viral. La
seconde, la région tardive, est exprimée à la fin du cycle
lytique suite à la réplication du génome viral. Elle encode
trois protéines: VP1, VP2 et VP3 qui composent la capside virale <Fine et al., 1968: Roblin et al., 1971; Frearson et Crawford, 1972; Gibson, 1974; Hewick et al., 1975).
L'avènement de la culture des cellules a facilité l'étude des propriétés du virus du polyome. Les cellules en culture ont été classées en différentes catégories, suivant leur réaction lorsqu'elles sont mises en contact avec le vi-rus. Les cellules dites permissives sont celles qui permet-tent la réplication de l'ADN viral et qui produisent les
protéines de la capside. Ceci mène à l'assemblage d'une
grande quantité de particules virales et à leur libération
suite à la lyse cellulaire. Seules les cellules de souris
appartiennent à la catégorie des cellules permissives. Il
existe également des cellules qu'on appelle semi-permissi-ves, parmi lesquelles on compte les cellules de hamster et les cellules de rat. Chez ces espèces, seule une faible proportion de la population cellulaire permet la réplication du génome viral et la production d'une faible quantité de
virions. Les cellules non-permissives, comme par exemple
les cellules de singe ne permettent aucune réplication dé-tectable du génome viral.
divisé en 100 unités à partir de son unique site EcoRI, en
progressant dans le sens des aiguilles d'une montre. Les
nucléotides sont numérotés dans le même sens à partir de
l'origine de réplication. Les unités transcriptionnelles précoce et tardive sont indiquées de part et d'autre de l'origine, de même que les protéines qu'elles encodent
<Tooze, 1981).
B. Le génome viral est ici linéarisé en son site Hindi!! à
44.6 unités sur la carte génomique <nucléotide 3918> de
façon à mettre en évidence la région précoce. Les portions
de cette région encodant les antigènes tumoraux petit T <ST>, T moyen <MT>, et grand T <LT> sont également
illustrées. Les traits pointillés correspondent aux
introns, alors que les traits pleins représentent la portion non traduite des messagers. Le premier cadre de lecture qui code pour la région commune aux protéines précoces est en
noir. Le second cadre de lecture qui détermine la séquence
d'acides aminés unique à grand T est représenté en hachuré
alors que le troisième cadre de lecture qui encode la
séquence unique à T moyen est en blanc. Nous avons
également indiqué les portions de la région précoce qui sont affectées dans les différentes classes de mutants: hr-t, mlt, et ts.
8
50 1 60 1 70 1 80 1
9p
9
110 20 13p
4p
Hindlll Barn Hl Eco RI Hindlll Hincll Hincll Hindlll 1 11 1 1 1 11 1 1 1 11 Il 1 1 1 1 1 Il 1 1 1 1 1 1 1 1 ?1Ç11111 1 1 1 1 11 1 1 1 1 11 1 1 1 1 1
4000 5000/0 1000 2000 3000
ST An
Lorsqu'on infecte des cellules semi-permissives ou non-permissives avec le virus du polyome, ou encore qu'on y in-troduit l'ADN viral par transfection, la région précoce du génome viral est transcrite et la synthèse d'ADN cellulaire est stimulée. On observe alors l'expression transitoire des
propriétés caractéristiques des cellules transformées.
Après quelques divisions, la plupart des cellules cessent de
transcrire l'ADN viral, le perdent et retournent à l'état
normal. C'est le phénomène de la transformation abortive. Quelques cellules, cependant, stabilisent le génome viral en
l'intégrant à leur ADN. Elles peuvent ainsi poursuivre
l'expression des séquences virales. Ces cellules acquièrent alors de nouvelles propriétés parmi lesquelles on compte des changements morphologiques, une augmentation de la densité de saturation, l'indépendance d'ancrage, la perte d'inhibi-tion de contact, la diminud'inhibi-tion des besoins en facteurs séri-ques ainsi que le pouvoir d'induire le développement de tu-meurs chez des animaux syngéniques <Tooze, 1981>. La région
précoce encode à elle seule toutes les fonctions virales
né-cessaires à ce processus <Fried et Griffin, 1977; Israel et
al., 1979; Bastin et al., 1980; Hassel! et al., 1980; Novak et al., 1980).
La région précoce a une capacité codante colinéaire d'environ 100Kd. Par trois événements d'épissage d'un même messager, elle produit trois protéines différentes, les an-tigènes tumoraux petit T <195 a.a., 22Kd>, T moyen (421
a.a., 55Kd> et grand T (785 a.a., 90Kd>. Ces antigènes ont
d'abord été mis en évidence grâce à leur capacité à réagir
avec le sérum d'animaux ayant développé des tumeurs induites
par le virus du polyome. Ils partagent une partie
N-termi-nale commune de 79 acides aminés. Petit T et T moyen parta-gent, en plus, leurs 112 résidus suivants. Chacun a une portion C-terminale qui lui est propre puisqu'après l'événe-ment d'épissage, les trois messagers utilisent des cadres de
lecture différents (figure lb> <Habel, 1965; Ito al., 1977 a,b; Hunter et al., 1978; Hutchinson et al., 1978; Smart et Ito, 1978; Schaffhausen et al., 1978).
Les premières études visant à élucider le ou les roles
joués par chacune des protéines précoces dans le phénomène de la transformation par le virus du polyome ont été compli-quées par le fait qu'elles sont encodées par des séquences
chevauchantes. Il était alors très difficile d'étudier les
altérations cellulaires induites par une des protéines pré-coces en absence des deux autres. Toutefois, certaines évi-dences ont été obtenues de façon indirecte par l'étude des propriétés de divers mutants. Ces mutants ont été regroupés en différentes classes selon la portion du génome et les protéines précoces affectées. Ce sont les mutants ts, hr-t, et mlt (figure lb). La production de génomes viraux modi-fiés encodant séparément chacune des trois protéines préco-ces <Treisman et al.,1981a; Zhu et al., 1984> a permis une progression plus rapide de nos connaissances de leurs
pro-priétés et de leur rOle dans la transformation néoplasique. En ce qui concerne l'antigène petit T, ces connaissan-ces sont encore assez limitées. Aucune activité
biochimi-que, qu'elle soit intrinsèque ou due à une association avec
d'autres protéines, n'a encore été attribuée à cet antigène
que l'on retrouve aussi bien dans le cytoplasme que dans le
noyau des cellules (Zhu et al., 1984). On sait que le
pe-tit T est nécessaire au cycle lytique dans les cellules de
souris, mais son rOle exact n'est pas connu. Les seules
évidences dont on dispose actuellement suggèrent qu'il exer-ce un effet stimulant sur la réplication du génome viral
CMagnusson et al., 1981; Nilsson et Magnusson, 1983a;
Tem-pleton et al., 1986; Berger et Wintersberger, 1986). Dans
le processus de la transformation, le petit T augmente l'ag-glutinabilité des cellules par les lectines lorsque le grand
T est également présent (Liang et al., 1984>. Il réduit
l'attachement des cellules au plastique <Zhu et al., 1984; Cuzin, 1984) et augmente, faiblement mais de façon reproduc-tible, la densité de saturation des cellules <Cherington et al., 1986; Noda et al., 1986>. Il a été proposé que cet
ef-fet résulte d'une activité du petit T comparable à celle des
facteurs de croissance: il permettrait aux cellules d'outre-passer l'inhibition de croissance résultant d'une forte den-sité cellulaire <Noda et al., 1986). Finalement, il peut complémenter l'antigène T moyen dans le processus d'induc-tion de tumeurs chez les rats nouveaux-nés <Asselin et al.,
1983).
L'antigène T moyen , quant à lui, se retrouve associé
aux membranes cellulaires, du côté du cytoplasme. La majori-té des molécules semblent associées au réticulum endoplasmi-que <Ito et al., 1977b; Ito, 1979; Zhu et al., 1984; Dilworth et al., 1986). Cette phosphoprotéine se retrouve sous deux formes de 56 Kd et 58 Kd qui se distinguent par
leur abondance et leur taux de phosphorylation.
In
vivo,la phosphorylation se fait majoritairement sur les résidus
sérine et thréonine, alors qu'in vitro dans les
immuno-précipitats, il y a phosphorylation préférentielle des
ré-sidus tyrosine <Eckhart et al., 1979; Schaffhausen et Benja-min, 1979; Smith et al., 1979; Schaffhausen et BenjaBenja-min, 1981a,b; Ségawa et Ito, 1982). Cette activité kinasique sur
les résidus tyrosine n'est pas intrinsèque au T moyen, mais
résulte plutôt de son association avec la
pro-duit de l'oncogène cellulaire CCourtneidge et Smith,
1983, 1984; Cartwright et al., 1985; Schaffausen et al.,
1985). Conséquemment à cette association, on observe une
augmentation de l'activité kinasique de la
se reflète autant dans le degré de phosphorylation
qui du T moyen in vitro que dans l'apparition de nouveaux sites de phosphorylation en N-terminal de la pp60c--rc{Bolen et al., 1984; Balen et Israel, 1985; Yonemoto et al., 1985).
Il semble que le T moyen, plutôt que d'avoir un effet stimu-lant direct sur l'activité kinasique de la pp6oc-•rc,
em-pêcherait, de par son association avec cette dernière, la phosphorylation de sa tyrosine 527. Cette tyrosine consti-tuerait un site régulateur dont la non-phosphorylation géné-rerait une forme plus active de la
1985; Cooper et al., 1986; Cartwright et al., 1986, 1987; Piwnica-Worms et al.,1987>. Dans les cellules transformées par le virus du polyome il existe une forte corrélation
en-tre le niveau d'activité kinasique associée à T moyen et le
degré d'expression du phénotype transformé <Eckhart et al, 1979; Schaffhausen et al., 1979; Smith et al., 1979; Ito et al, 1980; Raptis et al., 1985). De plus, la majorité des mutants du virus qui sont défectifs pour la transformation ne présentent
<Patschinsky
pas d'activité kinasique associée à T
et al., 1982; Courtneidge et al., 1983;
moyen Bol en
et Israel, 1985>, alors qu'on ne aucun exemple de
mutant efficace pour la transformation et négatif dans un
essai d'activité kinasique. Si le complexe T
moyen-semble essentiel à la transformation cellulaire
<Amini et al., 1986>, il n'est toutefois pas suffisant, puisqu'il existe au moins un exemple de mutant non-transfor-mant de polyome qui forme un complexe actif avec la
et al., 1981; Nilsson et al., 1983>.
L'antigène T moyen peut se lier à d'autres protéines
cellu-laires, mais la signification de ces associations n'est pas connue <Schaffhausen, 1982; Walter et al., 1982; Ito et al., 1983; Grussenmeyer et al., 1985; Walter et al., 1987).
de phcsphcrylaticn du phcsphatidylincsitcl, un phospholipide membranaire. Il a été suggéré que le complexe T
soit à l'origine de cette activité. Le métabolisme de la ferme phcsphcrylée du phcsphatidylincsitcl génère du diacylglycércl et de l'incsitcl triphosphate qui constituent des signaux importants dans la régulation de la croissance cellulaire CNishizuka, 1984). Il serait donc possible que ces seconds messagers scient impliqués dans la transformation par le virus du pclycme (Whitman et al.,
1985; Kaplan et al., 1986; Koch et al., 1986>. Au cours du cycle lytique, dans les cellules de souris, T moyen semble nécessaire à l'assemblage des virions CGarcea et al., 1983; Turler et Salomon, 1985). Dans le processus de la transfor-mation, le gène T moyen constitue le principal oncogène du virus du pclycme CMes et Hassel!, 1982; Treisman et al., 1981b). En effet, seul, leT moyen est suffisant peur transformer les lignées cellulaires établies <Treisman et al., 1981b; Dcncghue et al., 1984; Gélinas et Bastin, 1985). Par centre, il ne transforme pas les cellules primaires et ne peut induire le développement de tumeurs chez les rats nouveaux-nés <Rassculzadegan et al., 1982; Asselin et al., 1983, 1984). Dans certains systèmes expérimentaux, toute-fois, il peut transformer les cellules embryonnaires de peu-let <Kaplan et al., 1985; Kcrnbluth ·et al., 1986) et induire des tumeurs chez les poulets et les hamsters nouveaux-nés
L'antigène grand T, finalement, est une phosphoprotéine
que l'on retrouve dans le noyau des cellules, associé à
l'euchromatine <Schaffhausen et al., 1978; Silver et al.,
1978; Dilworth et al., 1986; Hassauer et al., 1986). Il
forme des oligomères capables de se lier à l'ADN
bicaténai-re. Il manifeste une grande affinité pour l'origine de
ré-plication virale ainsi que pour les promoteurs des régions
précoce et tardive, alors qu'il a une faible affinité pour
les séquences non-spécifiques <Gaudray et al., 1980,1981; Pomerantz et al., 1983; Cowie et Kamen, 1984; Dilworth et
al. , 1984). Il possède également une activité ATPase
<Gaudray et al., 1980>. Au cours du cycle lytique dans les cellules de souris, il stimule la synthèse de l'ADN cellu-laire et initie la réplication du génome viral <Francke et Eckhart, 1973; Schlegel et Benjamin, 1978>. Il réprime la transcription de la région précoce et stimule celle de la région tardive <Cogen, 1978; Farmerie et Folk, 1984). Dans le processus de la transformation, on accorde au grand T le pouvoir d'immortaliser les cellules embryonnaires de rat <Rassoulzadegan et al., 1983; Asselin et Bastin, 1985). Les cellules ainsi immortalisées peuvent ensuite être transfor-mées par T moyen. Lorsqu'il est porté par un vecteur
rétro-viral, le grand T supprime la différenciation des
préadi-pocytes en adipréadi-pocytes (Cherington et al., 1986). Il réduit la dépendance des cellules envers les facteurs sériques <Rassoulzadegan et al., 1982> et est impliqué dans les phé-nomènes d'intégration et d'excision du génome viral de celui
de la cellule hôte <Basilico et al., 1979a,b; Della Valle et al., 1981; Colantuoni et al., 1982). C'est son rôle dans la transformation des lignées cellulaires établies qui fait
moins l'unanimité. La controverse à ce sujet vient en
gran-de partie gran-de résultats qui ont été obtenus avec le mutant ts-a. Ce mutant, comme tout ceu:< de sa catégorie, est affec-té dans la portion distale de la région précoce. Les
sé-quences impliquées sont uniques à l'antigène grand T. Il en
résulte la production d'un grand T thermosensible qui se
comporte comme la protéine de type sauvage à la température
permissive (33°C), mais qui est instable à 39oC,
tempé-rature dite restrictive. Les mutants ts transforment les
cellules à la température permissive alors que leur
poten-ti el transformant est réduit de façon draspoten-tique à la
tempé-rature restrictive. Lorsqu'on s'est intéressé au
comporte-ment des transformants obtenus à température permissive,
après transfert à température restrictive, certaines équipes
ont rapporté que ces transformants conservent leur phénotype <Fried, 1965a,b; Di Mayorca et al., 1969; Eckhart, 1969).
Ceci suggérait un rôle pour grand T dans l'établissement,
mais non dans le maintien de la transformation. D'un autre
cOté l'équipe de Cuzin a observé deux catégories de
trans-formants. Les transformants de type N révertent au
phénoty-pe normal à la température restrictive et encodent un grand
T complet. Les transformants de type A demeurent
transfor-més lorsqu'on les transfère à la température restrictive.
géné-ralement interrompues dans la portion distale de la région
précoce. Elles produisent donc une forme tronquée de grand
T d'où la mutation ts est absente. Ces résultats ont amené
Cuzin et son groupe à suggérer que le maintien de l'état
transformé nécessite la présence d'au moins une forme tron-quée de grand T <Seif et Cuzin, 1977; Rassoulzadegan et al.,
1981). Ceci entre en contradiction avec les résultats qui
ont montré que leT moyen seul est suffisant à
l'établisse-ment et au maintien de la transformation dans les lignées cellulaires établies <Treisman et al., 1981b; Donoghue et al., 1984; Gélinas et Bastin, 1985). Par ailleurs, des
ex-périences similaires ont amené un autre groupe à constater
qu'il n'y a aucun lien entre la perte ou le maintien du
phé-notype transformé à température restrictive et la présence
ou l'absence d'une forme tronquée de grand T. Il semble
donc que dans leur système, grand T ne soit pas nécessaire au maintien de l'état transformé <Fluck et Benjamin,
Fluck et al., 1983; Winberry et al., 1985).
1979;
Le présent travail se divise en deux parties. Dans un premier temps, nous avons caractérisé un système nous per-mettant d'évaluer le potentiel oncogène de différents gènes
viraux par injection directe à des rats nouveaux-nés. Nous
montrons que pour @tre tumorigène, l'ADN doit @tre injecté sous forme linéaire. La coinjection d'ADN cellulaire de
rat, comme ne semble pas avoir d'effet
coinjection d'ADN plasmidique a un effet inhibiteur marqué. De plus, les quelques tumeurs obtenues dans ce cas étaient le plus souvent bénignes, contrairement aux tumeurs obtenues
sans ou par coinjection d'un
eucaryo-te. En effet, dans ces dernières conditions, le génome
vi-ral de type sauvage induit invariablement des tumeurs
ma-lignes.
Dans la seconde partie de ce travail, nous avons utili-sé le système classique de transfection de cellules en cul-ture pour étudier les propriétés des cellules dans lesquel-les le gène T moyen est introduit sans qu'il y ait sélection pour le phénotype transformé. Nous avons lié le gène T moyen au gène néo, un marqueur de sélection dominant. Lorsqu'on sélectionne pour l'expression de néo, on remar-que une variabilité dans le phénotype des différentes gnées obtenues allant de presque normal pour certaines
li-gnées à très transformé pour d'autres. Le phénotype observé
est en relation avec le taux de production de T moyen. Le
plasmide bicistronique portant les gènes T moyen et néo nous a également permis de suivre l'expression d'un gène adjacent
lorsque l'expression d'un premier gène est modifiée. Nous
nous sommes également intéressés à la dynamique des
popula-tions cellulaires. Les lignées transformées génèrent des
ré-vertants non-transformés à une fréquence de l'ordre de
Quant aux lignées non-transformées portant des co-pies silencieuses du gène T moyen, elles peuvent générer des
transformants, dans lesquels T moyen est produit, avec une fréquence de l'ordre de
to-e.
Finalement, nous montrons que le grand T peut reproduire ce phénomène de conversion des cellules non-transformées, portant des copies silencieuses du gène T moyen, au phénotype transformé. Cette conversion s'accompagne, encore là, d'une stimulation de l'expression du gène T moyen. Il semblerait que, dans notre système, le grand T agisse indirectement en amenant des changements dans la structure chromatinienne à proximité du gène T moyen, le rendant plus accessible à la machinerie transcriptionnelle.MATERIEL ET METHODES
I. Souche virale:
Pour ce travail, nous avons utilisé la souche A2 du vi-rus du polyome, un variant du type sauvage produisant de grandes plaques <Fried et al., 1974>.
II. Plasmides recombinants:
Le recombinant pPA8 contient le plus grand des deux fragments Hindi!! du génome viral, cloné au site Hindi!! du plasmide pBR322. Ce fragment d'ADN viral, qui s'étend entre 44.6 et 1.8 unités sur la carte génomique, contient l'origi-ne de réplication ainsi que les séquences encodant les anti-gènes petit T, T moyen et l'extrémité N-terminale de grand T
(Bastin et al., 1980).
pPB21 contient le génome viral complet cloné au site BamHI de pBR322. Dans ce recombinant, la région précoce est ininterrompue et contient toutes les séquences encodant les antigènes petit T, T moyen et grand T (Gélinas et al., 1981).
pPtsa résulte d'une construction semblable où le mutant
tsa de polyome a été cloné dans pBR322 au site BamHI. La
lésion responsable de la mutation ts est une mutation ponc-tuelle qui fait que la guanine au nucléotide 2172 du génome
de type sauvage est remplacée par une adénine dans le génome du mutant <Thomas et al., 1981).
pdl8 contient le génome du mutant diS cloné au site BamHI de pBR322. Ce mutant est caractérisé par une délétion de 90 paires de bases, entre les nucléotides 990 et 1080, qui affecte les antigènes T moyen et grand T <Smolar et Gr i f f i n , 1 981> •
pMT3 est une forme réduite du plasmide pPyMT1 et a été obtenu en lui retirant deux fragments Hindiii <Asselin et al. , 1983). A la suite d'une délétion spécifique de l'in-tron du gène T moyen, ces deux recombinants encodent unique-ment la protéine T moyen <Treisman et al., 1981b).
pdl8MT3 a été obtenu en introduisant l'insert viral de pd18 au site BamHI de pMT3. Les deu:< régions précoces du recombinant sont dans le même sens transcriptionnel.
pSV2neo contient le gène néo, un marqueur de sélection dominant, sous le contrOle du promoteur précoce de SV40
<Southern et Berg, 1982).
pneoMT3 a été construit en insérant le fragment BamHI-EcoRI de la portion virale de pMT3 dans le plasmide pSV2neo
<Bouchard et al., 1986).
pneoLT1 résulte du clonage d'un génome viral modifié n'encodant que l'antigène grand Tau site BamHI de pSV2neo
<Asselin et al., 1986).
pneoLtsa est une construction dérivée de pneoLT1 obte-nue en transférant la mutation tsa dans pneoLT1.
délé-tian de 30 paires de bases allant du nucléotide 1367 au nu-cléotide 1396 <Asselin et al., 1986).
pneoLT3 porte un grand T tronqué à partir du site EcoRI situé au nucléotide 1560 du génome viral CAsselin et Bastin, 1985).
pMT97A8t a été construit en liant par l'intermédiaire
de leur unique site Hindiii, les recombinants pMT97 et A8t. Il en résulte un plasmide portant un tandem de deux régions précoces non transformantes affectées par les lésions diffé-rentes.
pMT97 a été obtenu en transférant la mutation dl97 dans
le recombinant pMT3 <Asselin et al., 1986).
A8t résulte de la recircularisation du plus grand des deux fragments obtenus en digérant pPA8 avec l'enzyme Pst!. Ce recombinant porte les séquences de la région précoce du
génome du virus du polyome à partir du site Pst! au
nucléo-tide 484 jusqu'au site Hindiii au nucléonucléo-tide 1656. Il porte en plus les séquences du plasmide pBR322 entre le site Hindiii au nucléotide 29 et le site Pst! au nucléotide 3612.
Ces différents recombinants ont été amplifiés dans la
souche bactérienne HB101, puis purifiés par
centri-fugation à l'équilibre en gradient de chlorure de césium tel
que décrit par Maniatis et al. (1982). Les différents
enzy-mes nécessaires à ce travail ont été obtenus d'Amersham ou
de Boehringer-Mannheim Canada et utilisés selon les recom-mandations des fabricants.
III. Induction de tumeurs recombinants:
par injection de plasmides
L'ADN linéarisé par un enzyme de restriction est
injec-té, de façon sous-cutanée, à la base du cou de rats Fischer
<Charles River Canada !ne.) le jour de leur naissance. Cha-que injection comprend 2,0 ug d'ADN plasmidiCha-que avec ou sans
ADN contenu dans 50ul de tampon phosphate <PBS
A: 10mM Na2HP04, 2mM KH2P04 pH 6,8>. Les rats sont
ensuite observés deux fois par semaine. Les tumeurs appa-raissent sous forme de masses palpables localisées au site d'inoculation.
IV. Etablissement des lignées tumorales:
Lorsque la tumeur atteint environ 3 cm de diamètre, elle est extraite de l'animal le plus stérilement possible.
La masse tumorale est rincée abondamment à trois reprises
dans du Tris-salin C140mM NaCl, 5mM KCl, 1mM Na2HP04, 5mM glucose, 25mM Tris, 0,0015/. rouge phénol pH 7,4> puis
réduite en petits morceaux à l'aide de ciseaux à dissection.
A deux reprises, on resuspend les morceaux dans du
Tris-salin et on les laisse sédimenter pendant cinq minutes
avant de décanter le liquide. La tumeur émincée est ensuite transférée dans un flacon à trypsinisation <Bellco>. On lui fait subir trois trypsinisations (100 ml de trypsine 0,06/.
dans du Tris-salin) avec agitation à 37°C. Après chaque
trypsinisation le liquide est décanté et filtré à travers
une mousseline stérile, ajouté à SOml de sérum de veau et conservé sur la glace. Après la troisième trypsinisation,
les cellules sont centrifugées pendant cinq minutes à 1000
rpm, puis resuspendues dans 10ml de DMEM <Dulbecco Modified Eagle Medium> contenant 107. de sérum de veau, 27. de fongizo-ne <amphotéricifongizo-ne B, 2,Sug par ml>, et 17. d'antibiotiques
<Streptomycine 0,017., pénicilline 100 unités par ml>. Une fraction de la suspension cellulaire est diluée 1:3 dans du bleu de trypan <O.SX bleu de trypan, 0,857. NaCl> avant que
les cellules soient comptées. La coloration au bleu de
trypan permet d'évaluer le nombre de cellules vivantes, ces dernières demeurant incolores, alors que le colorant pénètre les cellules mortes. Les cellules sont ensuite distribuées
dans des falcons de 75 cm2 à raison de 2x106 cellules
vivantes par récipient et conservées dans un incubateur à
37°C <SX C02>•
V. Analyse pathologique des tumeurs:
Au moment de prélever la tumeur, l'avoir rincée
au Tris-salin, on prélève une tranche de tissu de quelques
millimètres d'épaisseur qui est fixée à la formaldéhyde,
avant d'être recouverte de paraffine, sectionnée et colorée
à L'analyse _pathologique des
dé-partement de pathologie de l'Université de Sherbrooke. VI. Transfection de cellules en culture:
Pour ce travail, nous avons utilisé la lignée
cellulai-re FR3T3. Cette lignée de cellules de rat a été établie
dans le laboratoire du Dr François Cuzin à l'Université de
Nice (Seif et Cuzin, 1977>. Elle est propagée dans du DMEM contenant 10/. de sérum de veau fétal, 2/. de fongizone, et 1/. d'antibiotiques <DMEM complet>, dans un incubateur à 37°C
dont l'atmosphère contient 5/. de La méthode de
transfection utilisée est celle au chlorure de calcium-DMSO décrite par Stow et Wilkie (1976>. Brièvement, les cellules
sont ensemencées la veille de l'expérience à raison de
5x10e cellules par pétri de 60mm. Le jour de la
transfec-tion, on ajoute à l'ADN à transfecter, contenu dans un
volu-me de 25ul de TE <Tris 10mM, EDTA 1mM pH B,O>, 450ul d'une
solution d'ADN de Thymus de veau à 10ug par ml de tampon
Hebs <140mM NaCl, 5mM KCl, 0,7mM Na2HP04, 5,5mM glucose, 20mM Hepes pH 7,1>, 100ul d'aprotinine (Sigma) et 35ul de CaCl2 2M. On agite immédiatement au vortex et on laisse reposer 10 minutes pendant lesquelles un précipité de
phos-phate de calcium commence à se former. Ensuite, on retire
le milieu couvrant les cellules et on y applique le
précipi-té après l'avoir de nouveau agiprécipi-té au vortex. Les pétris
sont incubés durant 30 minutes à 37°C et agités aux dix
milieu contenant tous les additifs habituels à l'exception de la fongizone. Nous utilisons du milieu dépourvu de
fon-gizone à partir de cette étape car nous avons remarqué à
plusieurs reprises que la fongizone peut interférer dans la formation du précipité de phosphate de calcium. Il en ré-sulte un précipité beaucoup plus gros qui tue rapidement les
cellules. Les cellules sont à nouveau incubées à
pour une période de cinq à six heures après laquelle on
pro-cède au choc au DMSO. Pour ce faire, on retire le milieu des pétris et on rince les cellules avec 5ml de milieu sans
fongizone. On dépose ensuite sur les cellules 1ml de DMEM
contenant 20/.
<vlv>
de DMSO. Cette solution est laisséesur les cellules pendant 40 secondes, après quoi on rince
les cellules à deux reprises avec 5ml de milieu complet,
toujours sans fongizone. On recouvre finalement les cellu-les avec 5ml du m@me milieu et cellu-les pétris sont incubés
pen-dant 36 heures à dans une atmosphère contenant 5/. de
Après cette période, la façon de traiter les cellu-les dépend du type de sélection que l'on désire effectuer.
Dans le cas l'on veut obtenir des foyers de cellules
transformées, le contenu de chaque est dépourvu de son
milieu, rincé avec 5ml de versène <0,5mM EDTA, 0,0015/. rou-ge phénol dans du PBS A>, et dispersé à la trypsine (0.06/.
dans du versène>. Les cellules sont redistribuées à raison
de 5x104 cellules par pétri de 60mm dans du milieu qui
peut maintenant contenir de la fongizone. Le milieu est
transformées apparaissent sur la mosaïque des cellules
nor-males de 14 à 21 jours plus tard. Lorsqu'on veut
sélec-tionner pour l'acquisition d'un marqueur de sélection domi-nant, les cellules de chaque pétri sont redistribuées dans
cinq nouveaux pétris contenant du milieu complet. De 12 à
18 heures plus tard, on applique la sélection. Si on veut
sélectionner pour l'expression du gène ngQ, on remplace le
milieu par du milieu contenant 400ug par ml de
l'antibioti-que 6418 <6418 sulfate, 6ibco>. Ce milieu est préparé à
partir d'une solution mère contenant 4mg de 6418 par ml de tampon Hepes 100mM pH 7,4. Si on désire sélectionner pour l'expression du gène hvgro, le milieu de sélection contient lOOug par ml d'hygromycine <Calbiochem>, et on le prépare à
partir d'une solution mère contenant 10 mg d'antibiotique par ml de PBS A. Dans les deux cas, le milieu de sélection est changé aux 5 jours et les colonies résistantes apparais-sent de 2 à 3 semaines plus tard.
VII. Clonage en agar noble:
Cette technique permet de tester l'indépendance
d'an-crage des lignées cellulaires en les soumettant à un milieu
de croissance semi-solide. La technique employée est celle de Monteil et al. (1984). En bref, 10,000 cellules conte-nues dans 3,5ml de milieu complet ayant une teneur de 0,33/. en agar noble <Difco> sont ensemencées entre deux couches de milieu contenant 0,66/. d'agar noble dans un pétri de 60mm et
incubées à Une fois par semaine, on alimente les cellules en renouvelant la couche supérieure de milieu cam-plet contenant 0,66/. d'agar noble. Les colonies visibles
macroscopiquement sont dénombrées de trois à quatre semaines
plus tard.
VIII. Evaluation du transformées:
potentiel tumorigène des cellules
Les cellules à tester sont dispersées à la trypsine,
centrifugées dans 10ml de DMEM complet pendant cinq minutes
à 1800rpm, lavées dans 10ml de PBS A de façon à obtenir
50,000 cellules par injection de 50 à 200ul de la suspension
cellulaire. Les cellules sont injectées dans le flanc de
jeunes rats Fischer agés d'environ un mois ou encore de
souris nues adultes <nu/nu, Flanagan, 1966; Pantelouris,
1968). On surveille ensuite 1 'apparition d'une masse
tumo-raie aux sites d'injection.
IX. Mesure du taux d'apparition des variants dans les populations cellulaires:
Pour ce faire, nous avons procédé à des analyses par
test de fluctuation selon la méthode de Luria et Delbruck
(1943). Brièvement, cette méthode consiste à obtenir un
grand nombre de cultures indépendantes, chacune issue d'un
de façon à obtenir une grande population finale et on dénom-bre les variants apparus dans chaque population. Pour con-tourner le problème causé par la possibilité de dénombrer plusieurs variants originant d'un seul et même événement mu-tatienne!, le taux d'apparition des variants est évalué à partir du nombre de cultures dans lesquelles il n'y a pas de variant <Po>. Le nombre de mutations correspond à: -lnPo. Le nombre de générations cellulaires dans une po-pulation non synchronisée est donné par
et correspondent respectivement au nombre final et initial de cellules dans la culture. Le taux d'apparition des variants, exprimé en mutations par génération cellulaire est donné par l'équation
1. Taux de conversion des cellules non-transformées au phénotype transformé:
Les cellules sont ensemencées dans des plaques de 96 puits de 6mm (Linbro) à raison d'une vingtaine de cellules par puits Chaque puit représente une culture in-dépendante. Nous avons évalué qu'à confluence, chacun con-tient en moyenne 18,000 cellules Les cellules arri-vent à confluence environ cinq semaines après l'ensemence-ment. Huit semaines après le début de l'expérience, on éva-lue la proportion des cultures dans lesquelles il n'y a pas eu apparition de cellules transformées <Po>.
2. Taux de conversion des cellules transformées au phénotype normal:
Les cellules transformées sont d'abord sous-clonées avant d'être ensemencées à faible densité dans des pétris de 100mm (5 à 10 cellules par cm2 ). Des colonies isolées,_
issues d'une seule cellule transformée sont préle-vées. Chaque colonie représente une culture indépendante. Chacune est dispersée en cellules isolées et réensemencée dans un pétri de 100mm de façon à ce que chaque cellule de la colonie mère puisse être dénombrée et identifiée d'après la morphologie de la colonie cellulaire à laquelle elle a donné origine. La valeur du correspond au nombre de colonies recensées alors que le Po correspond à la propor-tion des cultures indépendantes on n'a dénombré aucune colonie révertante.
3. Taux de conversion des cellules sensibles au 6418 en cellules résistantes:
Les lignées sont amplifiées à partir d'une seule cellu-le dans des puits de 6mm puits contient donc une culture indépendante. A confluence chacune est transfé-rée dans un puits de 15mm pour en continuer l'amplification. Peu avant qu'une culture n'atteigne à nouveau la confluence, ses cellules sont dispersées, comptées pour obtenir la va-leur de et transférées dans un pétri de 100mm. La
sé-lection est appliquée douze heures plus tard alors que le milieu couvrant les cellules est remplacé par du milieu con-tenant 400ug par ml de 8418. Le milieu de sélection est
changé tous les cinq jours. Deux à trois semaines plus
tard, les colonies résistantes au 8418 peuvent étre dénom-brées. La proportion des cultures ne contenant pas de colo-nie résistante au 8418 donne la valeur du Po.
4. Evaluation de la valeur statistique du test de fluc-tuation:
Dans le cadre des tests de fluctuation, le nombre de variants dans une culture va dépendre du moment où le ant initial est apparu. Dans certaines cultures les
vari-ants tôt et auront le temps de se diviser avant
qu'on les dénombre. Dans d'autres, ils plus
tard, n'auront pas nécessairement le temps de se diviser et seront moins nombreux. Dans d'autres cultures, enfin, il n'y aura pas de variant. La valeur statistique de la fluc-tuation observée peut ètre déterminée en la comparant avec une série d'échantillons tirés d'une culture contrôle déjà à
l'équilibre. La fluctuation dans le nombre de variants
de-vrait ètre beaucoup plus grande dans les populations clona-les du test que dans la population contrôle. Mathématique-ment, ceci s'exprime par le rapport de la variance sur la
moyenne. Cette valeur devrait ètre beaucoup plus grande
contre-le où elcontre-le devrait s'approcher de 1. Nous nous sommes assu-rés de rencontrer ces conditions pour nos différents tests de fluctuation. Un exemple est donné dans la légende du ta-bleau 10.
X. Mise en évidence transformées:
des antigènes T dans les lignées
1. Immunofluorescence: <Basilico et al., 1970>
On ensemence les cellules sur une lame porte-objet.
Lorsqu'elles atteignent 307. de confluence, on les lave à
trois reprises dans du tampon TBS à température de la pièce
(25mM Tris, O,SmM MgC12,140mM NaCl, SmM KCl, 0,7mM
CaCl2, pH 7,5>. Elles sont ensuite placées durant 15
mi-nutes à -2occ dans un mélange acétone:méthanol (2:1).
Cette étape sert à fixer et à perméabiliser les cellules.
On les laisse ensuite sécher pendant 30 minutes à la
tempé-rature de la pièce. On vérifie alors l'aspect des cellules
et on délimite sur la lame un champ à traiter. Les
cellu-les sont lavées trois fois dans du TBS à température de la
pièce. On dépose sur la zone choisie une goutte de sérum
anti-polyome dilué dans du TBS <1:10) et on incube 60
minu-tes à la température de la pièce dans un environnement
hu-mi de. Les lames sont ensuite lavées à nouveau trois fois
dans du TBS. Après avoir asséché le contour de la zone
marqué à la fluorescéine <Calbiochem> et dilué dans du TBS (1:60). On incube comme précédemment durant 60 minutes. On
lave à nouveau les lames à trois reprises dans du TBS. On
dépose sur le champ choisi une goutte d'un mélanoe
TBS:gly-cérol <1:1) et on y superpose une lamelle. Les cellules
peuvent alors être observées au microscope à lumière
ultra-violet. On peut également conserver les lames dans l'obscu-rité à 4°C.
2. Détection de l'activité kinasique associée à
l'antigène T moyen (in vitro):
i. Immunoprécipitation des antigènes T, marquage et électrophorèse:
Cette technique consiste à immunoprécipiter les
protéi-nes précoces de polyome présentes dans les lignées
cellulai-res, puis à incuber ces protéines en présence d'ATP
radioac-tif marqué On peut, après électrophorèse et
autora-diographie, détecter les protéines qui auront été marquées
suite à la réaction de phosphorylation in vitro. La
métho-de que nous employons est celle qui a été décrite par Schaf-fhausen et Benjamin <1981a). Lorsque les cellules, en
cul-ture dans des pétris de 60mm, arrivent à 807. de confluence,
on retire le milieu et on rince les cellules deux fois (5ml par lavage> avec du tampon PBS- froid (170mM NaCl, 3,3mM
pH 6,8>. On procède à l'extraction des protéines en ajoutant 1ml de tampon TEB C120mM NaCl , 18mM Tris- HCI
pH 7,0, 0,8mM 0,45mM 17. (v/v) NP40, 107.
Cv/v) glycérol). Les pétris sont incubés pendant 20 minutes à 4°C, et agités à intervalles réguliers de 5 minutes, après quoi, les lysats sant recueillis et transférés dans des tubes de 1,5ml. Les extraits sont centrifugés durant 5 minutes afin de les débarasser des débris cellulaires. Cha-que surnageant est transféré dans un tube coniCha-que de 15ml
<Starstedt> contenant déjà 40ul de protéine A-sépharose <Pharmacia, 507.
<vlv>
dans de l'eau bidistillée) et 2ul de sérum anti-polyome <19-2, Pallas et al., 1986). Les tubes sont maintenus sur la glace pendant 60 minutes, et réguliè-rement , on les agite doucement de façon à garder la protéi-ne A-sépharose en suspension. Pendant cette période d'immu-noabsorption les complexes antigène-anticorps vont être re-tenus par la protéine A-sépharose. On procède ensuite à une série de lavages ayant pour but d'éliminer ce qui se serait lié à la sépharose de façon non-spécifique. La sépharose est donc lavée deux fois au PBS+ froid, deux fois avec un tampon de lavage au chlorure de lithium froid <0,5M LiCl, 0,1M Tris-HCl pH 7,0> et une fois avec le tampon de kinase(5mM 20mM Tris pH 7,5> à la température de la
piè-ce. Chacun des lavages consiste à ajouter 5ml de tampon par tube, de façon à bien disperser la protéine A-sépharose. La sépharose est récupérée en centrifugeant les tubes jusqu'à 2000rpm et en arrêtant la centrifugeuse <HB-4, Sorvall) dès
qu'elle atteint cette vitesse. Après chaque centrifugation, le surnageant est enlevé par aspiration. Après cette série
de lavages, on peut procéder à la réaction de kinase in
vitro qui se fait dans un volume de 400ul de tampon de
kina-se contenant lOOuCi de par réaction. On laisse
la réaction procéder pendant 15 minutes à la température de
la pièce, en agitant doucement aux cinq minutes de façon à
garder la sépharose en suspension. On lave ensuite la sé-pharose, comme précédemment, deux fois au PBs- froid, deux fois avec le tampon de lavage au chlorure de lithium froid, deux fois avec du tampon RIPA froid <150mM NaCl, !OmM Tris pH 7,5, EDTA lmM, 1/. NP40, 1/. déoxycholate de sodium, 0,17. SDS) et finalement, deux fois dans de l'eau bidistillée froide. Les échantillons ainsi lavés sont conservés sur la glace jusqu'à ce qu'on les prépare pour l'électrophorèse.
Juste avant de déposer les échantillons sur gel, on ajoute à
chacun 30ul de tampon de dissociation (5/. Cp/v) SDS, 5/. Cv/v) 2-mercaptoéthanol, 20/.
<vlv>
glycérol, 125mM Tris pH 6,8, 0,0075/. Cp/v) bleu de bromophénol et on fait bouillir pendant deux minutes. Il est important d'ajouter le tamponde dissociation à la toute dernière minute, puisque le SDS
qu'il contient rend les protéines plus sensibles aux
protéa-ses. On fait migrer les échantillons sur un gel
d'acryla-mide 12/., dans un tampon 25mM Tris, 192mM glycine, 0,1/. SDS (pH 8,3 sans ajustement>, en présence de protéines-marqueur précolorées <BRL, 14,300-200,000 daltons>. La migration se fait à 70 volts durant une nuit.
ii. Traitement des gels pour autoradiographie:
Après l'électrophorèse, on trempe le gel pendant 30 mi-nutes dans une solution contenant 7,5/. <vlv> d'acide acéti-que et 5/. (v/v) de méthanol, ceci afin de fixer les
protéi-nes. Ensuite, on sèche le gel durant 90 minutes sous vide
avant de l'exposer à un film Kodak RP1 à -70°C en présence
d'un écran. Alternativement, on peut, avant d'exposer le
gel, le traiter de façon à mettre en évidence la
phosphory-lation préférentielle des résidus tyrosine. En effet, grâce
à son noyau aromatique, la tyrosine demeure stable en milieu
alcalin, alors que les résidus sérine et thréonine qui ne portent pas de noyau aromatique sont moins stables. Il en
résulte qu'en milieu alcalin le lien entre un groupement
phosphate et un résidu tyrosine demeure stable alors que le lien entre un groupement phosphate et un résidu sérine ou thréonine est hydrolysé. Dans ce cas, après l'électro-phorèse, le gel est fixé pendant une heure dans une solution contenant 7,5/. d'acide acétique et 5/. de méthanol, puis
rin-cé à l'eau. Il est ensuite incubé durant 2 heures à 55°C
dans une solution d'hydrolyse (lM NaOH, 10mM KzHP04>,
après quoi, il est de nouveau rincé à l'eau. Il est ensuite
traité 30 minutes dans une solution contenant 15/. d'acide acétique et 50/. de méthanol, puis 30 autres minutes dans une solution contenant 7,5/. d'acide acétique et 5/. de méthanol. Finalement, le gel est trempé 10 minutes dans du glycérol 1/.
Cv/v), avant d•être séché et exposé comme précédemment.
XI. Caractérisation du patron d'intégration de l'ADN viral dans 1·ADN des lignées cellulaires:
1. Extraction de l'ADN cellulaire:
Les lignées dont on veut extraire l'ADN sont cultivées dans des pétris de 100mm jusqu'à confluence. Le milieu est alors retiré et les cellules sont rincées à deux reprises dans du PBS A préchauffé à 37°C. Les cellules sont ensui-te lysées par l'addition de lml de tampon A (10mM NaCl, lOmM EDTA, lOmM Tris,
o,s;.
SDS, pH 7,9> contenant SOug par ml de protéinase K, par pétri. Le tampon contenant la protéinase K est incubé pendant une heure à 37oc avant d'être ajouté aux cellules, ce qui permet à la protéinase K, qui agit ici comme inhibiteur des DNases, de s'autoactiver. Les pétris sont agités doucement, puis on laisse agir le tampon durant 10 minutes à la température de la pièce, après quoi, on re-cueille les lysats à l'aide d'un policeman et on les trans-fère dans des tubes stériles. On laisse ensuite agiter dou-cement durant 12 heures à 37°C ou toute une.nuit à latem-pérature de la pièce. On ajoute ensuite un volume de phénol contenant 0.17. de 8-hydroxyquinoline et saturé en TE, pour extraire les protéines du mélange. Le tout est agitépen-dant une à la température de pièce, puis