Libération vectorielle du virus de l'hépatite E dans un modèle d'hépatocytes polarisés in vitro et optimisation de la culture pour la quantification de son infectiosité
197
0
0
Texte intégral
Figure
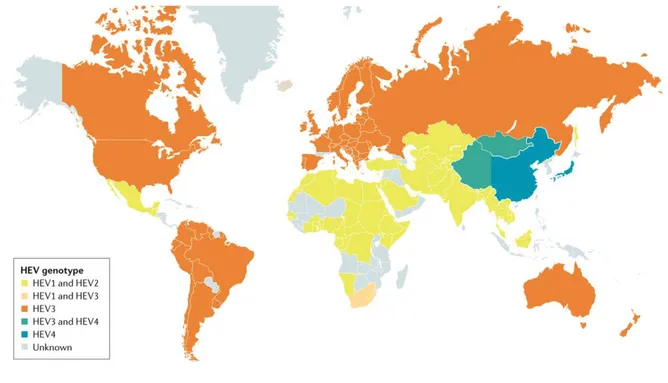
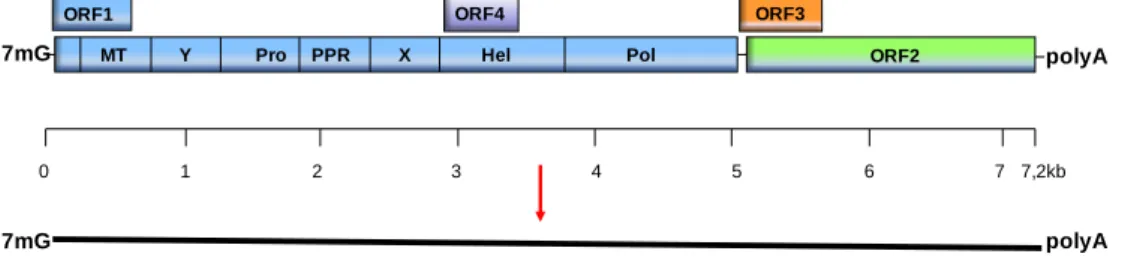
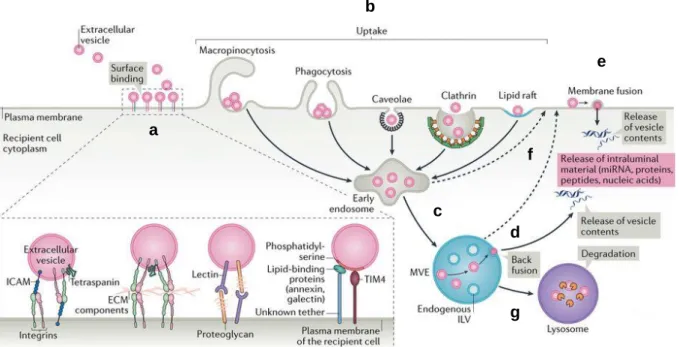

+5
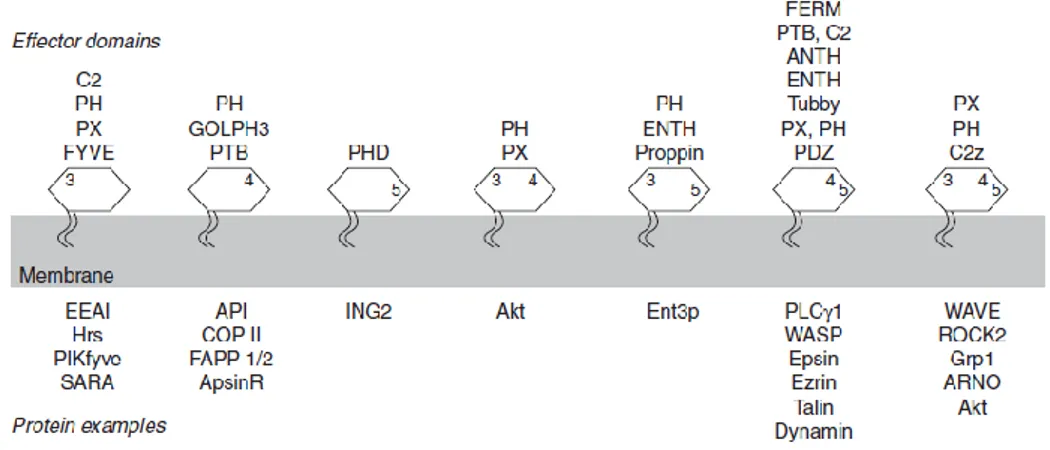



Documents relatifs