i
DELPHINE BOYER GROULX
CARACTÉRISTIQUES MORPHO-PHYSIOLOGIQUES DES
FAMILLES BIPARENTALES D’ÉPINETTE BLANCHE (PICEA
GLAUCA (MOENCH) VOSS) EN RÉPONSE AUX
CHANGEMENTS CLIMATIQUES : CAS DE
L’AUGMENTATION DE LA TEMPÉRATURE ET DE LA
CONCENTRATION EN CO
2ET DE LEUR INTERACTION
Mémoire présenté
à la Faculté des études supérieures et postdoctorales de l’Université Laval dans le cadre du programme de maîtrise en Sciences forestières
pour l’obtention du grade de Maître ès sciences (M. Sc.)
DÉPARTEMENT DES SCIENCES DU BOIS ET DE LA FORÊT FACULTÉ DE FORESTERIE, DE GÉOGRAPHIE ET DE GÉOMATIQUE
UNIVERSITÉ LAVAL QUÉBEC
2012
ii
Résumé court
Selon le scénario d’émission A1B défini par le Groupe d’experts intergouvernemental sur l’évolution du climat (GIEC), la concentration atmosphérique en CO2 pourrait
doubler d’ici 2100 et, au Canada, la température moyenne pourrait augmenter de 4,3 °C. Dans une expérience en conditions contrôlées, des plants d’épinette blanche, issus des meilleures familles biparentales identifiées par le programme d’amélioration génétique du Québec, ont été soumis à une combinaison de deux régimes de température (actuelle vs. élevée) et de deux régimes de CO2 (380 vs. 760 ppm) pendant deux saisons de
croissance.
L’interaction entre l’augmentation de la température et le CO2 n’a pas eu d’effet
significatif sur la croissance, la nutrition minérale, les échanges gazeux et les caractéristiques foliaires des plants au stade juvénile. Par ailleurs, la performance relative entre les 20 familles n’a pas été modifiée, suggérant qu’elles devraient maintenir leur performance supérieure en présence des conditions climatiques futures prédites.
iii
Abstract
According to the A1B emissions scenario of the Intergovernmental Panel on Climate Change’s (IPCC) 2007 report, atmospheric CO2 concentration will double before 2100
and, in Canada, average temperatures could increase by 4.3 °C. In a growth chamber experiment, we subjected white spruce seedlings from the best performing full-sib families in Québec’s current tree breeding program to a combination of two temperature regimes (present day vs. anticipated future temperatures) and two levels of CO2 (380 vs. 760 ppm) during two growing seasons.
The interaction between CO2 and temperature increases had no significant effect on
growth characteristics, mineral nutrition, gas exchange and needle characteristics of seedlings at the juvenile stage. Furthermore, the relative performance between 20 families did not change, suggesting that they should maintain their superior performance under predicted future climate conditions.
iv
Résumé long
Les programmes d’amélioration génétique visent la sélection d’arbres supérieurs qui serviront à établir des plantations de haute productivité adaptées aux conditions climatiques actuelles et futures. Or, les conditions climatiques futures seront vraisemblablement différentes des conditions actuelles. En effet, selon le scénario d’émission A1B, défini par le Groupe d’experts intergouvernemental sur l’évolution du climat (GIEC), la concentration atmosphérique en CO2 pourrait doubler à 760 ppm
d’ici 2100 et, au Canada, la température moyenne pourrait augmenter de 4,3 °C (GIEC 2007). À cet égard, les programmes de reboisement devraient tenir compte de ces changements climatiques en évaluant les réponses morpho-physiologiques des différentes sources génétiques de semences vis-à-vis des principales variables environnementales (CO2 et température) et de leur interaction.
Dans une expérience en conditions contrôlées (température, CO2, humidité relative,
lumière, photopériode, etc.), des plants d’épinette blanche, représentant les meilleures familles biparentales identifiées par le programme d’amélioration génétique du Québec, ont été soumis à une combinaison de deux régimes de températures (actuelles vs. futures prédites) et de deux régimes de CO2 (380 vs. 760 ppm) pendant leur seconde et
leur troisième saison de croissance. La croissance des parties aériennes et des racines, ainsi que la nutrition minérale ont été évaluées à la fin de la deuxième et de la troisième saison de croissance. Des mesures d’échanges gazeux, de transpiration cuticulaire, de quantité de cire cuticulaire ainsi que de surface foliaire spécifique ont également été effectuées sur les plants âgés de trois ans.
L’augmentation de la température et de la concentration en CO2 a eu peu d’effet sur les
variables morpho-physiologiques des plants de deux ans (2+0) et de trois ans (3+0) d’épinette blanche. Ainsi, l’élévation seule de la température ou de la concentration en CO2 s’est traduite par une augmentation de la hauteur des plants (2+0) ainsi que du
diamètre et de la masse sèche des racines des plants (3+0). Des différences entre les familles ont été observées pour la majorité des variables morpho-physiologiques des
v plants (2+0) et (3+0), c’est-à-dire pour toutes les variables de croissance, la nutrition minérale, la transpiration ainsi que la conductance stomatique. Aucune interaction famille*traitement n’ayant été significative, les résultats de cette expérience indiquent que la performance relative entre les familles d’épinette blanche ne devrait pas être modifiée suite à une augmentation de la température et de la concentration en CO2. Nos
résultats suggèrent que les meilleures familles actuellement sélectionnées dans le cadre du programme d’amélioration génétique de l’épinette blanche au Québec devraient maintenir leur performance supérieure en présence des conditions climatiques futures prédites. De plus, l’héritabilité familiale s’est révélée similaire et élevée (supérieure à 0,4) entre les traitements de température * CO2, ce qui signifie que les traitements n’ont
vi
Avant-propos
Je tiens à remercier mon directeur de maîtrise Dr. Hank Margolis, professeur titulaire de l’Université Laval, ainsi que mon co-directeur Dr. Mohammed S. Lamhamedi, chercheur scientifique à la Direction de la recherche forestière (DRF) du ministère des Ressources naturelles et de la Faune (MRNF) du Québec pour leur support, leurs conseils ainsi que pour leur encadrement.
Je tiens à remercier le personnel du Centre de Foresterie des Laurentides (CFL) impliqué au sein du projet, Jean Beaulieu, Pierre Bernier, Marie Deslauriers, Yves Dubuc et Alain Labrecque. Je remercie également Madame Josianne De Blois, statisticienne au Service du soutien scientifique (DRF, MRNF), l’équipe technique (MM. Mario Renaud et Pascal Desjardins, ainsi que Mme Linda Veilleux) du Dr. Mohammed S. Lamhamedi (DRF, MRNF), ainsi que le laboratoire de chimie organique et inorganique (DRF, MRNF) pour les analyses minérales des plants et des substrats.
L’aide du personnel et des étudiants de l’Université Laval a grandement été appréciée, Debbie Christiansen-Stowe, Carole Coursolle, Marie Coyea, Vincent Garneau, François Larochelle, Steeve Pépin pour son aide avec le LI-COR, Étienne St-Pierre et Nadya Wahid. Je veux remercier particulièrement Sylvie Carles qui m’a offert soutien, conseils et disponibilité.
Le soutien financier a été assuré par le Fonds de recherche du Québec - Nature et technologies (FRQNT), le Conseil de recherches en sciences naturelles et en génie du Canada (CRSNG), la DRF, ainsi que le Service canadien des forêts (Québec, Canada). Cette maîtrise est réalisée dans le cadre du projet 2009-FT-130171 qui s’intitule : «Évaluation de la plasticité physiologique des familles d’épinette blanche en vue de maximiser la productivité des plantations en réponse aux changements climatique» (lien : http://www.mrn.gouv.qc.ca/publications
vii Mes derniers remerciements vont à ma mère, mon père et ma sœur qui m’ont toujours encouragée à me dépasser dans mes études. Merci à tous mes amis pour leur soutien moral et à Mathieu de croire en moi.
viii Nous n’héritons pas la terre de nos parents,
nous l’empruntons à nos enfants [Antoine de Saint-Exupéry]
ix
Table des matières
Résumé court...ii
Abstract ... iii
Résumé long...iv
Avant-propos...vi
Table des matières...ix
Liste des tableaux...xi
Liste des figures ... xii
Introduction générale ... 1
Objectifs et hypothèses de recherche ... 3
Objectifs ... 3
Hypothèses ... 3
Références ... 4
CHAPITRE 1 : REVUE DE LITTÉRATURE ... 5
1.1. Changements climatiques ... 5
1.1.1. Changements climatiques observés ... 5
1.1.2. Changements climatiques envisagés ... 6
1.1.3. Stratégies et politiques ... 9
1.2. Effets des changements climatiques sur les caractéristiques morpho-physiologiques des arbres ... 13
1.2.1 Rappel sur la photosynthèse... 13
1.2.2. CO2... 16
1.2.3. Température ... 26
1.2.4. Effets simultanés de la température et de la concentration en CO2 sur les caractéristiques morpho-physiologiques... 30
1.3. Génétique ... 37
1.3.1. État des connaissances ... 37
1.4. Conclusion générale ... 56
1.5. Références ... 59
CHAPITRE 2 : EFFETS DE L’AUGMENTATION DE LA TEMPÉRATURE ET DE LA CONCENTRATION EN CO2 ET DE LEUR INTERACTION SUR LES CARACTÉRISTIQUES MORPHO-PHYSIOLOGIQUES DE FAMILLES BIPARENTALES D’ÉPINETTE BLANCHE (PICEA GLAUCA (MOENCH) VOSS) .... 66
2.1. Introduction ... 66
2.2. Matériel et méthodes ... 71
2.2.1. Matériel génétique... 71
2.2.2. Production des plants (1+0) en serre... 71
2.2.3. Application des traitements de CO2 (C) et de température (T) en conditions contrôlées pendant les deuxième et troisième saisons de croissance... 72
2.2.4. Caractérisation morphologique et nutrition minérale des plants 2+0 et 3+0 ... 76
2.2.5. Caractérisation physiologique des plants 3+0... 76
x 2.3.1. Effets des traitements de température et de CO2 sur les caractéristiques
morpho-physiologiques des plants d’épinette blanche ... 82
2.3.2. Effets des familles sur les caractéristiques morpho-physiologiques... 83
2.3.3. Corrélations entre les moyennes familiales des caractéristiques morpho-physiologiques des plants d’épinette blanche (3+0) ... 89
2.3.4. Regroupement des familles effectué par le test de Scott-Knott ... 89
2.3.5. Héritabilités familiales ... 93
2.4. Discussion ... 95
2.4.1. Effet de la température et du CO2 sur les caractéristiques morphologiques ... 95
2.4.2. Effet de la température et du CO2 sur les caractéristiques physiologiques... 96
2.4.3. Effet des familles sur les caractéristiques morpho-physiologiques ... 98
2.4.4. Interaction entre les traitements de température et de CO2 et les familles ... 99
2.5. Conclusion ... 100
2.6. Références ... 101
CHAPITRE 3 : CONCLUSION GÉNÉRALE ... 106
3.1. Sommaire des résultats ... 106
3.2. Implications... 107
3.3. Limites de l’étude... 108
3.4. Recommandations pour les recherches futures ... 110
3.5. Références ... 112
Annexe 1 ... 113
xi
Liste des tableaux
Tableau 1.1. Projections des valeurs moyennes du réchauffement en surface à l’échelle du globe pour la période 2090-2099. (GIEC 2007)... 8 Tableau 1.2. Synthèse de quelques travaux sur les effets d’une augmentation de la
concentration en CO2 sur les caractéristiques de croissance, les échanges gazeux et les
caractéristiques foliaires de jeunes plants d’essences résineuses. ... 23 Tableau 1.3. Synthèse de quelques travaux sur les effets d’une augmentation simultanée de la
température et du CO2 sur les caractéristiques de croissance, les échanges gazeux et les
caractéristiques foliaires de différentes espèces ligneuses. ... 33 Tableau 1.4. Synthèse de quelques travaux sur les différences de performance entre les
provenances d’épinette blanche de l’Est de l’Amérique du Nord en termes de caractéristiques morphologiques. ... 41 Tableau 1.5. Synthèse de quelques travaux sur les différences de performance entre les familles
d’épinette blanche de l’Est de l’Amérique du Nord en termes de caractéristiques morpho-physiologiques. ... 43 Tableau 1.6. Synthèse de quelques travaux sur les différences de performance en termes de
caractéristiques morpho-physiologiques entre les provenances, les familles et les clones de conifères en réponse à l’augmentation de la température et/ou de la concentration en CO2. ... 48
Tableau 2.1. Moyennes, erreur-types et moyennes familiales minimale et maximale des caractéristiques morphologiques de plants (2+0) et (3+0) représentant 20 familles biparentales d’épinette blanche et ayant été soumis à la combinaison de deux niveaux de traitements de température (température actuelle : moyennes pour 1990-2000 vs. température élevée : moyennes prédites pour 2090-2100) et de CO2 (CO2 actuel : 380 ppm
vs. CO2 élevé : 760 ppm) pendant leur deuxième et troisième saisons de croissance et
résultats des analyses statistiques testant les effets simples des traitements. ... 86 Tableau 2.2. Moyennes, erreur-types et moyennes familiales minimale et maximale des
caractéristiques physiologiques de plants (3+0) représentant 20 familles biparentales d’épinette blanche et ayant été soumis à la combinaison de deux niveaux de traitements de température (température actuelle : moyennes pour 1990-2000 vs. température élevée : moyennes prédites pour 2090-2100) et de CO2 (CO2 actuel : 380 ppm vs. CO2 élevé : 760
ppm) pendant leur deuxième et troisième saisons de croissance et résultats des analyses statistiques testant les effets simples des traitements... 87 Tableau 2.3. Corrélations entre les moyennes familiales des caractéristiques
morpho-physiologiques de plants (3+0) de 20 familles biparentales d’épinette blanche soumis à des traitements de température et de CO2 pendant deux saisons de croissance
consécutives. ... 90 Tableau 2.4. Moyenne des 60 familles (3 plants par famille) (et erreur-type), pourcentage de la
variance totale attribuable aux familles (%σ2
fam) et héritabilité familiale (h2fam et
erreur-type) des caractéristiques de croissance de plants (3+0) d'épinette blanche soumis, en conditions contrôlées, aux conditions actuelles de température et de CO2 (Ta Ca) ou à une
augmentation de la température et de la concentration en CO2 (Te Ce) durant deux saisons
xii
Liste des figures
Figure 1.1. Scénarios d'émissions (lignes pleines) de GES pour la période 2000-2100 en l'absence de politiques climatiques additionnelles. La zone en gris représente l’intervalle au 80e percentile des scénarios publiés (fourchette post-SRES) et les lignes en pointillés
délimitent la plage complète des scénarios (GIEC 2007). ... 7 Figure 1.2. Projections relatives aux températures à la surface du globe pour la période
2000-2100, la portion foncée des barres à droite du graphique représentant la valeur la plus probable et celle plus pâle précisant l’intervalle probable. (GIEC 2007). ... 7 Figure 1.3. Émissions mondiales de GES, par pays, en mégatonnes (Mt) d’équivalent de CO2
pour 2005. Adapté de Anonyme (2012). ... 11 Figure 1.4. Émissions totales de GES au Canada, par secteur économique, en mégatonnes (Mt)
pour 2005. Adapté de Anonyme (2012). ... 11 Figure 1.5. Processus de la photosynthèse. ... 15 Figure 1.6. Chambre de croissance. ... 18 Figure 1.7. Chambres à ciel ouvert (OTC). http://www.wsl.ch/ozone/ICP-Forests_2001.ehtml. .... 18 Figure 1.8. Enceinte à branches (branch bags), pour la mesure d’échanges gazeux. (Saugier
1999) ... 18 Figure 1.9. Système FACE. http://www.bnl.gov/face/faceProgram.asp. ... 18 Figure 1.10. Organigramme des principaux effets des changements climatiques sur les
caractéristiques morpho-physiologiques et la variabilité familiale des espèces ligneuses. ... 58 Figure 2.1. Variations saisonnières (1er mai au 30 septembre) de la photopériode et des
températures moyennes journalières actuelles et futures (élevées) programmées entre 7 h et 13 h dans les chambres à environnement contrôlé. ... 75 Figure 2.2. a) Différence entre les moyennes des traitements de température actuelle (moyenne
des températures de 1990-2000) vs. température élevée (températures futures prédites pour 2089-2099) et entre b) les moyennes des traitements de CO2 actuel (380 ppm) vs.
CO2 élevé (760 ppm) pour le diamètre ainsi que la masse sèche des racines des plants
(3+0) de 20 familles biparentales d’épinette blanche suite à une exposition aux traitements durant deux saisons de croissance. ... 88 Figure 2.3. Moyennes familiales de la hauteur et les masses sèches des parties aériennes et des
racines de plants (3+0) de 20 familles biparentales d’épinette blanche soumis à des traitements de température et de CO2 pendant leur deuxième puis troisième saisons de
croissance. Les familles ont été classées par ordre croissant de masses sèches des parties aériennes. ... 91 Figure 2.4. Moyennes familiales de la capacité photosynthétique, la transpiration et la
conductance stomatique à l’eau de plants (3+0) de 20 familles biparentales d’épinette blanche soumis à des traitements de température et de CO2 pendant leur deuxième puis
troisième saison de croissance. Une corrélation positive a été trouvée significative entre la transpiration et la conductance stomatique. Les familles ont été classées par ordre de transpiration croissante... 92
1
Introduction générale
L’épinette blanche est une essence largement répandue en Amérique du Nord et figure également parmi les espèces les plus utilisées dans les programmes de reboisement au Canada (Nienstaedt et Teich 1972). Un total de 150 millions de plants est mis en terre annuellement au Québec, dont environ 25 millions de plants d’épinette blanche (Beaulieu et al. 2009; Parent 2010). Il s’agit d’une espèce recherchée par l’industrie forestière notamment à cause de la qualité de son bois et de sa croissance supérieure.
Depuis les années 50, l’épinette blanche fait l’objet d’un programme d’amélioration génétique visant l’amélioration du rendement des plantations et de la qualité du bois (Beaulieu et al. 2009). Au cours des années 80, des vergers à graines de première génération ont été implantés. Ces vergers étaient principalement composés d’arbres plus ayant été sélectionnés à partir de leur phénotype en forêt naturelle sur une base régionale (Anonyme 2000). Les responsables de l’amélioration génétique ont ensuite testé et recommandé des sources de semences afin d’implanter une nouvelle génération de vergers à graines pour l’épinette blanche en 1996 (Anonyme 2000). Actuellement, le réseau de vergers à graines de l’épinette blanche est composé de 21 vergers de première génération et de 3 vergers de deuxième génération (André Deshaies, DGPSP, communication personnelle, 2012). En 2010, 98 % des semences d’épinette blanche ensemencées provenaient de sources améliorées génétiquement (André Deshaies, DGPSP, communication personnelle, 2011).
Ces arbres supérieurs produits par les programmes d’amélioration génétique sont la source de semences qui seront utilisées pour établir les plantations futures de haute productivité adaptées aux conditions climatiques actuelles et futures. Or, les changements climatiques sont un phénomène maintenant bien reconnu, dont une des principales origines est l’émission, notamment de dioxyde de carbone (CO2) (GIEC 2007). Ainsi, d’ici 2100, selon
les prévisions du scénario d’émission A1B du groupe d’experts intergouvernemental sur l’évolution du climat (GIEC), la concentration en CO2 atmosphérique pourrait doubler à
2 scénario d’émission se base sur une croissance économique très rapide, une population qui atteint un sommet au milieu du présent siècle, l’adoption rapide de nouvelles technologies plus efficaces, de même qu’une évolution technologique vers l’équilibre entre les sources d’énergie fossile et non fossile. Les changements climatiques envisagés pourraient avoir des répercussions importantes sur les caractéristiques morpho-physiologiques des arbres, comme les caractéristiques de croissance (Campagna et Margolis 1989; Groninger et al. 1996; Hou et al. 2011), les échanges gazeux (Ainsworth et Long 2005; Hou et al. 2011; Wertin et al. 2010) et les caractéristiques foliaires (Cape et Percy 1993; Hou et al. 2011).
Ce projet porte sur les réponses morpho-physiologiques de familles biparentales d’épinette blanche, ainsi que sur leur variabilité et héritabilité familiales suite à l’augmentation seule et simultanée de la température et de la concentration en CO2. Les plants, issus de
croisements dirigés entre les meilleurs parents, ont été produits en serre à la pépinière forestière gouvernementale de Saint-Modeste. Ils ont ensuite été cultivés en conditions contrôlées durant leur deuxième et leur troisième saison de croissance. Les variables de croissance (hauteur, diamètre, masse sèche des parties aériennes et des racines) et les concentrations en éléments minéraux (N, P et K) ont été évaluées à la fin de la deuxième et de la troisième saison de croissance. Des mesures d’échanges gazeux, de transpiration cuticulaire, de quantité de cire cuticulaire, ainsi que de la surface foliaire spécifique ont également été effectuées sur les plants âgés de trois ans.
Une revue de littérature portant sur les changements climatiques, ainsi que sur leurs effets sur les caractéristiques morpho-physiologiques et la variabilité génétique des arbres constitue le premier chapitre de ce mémoire. Le deuxième chapitre présente l’étude des effets d’une augmentation seule et simultanée de la température et de la concentration en CO2 sur les caractéristiques morpho-physiologiques et la variabilité familiale de 20 à 60
familles biparentales d’épinette blanche. Finalement, le dernier chapitre est constitué de la conclusion générale, résumant les principaux résultats obtenus de l’étude et présentant la pertinence et les limites de cette étude, ainsi que des suggestions pour des recherches futures.
3
Objectifs et hypothèses de recherche
Objectifs
1) Évaluer les réponses morpho-physiologiques (croissance, nutrition minérale, échanges gazeux et caractéristiques foliaires) de plants de 20 familles biparentales d’épinette blanche en réponse à l’augmentation seule et simultanée de la température et de la concentration en CO2 durant leur deuxième (2+0) et troisième
(3+0) saison de croissance.
2) Estimer l’effet des changements climatiques sur l’amplitude de la variation génétique et l’héritabilité familiale des variables morpho-physiologiques de plants de 60 familles biparentales d’épinette blanche.
Hypothèses
Les familles d’épinette blanche ne réagissent pas de la même façon, en termes de morpho-physiologie des plants et des aiguilles, à une augmentation de la température et/ou de la concentration en CO2.
Les changements climatiques modifieront de façon significative la variance et l’héritabilité familiale des caractéristiques morpho-physiologiques des plants (3+0) d’épinette blanche.
4
Références
Ainsworth, E.A. et Long, S.P. 2005. Tansley review. What have we learned from 15 years of free-air CO2 enrichment (FACE)? A meta-analytic review of the responses of
photosynthesis, canopy properties and plant production to rising CO2. New
Phytologist 165 : 351-372.
Anonyme. 2000. L'amélioration génétique des arbres au Québec. Gouvernement du Québec - Ministère des Ressources naturelles et de la Faune et Ressources naturelles Canada - Service canadien des forêts. p. 21.
Beaulieu, J., Daoust, G., Deshaies, A., Lamhamedi, M.S., Rainville, A., et Tourigny, M. 2009. Amélioration génétique des arbres, gestion des vergers à graines et de semences, et production de plants forestiers. Dans Manuel de foresterie. O.I.F.Q. Les Presses de l'Université Laval, Québec. p. 1095-1146.
Campagna, M.A., et Margolis, H.A. 1989. Influence of short-term atmospheric CO2
enrichment on growth, allocation patterns, and chemistry of black spruce seedlings at different stages of development. Canadian Journal of Forest Research 19 : 773-782. Cape, J.N., et Percy, K.E. 1993. Environmental influences on the development of spruce
needle cuticles. New Phytologist 125 : 787-799.
GIEC. 2007. Bilan 2007 des changements climatiques. Contribution des Groupes de travail I, II et III au quatrième Rapport d'évaluation du Groupe d'experts intergouvernemental sur l'évolution du climat. 103 p.
Groninger, J.W., Seiler, J.R., Zedaker, S.M., et Berrang, P.C. 1996. Effects of CO2
concentration and water availability on growth and gas exchange in greenhouse-grown miniature stands of loblolly pine and red maple. Functional Ecology 10 : 708-716.
Hou, Y., Qu, J., Luo, Z., Zhang, C., et Wang, K. 2011. Morphological mechanism of growth response in treeline species Minjiang fir to elevated CO2 and temperature.
Silva Fennica 45(2) : 181-195.
Nienstaedt, H., et Teich, A. 1972. The genetics of white spruce. Dans USDA Forest Service Research Paper, WO - 15. U.S.D.A. 24 p.
Parent, B. 2010. Ressources et industries forestières - Portrait statistique juin 2009. Gouvernement du Québec - Ministère des Ressources naturelles et de la Faune - Direction du développement de l'industrie des produits forestiers. p. 498.
Wertin, T.M., McGuire, M.A., et Teskey, R.O. 2010. The influence of elevated temperature, elevated atmospheric CO2 concentration and water stress on net
photosynthesis of loblolly pine (Pinus taeda L.) at northern, central and southern sites in its native range. Global Change Biology 16 : 2089-2103.
5
CHAPITRE 1 : REVUE DE LITTÉRATURE
1.1. Changements climatiques
Le Groupe d’experts intergouvernemental sur l’évolution du climat (GIEC) a produit plusieurs rapports depuis son établissement en 1988 par l’Organisation météorologique mondiale (OMM) et le Programme des Nations Unies pour l’Environnement (PNUE). Ces rapports sont devenus des ouvrages de référence autant pour les décideurs, les experts, les scientifiques que les étudiants chercheurs. Le Groupe d’experts intergouvernemental sur l’évolution du climat a publié en 2007 son dernier rapport de synthèse faisant état des changements climatiques dans le monde. Les prochaines sections sur les changements climatiques observés et envisagés se basent en grande partie sur ce document afin d’en faire ressortir les points importants sur l’évolution du climat, ensuite sont présentées quelques stratégies et politiques sur les changements climatiques.
1.1.1. Changements climatiques observés
Entre 1970 et 2004, les émissions mondiales de gaz à effet de serre (GES) associées aux activités humaines ont augmenté de 70 % (GIEC 2007). L’essentiel de l’augmentation de la température depuis 1950 environ est attribuable aux émissions de GES anthropiques. Par exemple, le secteur forestier, incluant le déboisement, a été responsable de 17,4 % des émissions totales de GES pour l’année 2004 (GIEC 2007). Le CO2 (dioxyde de carbone), le
CH4 (méthane) ainsi que le NO2 (oxyde nitreux), sont des GES importants dont les
concentrations dans l’atmosphère ont fortement augmenté depuis 1750. En effet, en 2005, la concentration en CO2 atmosphérique s’élevait à 379 ppm (partie par million), ce qui est
considérablement plus élevé que ce qui a été observé au cours des 650 000 dernières années. Les observations sur le climat au cours des cinq derniers siècles confirment le réchauffement du système climatique, la seconde moitié du XXe siècle ayant été la plus chaude de cette période en hémisphère Nord. Entre 1906 et 2005, la surface du globe s’est réchauffée en moyenne de 0,74 [0,56-0,92] oC, avec une augmentation plus importante aux latitudes élevées de l’hémisphère Nord. Certains phénomènes ont eu tendance à augmenter
6 durant la période de 1900 à 2005, par exemple les précipitations dans l’est de l’Amérique du Nord. De plus, les fortes précipitations se sont accrues, de même que les vagues de chaleur au niveau des terres émergées. On note également que depuis 50 ans, le nombre de journées et de nuits chaudes a augmenté.
1.1.2. Changements climatiques envisagés
Ces changements climatiques auront de nombreuses répercussions sur le milieu naturel, particulièrement aux latitudes élevées de l’hémisphère Nord (GIEC 2007). Notamment, en forêt boréale, le réchauffement pourrait accroître le risque de modification des aires de répartition des espèces, ainsi que changer la tendance de la biosphère de servir comme puits net important (GIEC 2007). Quelques-uns des changements qui pourraient également être observés dans le climat futur sont l’augmentation de la fréquence des températures très élevées, des vagues de chaleur, des précipitations aux latitudes élevées ainsi que des épisodes de fortes précipitations (GIEC 2007; Rahmstorf et Coumou 2011).
Différents scénarios d’émissions de GES sans politiques climatiques additionnelles ont été élaborés pour la période de 2000 à 2100 (Figure 1.1). Ces scénarios ont été établis dans le Special Report on Emissions Scenarios (SRES) en tenant compte des changements démographiques, technologiques et économiques (GIEC 2000). Notamment, le scénario A1B se base sur une croissance économique très rapide, une population qui atteint un sommet au milieu du présent siècle, ainsi que l’adoption rapide de nouvelles technologies plus efficaces. Ce scénario considère également une évolution technologique vers l’équilibre entre les sources d’énergie fossile et non fossile, la concentration de GES en équivalent CO2 étant estimée à 850 ppm en 2100 (GIEC 2007).
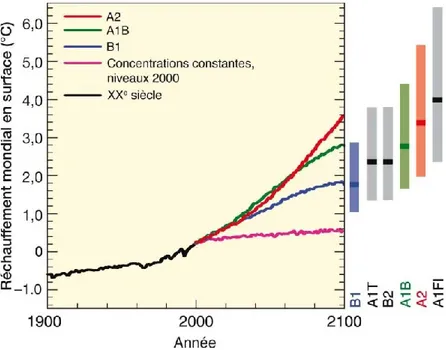
Ces mêmes scénarios ont été considérés pour modéliser les variations possibles des températures (Figure 1.2). Toujours selon le scénario A1B, l’augmentation la plus probable de température d’ici 2090-2099 est de 2,8 °C, avec un intervalle de 1,7 à 4,4 °C, par rapport à 1980-1999 (Tableau 1.1).
7 Figure 1.1. Scénarios d'émissions (lignes pleines) de GES pour la période 2000-2100 en l'absence de
politiques climatiques additionnelles. La zone en gris représente l’intervalle au 80e percentile des scénarios publiés (fourchette post-SRES) et les lignes en pointillés délimitent la plage complète des scénarios (GIEC 2007).
Figure 1.2. Projections relatives aux températures à la surface du globe pour la période 2000-2100, la portion foncée des barres à droite du graphique représentant la valeur la plus probable et celle plus pâle précisant l’intervalle probable. (GIEC 2007).
8 Tableau 1.1. Projections des valeurs moyennes du réchauffement en surface à l’échelle du globe pour la période 2090-2099. (GIEC 2007).
9
1.1.3. Stratégies et politiques
Représentant un tournant dans l'émergence de la conscience environnementale à l’échelle planétaire, la conférence de Stockholm, qui a eu lieu en 1972, a été la première conférence des Nations Unies sur l’environnement humain. Des délégués de 113 pays de même que plusieurs grandes institutions internationales, par exemple la FAO (Food and Agriculture Organization), l’UNESCO (United Nations Educational, Scientific and Cultural Organization) et la Banque mondiale, ont été réunis. Quelques 109 recommandations ont été élaborées par les conférenciers, notamment des recommandations environnementales, sociales et économiques dont l’objectif est d’aider les décideurs à tenir compte de l'environnement lors de l’élaboration des stratégies et des politiques reliées au développement socio-économique. Vingt ans plus tard a eu lieu la conférence des Nations Unies sur l’environnement et le développement à Rio de Janeiro. Plusieurs documents ont été produits. Ainsi, la Déclaration de Rio sur l’environnement et le développement définit, entre autres, la notion de développement durable. La Convention-cadre des Nations Unies sur les changements climatiques (CCNUCC) a également été produite, mais ce n’est qu’en 1994, après la ratification par 50 pays, qu’elle a officiellement été adoptée. Plusieurs principes y ont été élaborés, par exemple celui d’équité intergénérationnelle dont l’objectif était la stabilisation des émissions en 2000 au niveau de 1990.
Depuis cette adoption, une Conférence des Parties (CdP) a lieu annuellement. Cela permet de vérifier l’application des objectifs de la CCNUCC par tous les États en faisant partie. L’analyse des résultats du deuxième rapport du GIEC au cours de la première CdP en 1995 a permis de conclure que la stabilisation des émissions de GES au niveau de 1990 dès 2000 ne serait pas suffisant. Lors de la troisième CdP, en 1997, certaines parties ont donc signé le Protocole de Kyoto, à l’intérieur duquel des objectifs juridiquement contraignants de réduction des émissions ont été négociés. Ce n’est toutefois qu’en février 2005 qu’il est entré en vigueur, à la suite de la signature de la Russie, permettant ainsi d’obtenir la signature de plus de 55 pays producteurs de plus de 55 % des émissions de GES. Pour la période d’engagement de 2008 à 2012, un des objectifs d’émissions chiffrés contraignants visait la réduction des émissions globales d’au moins 5 % par rapport aux niveaux de 1990. La deuxième phase du protocole couvrira la période de 2013 à 2017.
10 La Canada a signé et ratifié le protocole de Kyoto en 1997 pour la première phase de 2008 à 2012. Toutefois, en décembre 2011, le pays s’est retiré du protocole. Selon le ministre de l'Environnement, Peter Kent, l’objectif de réduction des émissions de GES de 6 % par rapport à 1990 était trop ambitieux. Le Canada n’aurait pas été capable de respecter ses engagements à l’échéance de la première phase. Aucun engagement ne sera pris pour la deuxième phase du protocole. Le Canada dispose tout de même d’un plan d’action sur les changements climatiques visant un objectif de réduction, d’ici 2020, de 17 % du niveau des émissions de 2005 correspondant à 740 Mt en équivalent CO2 (Anonyme 2012) (Figure
1.3). D’ailleurs, une partie de l’ambitieux objectif de réduction des émissions a été atteint en 2010, les émissions de GES ayant diminué à 692 Mt d’équivalent CO2 (Figure 1.3). Des
mesures ont ainsi déjà été mises en place, telles que l’institution des règlements sur les émissions de gaz à effet de serre dans les secteurs dont les émissions sont les plus élevées, c’est-à-dire les transports, le pétrole et gaz et l’électricité (Figure 1.4). Le secteur des transports étant la source d’émissions de GES la plus importante, depuis le 15 décembre 2010, il doit y avoir en moyenne 5 % de carburant renouvelable dans l'essence et 2 % dans le diesel. Dans le cadre de l'Accord de Copenhague, le Canada a de plus versé 400 millions de dollars, au cours de 2010-2011, pour le soutien des pays en voie de développement dans le but de réduire leurs émissions de GES ainsi que de s’adapter aux effets des changements climatiques. Pour les exercices financiers 2011-2012 et 2012-2013, 800 millions de dollars seront attribués par le Canada au soutien des initiatives internationales de lutte contre les changements climatiques (Anonyme 2012).
11 Figure 1.3. Émissions totales de GES au Canada, en mégatonnes (Mt) d’équivalent de CO2 pour 1990 à
2010. Adapté de http://www.ec.gc.ca/ges-ghg/default.asp?lang=Fr&n=8BAF9C6D-1. .
Figure 1.4. Émissions totales de GES au Canada, par secteur économique, en mégatonnes (Mt) d’équivalent de CO2 pour 2010. Adapté de
http://www.ec.gc.ca/ges-ghg/default.asp?lang=Fr&n=AC8F85A5#s32. . Bâtiments 79 Agriculture 69 Déchets et autres 50 Électricité 99 Industries dépendantes du commerce produisant des émissions élevées 75 Transports 166 Pétrole et gaz 154 589 718 740 726 751 731 690 692 550 600 650 700 750 800 1990 2000 2005 2006 2007 2008 2009 2010
12 Les mesures de réduction des émissions de GES par le gouvernement du Canada doivent être complétées par des efforts des gouvernements provinciaux. À cet égard, le Québec compte parmi les provinces engagées dans la lutte aux changements climatiques. Il est d’ailleurs un chef de file en matière de lutte, notamment à cause de sa production d’énergie propre, soit l’hydroélectricité (MDDEP 2012). Avec sa Stratégie gouvernementale d’adaptation aux changements climatiques et son Plan d’action 2013-2020 sur les changements climatiques (PACC 2013-2020), qui sont en finalisation, le Québec vise la réduction des émissions de GES ainsi que l’adaptation aux changements climatiques pour les prochaines années. L’objectif de réduction des émissions est de 20 % sous le niveau de 1990 d’ici 2020 (MDDEP 2012). Les avancées du Québec réalisées en termes de développement durable et de réduction des émissions de GES depuis les dernières années sont dues, entre autres, au Plan d’action 2006-2012 sur les changements climatiques (PAAC). De plus, lors de la onzième CdP, qui a eu lieu à Montréal en 2005, le Québec et le Manitoba ont tenu le troisième Sommet des leaders sur les changements climatiques. À la suite de cette rencontre, un document visant l’engagement pour la lutte aux changements climatiques ainsi que la collaboration des États fédérés de plusieurs régions du monde pour la construction de la future économie verte a été produit. Il s’agit de la Déclaration commune sur les changements climatiques. Faisant partie de multiples organisations et partenariats, à titre de participant ou d’observateur, le Québec se voit parmi les leaders en Amérique du nord pour la lutte aux changements climatiques (MDDEP 2012).
13
1.2. Effets des changements climatiques sur les caractéristiques
morpho-physiologiques des arbres
L’augmentation de la température et de la concentration en CO2, associée aux changements
climatiques, pourrait avoir des répercussions importantes sur la croissance et le développement des arbres (Bazzaz 1990). Plusieurs études ont été réalisées sur le sujet. Cette section présente, notamment, les résultats de certaines études ayant évalué les effets d’une augmentation seule ou simultanée de la concentration en CO2 et de la température sur
les caractéristiques de croissance (hauteur, diamètre et masses sèches), les échanges gazeux (photosynthèse et conductance stomatique) et les caractéristiques foliaires (surface foliaire spécifique).
1.2.1. Rappel sur la photosynthèse
La photosynthèse au niveau de la feuille est généralement modélisée à partir du modèle de Farquhar, c’est-à-dire le premier modèle mathématique de la photosynthèse ayant une base mécanistique en terme de biochimie du processus (Farquhar et al. 1980). Le modèle de Farquhar permet de modéliser les différentes étapes de la phase lumineuse et de la phase sombre. Il tient compte que la photosynthèse est limitée par le plus lent des trois processus, soit Je, Jc et Js, chacun étant décrit comme des expressions différentes du taux d’absorption du CO2. Le processus biochimique de la photosynthèse peut ainsi être limité par la lumière
et ses effets sur le transport d’électron contrôlant la régénération de la RuBisCO (Je), par le taux de carboxylation de la RuBisCO (Jc), ou encore par la demande en produits de la
photosynthèse, tel que le sucrose, utilisé au niveau du métabolisme cellulaire (Js) (Collatz
et al. 1991). La photosynthèse a été modélisée de cette façon afin de comprendre le processus autant au niveau du chloroplaste que de l’écosystème et même des grilles de cellules (100 x 100 km) des modèles de circulation générale, décrivant les interactions entre la surface de la terre et l’atmosphère, le climat, ainsi que la météorologie (Sellers et al. 1986, 1992). Selon Farquhar et al. (1980), la façon dont ces deux processus varient, en
14 valeur absolue ou relative, pourrait bien être la clé de la compréhension de l’écophysiologie des espèces végétales.
Avant d’aborder les effets des changements climatiques sur les caractéristiques morpho-physiologiques, voici un rappel du fonctionnement de la photosynthèse sous le climat actuel (Figure 1.5). Ce processus se résume selon la formule suivante :
6CO2 + 12H2O + Énergie lumineuse -> C6H12O6 + 6O2 + 6H20
La photosynthèse a lieu à l’intérieur des chloroplastes, dans les cellules chlorophylliennes, et se divise en deux phases. Dans les thylakoïdes, la phase lumineuse produit de l’ATP et du NADPH + H+. Ces derniers sont utilisés lors de la phase de fixation du carbone,
c’est-à-dire le cycle de Calvin, divisé en trois étapes. La première étape est l’incorporation du CO2,
catalysée par la RuBisCO, la protéine la plus abondante de la biosphère. Cette réaction donne deux molécules d’AGP (acide phosphoglycérique) qui sont utilisées dans la deuxième étape, soit la réduction d’AGP en glucides à 3 atomes de carbone (aldéhyde phosphoglycérique). Ces glucides sont nécessaires à la troisième et dernière étape du cylce de Calvin, la régénération de la RuBisCO et servent également à la croissance.
La RuBisCO a une fonction carboxylase, soit la fixation du CO2, et une fonction
oxygénase, soit la fixation de l’O2, appelée photorespiration. À la concentration en CO2
ambiante, à la lumière saturante et à la température optimale, la photosynthèse des espèces d’arbre est toujours limitée par la RuBisCO (Ainsworth et Rogers 2007, Ghannoum et al. 2010; Sage et Kubien 2007).
15 Figure 1.5. Processus de la photosynthèse, a) à l’intérieur du chloroplaste et b) détail des étapes du Cycle de Calvin (adapté de http://www.snv.jussieu.fr).
16
1.2.2. CO2
1.2.2.1. Enrichissement en CO2
Afin d’étudier et d’évaluer les conséquences possibles d’une augmentation de la concentration en CO2 dans le climat futur sur les espèces végétales, l’enrichissement
artificiel en CO2 de l’air peut s’avérer très utile afin de simuler les conditions futures
prédites. Plusieurs techniques d’enrichissement en CO2 de l’air environnant des espèces de
plantes ou d’arbres sont utilisées entre autres en recherche. Ces techniques peuvent différer notamment selon la dimension des végétaux, la durée de l’enrichissement en CO2 ou encore
selon le nombre de facteurs à considérer simultanément. Quatre de ces techniques sont présentées ci-dessous.
Les chambres de croissance sont une de ces techniques (Figure 1.6). L’environnement contrôlé y permet la modification simultanée de plusieurs facteurs, par exemple la concentration en CO2, la température et l’humidité relative de l’air (Ceulemans et
Mousseau 1994). Par contre, un des facteurs limitant à l’intérieur des chambres de croissance est la luminosité relativement peu élevée, même avec l’ajout de lumières supplémentaires (Ceulemans et Mousseau 1994). Cette technique a souvent été utilisée pour des expériences à court terme utilisant des végétaux de petites dimensions, par exemple des plantes ou encore des plants d’arbres.
Pour les expériences à plus long terme d’enrichissement en CO2, les chambres à ciel ouvert
(OTP - open top chambers) sont couramment utilisées (Figure 1.7) (Ceulemans et Mousseau 1994). Elles permettent l’étude de la canopée des plantes et de petits arbres. Par contre, cette technique nécessite une grande quantité de CO2. De plus, elle ne permet pas
d’évaluer l’interaction avec d’autres paramètres, puisque les facteurs environnementaux naturels à l’extérieur, tels que la température, l’humidité relative ainsi que le vent, ne peuvent pas être contrôlés.
Les deux techniques précédentes sont limitées par leur taille et ne sont pas adéquates pour des arbres matures ou des forêts. Une alternative pour les arbres de plus grande dimension
17 est donc de traiter les branches individuellement, à l’aide d’enceintes à branche (branch bags) (Figure 1.8), assumant toutefois l’autonomie des branches (Ceulemans et Mousseau 1994). L’enceinte agit comme un système ouvert, l’air autour des branches étant enrichi continuellement en CO2 et, à intervalles réguliers, elle se ferme pour permettre
l’enregistrement des données (Saugier 1999). L’enregistrement de la diminution de la concentration en CO2 ainsi que de l’augmentation de la quantité de vapeur d’eau durant
quelques minutes permet de mesurer la photosynthèse de même que la transpiration de ces branches. Même si les résultats associés aux études ayant utilisé des enceintes à branche ne peuvent être directement utilisés pour représenter un arbre en entier, ils peuvent aider à clarifier les réponses des espèces végétales dans un environnement enrichi en CO2
(Roberntz et Stockfors 1998).
Finalement, le système FACE (free air CO2 enrichment) permet l’enrichissement en CO2 de
l’air autour des arbres de grande dimension, tout en conservant les conditions naturelles de croissance (Figure 1.9) (Ashton et al. 2012). Il s’agit d’une des seules techniques pour la mesure de certains des effets des changements climatiques sur un écosystème forestier en entier (Ceulemans et Mousseau 1994). Par contre, ce système nécessite un investissement et des coûts d’entretien importants et ne permet pas de contrôler les facteurs comme la température ou l’humidité de l’air. Par ailleurs, une étude sur deux espèces d’arbres tropicaux en C3 a démontré que le système FACE pouvait sous-estimer les effets potentiels
d’une concentration élevée en CO2 sur les caractéristiques morpho-physiologiques des
espèces végétales (Holtum et Winter 2003). La fluctuation continue de la concentration en CO2 à laquelle sont exposées les espèces végétales, causée par des ajustements constants
18 Figure 1.6. Chambre de croissance. Figure 1.7. Chambres à ciel ouvert (OTC).
http://www.wsl.ch/ozone/ICP-Forests_2001.ehtml.
Figure 1.8. Enceinte à branches (branch bags), pour la mesure d’échanges gazeux. (Saugier 1999)
Figure 1.9. Système FACE.
19 1.2.2.2. Effets de la concentration en CO2 sur les caractéristiques morpho-physiologiques
En théorie, les végétaux répondent à l'élévation de la concentration en CO2 atmosphérique,
notamment, en augmentant leur photosynthèse (Chaturvedi et al. 2009; Medlyn et al. 1999; Sholtis et al. 2004), généralement de façon non linéaire (Luo et al. 1998), ainsi qu’en diminuant leur conductance stomatique, leur transpiration (Ainsworth et al. 2003; Ainsworth et Rogers 2007; Chaturvedi et al. 2009; Saxe et al. 1998) et possiblement leur taux de respiration (Gonzalez-Meler et al. 2004). À court terme, une augmentation de la concentration en CO2 stimule la fonction carboxylase de la RuBisCO (Stitt 1991)
davantage que sa régénération (Ghannoum et al. 2010). Ainsi, même si une augmentation de la photosynthèse nette est observable (Ceulemans et Mousseau 1994; Eamus et Jarvis 1989), la RuBisCO reste toujours limitante chez les espèces d’arbres (Ainsworth et Rogers 2007). L’augmentation de la photosynthèse peut se caractériser par une augmentation de la croissance (Kirschbaum 2011). La réponse de la croissance à l’augmentation de la photosynthèse a tendance à être supérieure sous des conditions optimales, notamment lorsque l’accès aux nutriments est adéquat (Kirschbaum 2011). Toutefois, en pratique, ce n'est pas toujours le cas, puisqu'il est possible que l'arbre ne soit pas capable d'utiliser ou d'entreposer ces glucides supplémentaires (Stitt 1991). Entre autres, il est possible qu’une espèce soit limitée par des contraintes génétiques, sa croissance pourra répondre à une augmentation de la disponibilité des ressources jusqu’à la limite fixée par ces contraintes génétiques (Kirschbaum 2011).
À long terme, c’est-à-dire quelques semaines à plusieurs mois (Jach et Ceulemans 2000; Sage et al. 1989), l'exposition à des concentrations élevées en CO2 aurait tendance à inhiber
la photosynthèse (Chaturvedi et al. 2009) à cause de l’acclimatation à ces conditions (Ceulemans et Mousseau 1994, Stitt 1991). Selon Sage (1994), l’acclimatation se définit comme étant les réponses morpho-physiologiques qui améliorent la performance et la survie d’un individu (espèce végétale) par l’augmentation de la croissance, l’efficacité d’utilisation des ressources, l’efficacité de la reproduction, la tolérance aux stress et/ou la durée de vie sous des conditions environnementales différentes. L’acclimatation de la photosynthèse peut s’expliquer par le fait que les enzymes présents dans le cycle de Calvin, en particulier la RuBisCO, ont tendance à diminuer à la suite de l’accumulation d’hydrates
20 de carbone (Makino et Mae 1999; Stitt 1991), pouvant se traduire par la diminution de l’efficience et de la capacité de régénération de la RuBisCO (Jach et Ceulemans 2000). Il a tout de même été observé qu’une exposition à long terme (quatre ans) à des concentrations élevées en CO2 s’est traduite par une augmentation de la biomasse racinaire et aérienne
chez Populus deltoides (Barron-Gafford et al. 2005).
La réponse de la photosynthèse à l’augmentation de la concentration en CO2 varie souvent
avec d’autres variables physiologiques et environnementales (Booker et Maier 2001; Cao et al. 2007; Kirschbaum 2011; Sholtis et al. 2004). Par exemple, une étude sur Arachis hypogaea L. a mis en évidence que la photosynthèse avait tendance à être inhibée par l’éthylène (Pallas et Kays 1982). Dans un contexte de changements climatiques, l’azote (N) pourrait plus particulièrement jouer un rôle critique quant à la régulation de la réponse de la photosynthèse des espèces végétales (Bauer et Berntson 2001). En effet, l’azote est un composant majeur de la RuBisCO, ainsi que d’autres enzymes et structures associées à la photosynthèse. Par ailleurs, il s’agit d’un élément limitant au plan de la productivité de la plupart des écosystèmes terrestres en zone tempérée. Plusieurs études ont déjà observé un effet positif de l’ajout d’azote sur le plan de la stimulation de la photosynthèse et de la croissance par une concentration élevée en CO2 (Cao et al. 2007; Roberntz et Stockfors
1998; Wang et al. 1994). Ainsi, l’augmentation de la disponibilité en N pourrait augmenter le potentiel des espèces végétales à répondre positivement en terme de croissance à l’augmentation de la concentration en CO2 (Bazzaz 1990; Luo et al. 2006). Une faible
concentration en azote foliaire limite la photosynthèse (Crous et al. 2008; Makino et Mae 1999), augmentant linéairement avec l’augmentation de la concentration en azote foliaire, alors qu’une trop grande assimilation d’azote peut être toxique et causer une diminution du taux de photosynthèse (Landis 1989). Les résultats d’une étude sur le bouleau blanc (Betula papyrifera Marsh.) ont ainsi montré une augmentation du taux de photosynthèse avec la concentration en azote foliaire jusqu’à un niveau optimal puis une diminution du taux de photosynthèse, sans différence significative entre les traitements de CO2 ambiant et CO2
élevé (Cao et al. 2007).
La distribution de l’azote dans les arbres est fonction notamment des tissus, de l’âge, du stade de développement et de la saison. Les plus fortes concentrations d’azote se retrouvent
21 dans les tissus comportant des cellules physiologiquement actives. Les arbres au stade juvénile ont donc des puits très forts. L’absorption de l’azote est intense dans les phases initiales de croissance rapide. Par ailleurs, une étude de Ainsworth et al. (2003) sur une plante en C3 a montré qu’à la suite de la récolte des parties aériennes, les repousses
permettaient d’avoir des puits très forts dans la plante. Cela permettrait notamment de supprimer l’acclimatation de la photosynthèse à une augmentation de la concentration en CO2, puisqu’elle est fonction de la force des puits (Ainsworth et al. 2003; Makino et Mae
1999; Roberntz et Stockfors 1998). Plus particulièrement chez les espèces ligneuses, la concentration foliaire en éléments minéraux (azote, potassium, phosphore, magnésium, etc.) a tendance à diminuer à la suite de leur exposition à des concentrations élevées en CO2
(Medlyn et al. 1999; Roberntz et Stockfors 1998; Sholtis et al. 2004; Wullschleger et al. 1997). Pour expliquer ce phénomène, deux explications sont possibles : 1) un effet de dilution causé par une augmentation de la quantité de glucides produits, et donc de la croissance (Booker et Maier 2001; Roberntz et Stockfors 1998), ou 2) un mécanisme de rétroaction qui induit une translocation des nutriments vers d’autres parties de la branche ou de l’arbre (Roberntz et Stockfors 1998).
La stratégie des plantes en C3 et en C4 est différente notamment au plan de la
photosynthèse. Par unité d’azote, elle est plus élevée chez les plantes en C4. Chez ce type
de plante, tout comme les plantes en C3, l’augmentation de la concentration en CO2
s’accompagne d’une diminution de la conductance stomatique et de la transpiration. Toutefois, contrairement aux plantes en C3 elles maintiennent leur photosynthèse stable
(Adam et al. 2000). Cela se traduit par une meilleure efficacité d’utilisation de l’eau pour les espèces en C4, un avantage considérant que la fréquence des épisodes de sécheresse
devrait augmenter dans le climat futur prédit (GIEC 2007).
Il est possible de trouver dans la littérature plusieurs études portant sur les effets d’une augmentation de la concentration en CO2 sur les caractéristiques morpho-physiologiques
des espèces ligneuses. Ces études fournissent une multitude d’observations qui peuvent être utiles notamment pour anticiper les effets bénéfiques potentiels résultant d’une augmentation de la photosynthèse (Kirschbaum 2011). Cependant, les espèces forestières ne réagissent pas de façon similaire à l’augmentation du CO2 en matière de croissance et de
22 réponses physiologiques (Bazzaz et al.1990; Hou et al. 2011). Le tableau 1.2 résume quelques-unes de ces études mettant l’accent principalement sur les essences forestières. Ainsi, la majorité des études présentées ont observé qu’une augmentation de la concentration en CO2 entre 360 et 660 ppm se traduit soit par un effet non significatif ou un
effet positif (de 9 à 303 %) sur les caractéristiques morpho-physiologiques des espèces ligneuses. Pour toutes les études, des plants de moins de deux ans ont été utilisés, l’enrichissement en CO2 ayant été effectué soit en serre ou soit en chambre de croissance,
durant quelques semaines jusqu’à deux saisons de croissance. En pratique toutefois, en production de plants, il n’est généralement pas possible d’enrichir l’air en CO2 dans le but
d’obtenir une croissance supérieure. Par ailleurs, lorsque les plants sont cultivés en serres, il a été démontré que la concentration en CO2 a tendance à diminuer comparativement à la
concentration de l’air à l’extérieur. Ainsi, afin de conserver un taux de photosynthèse au moins équivalent à celui des plants cultivés à l’extérieur, dans des conditions pratiques, il a été recommandé par Thongbai et al. (2011) d’apporter une bonne circulation de l’air pour avoir une concentration en CO2 à l’intérieur des serres la plus élevée possible.
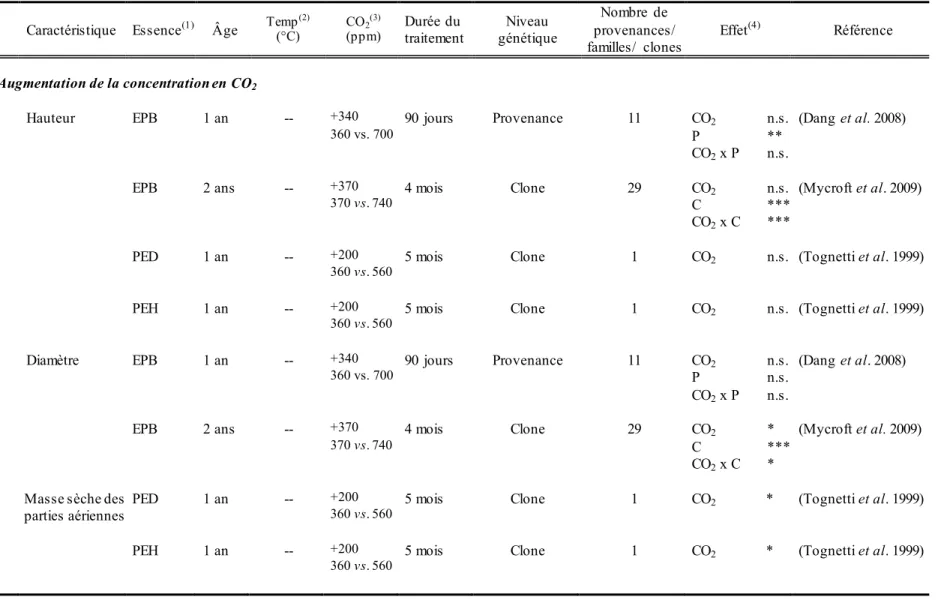
23 Tableau 1.2. Synthèse de quelques travaux sur les effets d’une augmentation de la concentration en CO2 sur les caractéristiques de croissance, les échanges
gazeux et les caractéristiques foliaires de jeunes plants d’essences résineuses.
Caractéristique Nom de l’espèce CO2 (ppm)(1) Conditions de croissance début du traitement Âge des plants au traitementDurée du (2) Effet(3) Référence Hauteur Picea mariana Mill.
(Épinette noire) +660 (340 vs. 1000) Serre 34 jours 78 jours 120 jours 207 jours 3 sem 6 sem 3 sem 6 sem 3 sem 6 sem 3 sem 6 sem n.s. +15 n.s. +15 n.s. n.s. n.s. n.s. (Campagna et Margolis 1989) +660 (environ 340
vs. 1000) Serre Semences 115 jours + (Lord et al. 1993)
Pinus banksiana Lamb.
(pin gris) +660 (340 vs. 1000) Serre 57 jours 99 jours 2 ans 3 sem 6 sem 3 sem 6 sem 3 sem 6 sem n.s. n.s. n.s. n.s. n.s. n.s. (Campagna 1989)
+360 (360 vs. 720) Serre 1 an 4 mois n.s. (Zhang et Dang 2007)
Pinus taeda L. (pin à
encens) +397 (401 vs. 798) Serre Semences 2 saisons +14 (Groninger et al. 1996) Diamètre Picea glauca (Moench)
Voss (épinette blanche)
+360 (360 vs. 720) Serre 1 an 4 mois + (Zhang et Dang 2007)
Picea mariana Mill.
(épinette noire)
+660 (environ 340
vs. 1000)
Serre Semences 115 jours + (Lord et al. 1993) +360 (360 vs. 720) Serre 1 an 4 mois + (Zhang et Dang 2007)
Pinus banksiana Lamb.
24 Caractéristique Nom de l’espèce CO2 (ppm)(1) Conditions de croissance début du traitement Âge des plants au traitementDurée du (2) Effet(3) Référence
Masse sèche des parties
aériennes (aiguilles) Pinus banksiana Lamb. (pin gris) +660 (340 vs. 1000) Serre 57 jours 99 jours 2 ans 3 sem 6 sem 3 sem 6 sem 3 sem 6 sem n.s. n.s. +13 n.s. +9 n.s. (Campagna 1989)
Masse sèche des parties aériennes (tige)
Pinus banksiana Lamb.
(pin gris) +660 (340 vs. 1000) Serre 57 jours 99 jours 2 ans 3 sem 6 sem 3 sem 6 sem 3 sem 6 sem n.s. n.s. +9 n.s. n.s. n.s. (Campagna 1989)
Masse sèche des racines Picea mariana Mill.
(épinette noire) +660 (environ 340 vs. 1000) Serre Semences 115 jours + (Lord et al. 1993) Pinus banksiana Lamb.
(pin gris) +660 (340 vs. 1000) Serre 57 jours 99 jours 2 ans 3 sem 6 sem 3 sem 6 sem 3 sem 6 sem n.s. n.s. n.s. n.s. n.s. n.s. (Campagna 1989)
Masse sèche totale Picea mariana Mill.
(épinette noire) +660 (340 vs. 1000) Serre 34 jours 78 jours 120 jours 207 jours 3 sem 6 sem 3 sem 6 sem 3 sem 6 sem 3 sem 6 sem + + + + + + n.s. n.s. (Campagna et Margolis 1989) +360 (340 vs. 700) Chambres de croissance
Semences 35 jours +54 (Johnsen 1993) +660 (environ 340
vs. 1000)
25 Caractéristique Nom de l’espèce CO2 (ppm)(1) Conditions de croissance début du traitement Âge des plants au traitementDurée du (2) Effet(3) Référence
Masse sèche totale (suite) Pinus banksiana Lamb. (pin gris) +660 (340 vs. 1000) Serre 57 jours 99 jours 2 ans 3 sem 6 sem 3 sem 6 sem 3 sem 6 sem n.s. n.s. +14 n.s. +8 n.s. (Campagna 1989)
Pinus taeda L. (pin à
encens) +397 (401 vs. 798) Serre Semences 2 saisons +43 (Groninger et al. 1996) Photosynthèse Picea mariana Mill.
(épinette noire)
+360 (340 vs. 700) Chambres de croissance
Semences 35 jours + (Johnsen 1993)
Pinus taeda L. (pin à encens)
+397 (401 vs. 798) Serre Semences 2 saisons +303 (Groninger et al. 1996) Conductance stomatique Pinus taeda L. (pin à
encens)
+397 (401 vs. 798) Serre Semences 2 saisons n.s. (Groninger et al. 1996)
(1) Correspond à l’augmentation (lorsque précédé d’un +) de la concentration en CO
2 et à la concentration en CO2 des traitements (entre parenthèses), en ppm. (2) sem semaine.
(3) + effet positif d’une augmentation de la température et/ou de la concentration en CO
2, - effet négatif d’une augmentation de la température et/ou de la
concentration en CO2, les chiffres qui suivent + ou - correspondent à la valeur, en pourcentage (%), de la variation associée au traitement d’augmentation de la
26
1.2.3. Température
Tout comme la concentration en CO2, l’évaluation des effets de l’augmentation de la
température peut s’avérer très utile afin d’évaluer certains des effets principaux des changements climatiques. En effet, la température est un des pricipaux éléments controlant la distribution et la productivité des espèces végétales, dont les effets sur les processus physiologiques sont importants (Sage et Kubien 2007). Un environnement contrôlé est nécessaire afin de maintenir la température désirée. Ainsi, les chambres de croissance peuvent être utilisées, permettant de contrôler, de modifier et de simuler simultanément plusieurs variables environnementales importantes associées aux changements climatiques.
L’augmentation de la température de l’air observée depuis quelques années et due aux changements climatiques pourrait affecter la structure et le fonctionnement des écosystèmes forestiers, notamment en diminuant la capacité d’assimilation du CO2 atmosphérique
(Gunderson et al. 2010). Selon le degré d’adaptation des espèces, le rythme du réchauffement et le mode de développement socio-économique, le risque d’extinction des espèces pourrait être plus ou moins important, pouvant atteindre jusqu’à 30 %, et même plus (GIEC 2007). Pour être en mesure de prédire les effets du réchauffement climatique, il est impératif de comprendre la façon dont la température affecte la séquestration du carbone via la photosynthèse (Sage et Kubien 2007). La prochaine section fait donc état des connaissances au sujet de la réponse des espèces végétales à l’augmentation de la température.
1.2.3.1. Effets de la température sur les caractéristiques morpho-physiologiques
Une augmentation de la température de l’air peut se traduire par un changement au plan de la réponse morpho-physiologique des espèces végétales. L’augmentation de la température jusqu’à l’atteinte de la température optimale pour un processus en particulier se traduit par un effet positif, alors que l’augmentation au-delà de la température optimale peut se traduire par un effet négatif sur les caractéristiques morpho-physiologiques. Par exemple, le taux de photosynthèse a tendance à augmenter à partir de 0 ºC jusqu’à l’atteinte de la température optimale et diminuer rapidement avec une augmentation subséquente (Hou et