Maîtrise du chiendent commun (Elymus repens) par
l’emploi d’une bâche de plastique opaque
Mémoire
Amélie Picard
Maitrise en biologie végétale
Maître ès sciences (M. Sc.)
Québec, Canada
Maîtrise du chiendent commun (Elymus repens) par
l’emploi d’une bâche de plastique opaque
Mémoire
Amélie Picard
Sous la direction de :
iii
Résumé
Le chiendent commun (Elymus repens), étant une graminée adventice, vivace, envahissante et difficile à réprimer, il importe de trouver une méthode pour le maîtriser à long terme. Cette méthode doit prioriser la destruction des organes souterrains du chiendent, rhizomes et bourgeons des rhizomes, organes qui assurent sa dispersion. L’utilisation d’une bâche de plastique opaque est une méthode qui n’a pas été étudiée chez le chiendent, mais qui connaît du succès auprès d’autres adventices vivaces envahissantes (Lepidium latifolium et
Pueraria montana). C’est une méthode qui peut être utilisée pour différentes durées et à
différents moments dans la saison. Cette étude a permis de montrer que l’application d’une bâche de plastique opaque sur un terrain infesté de chiendent procure une maîtrise efficace de l’adventice. De plus, les résultats indiquent que la bâche peut être installée dès le printemps ou plus tard dans la saison pour obtenir un effet semblable sur les rhizomes du chiendent. Aussi, elle peut être installée pendant 6, 12 ou 18 semaines et avoir le même effet à long terme sur les rhizomes du chiendent. Les bâches ont diminué de 70 % la biomasse sèche des rhizomes ainsi que le nombre de bourgeons sur les rhizomes dans la même année. Cet effet s’est poursuivi dans le temps, un an après le retrait des bâches, où les rhizomes du chiendent étaient encore réduits en moyenne à 70 %.
iv
Table des matières
Résumé ... iii
Table des matières ... iv
Liste des figures ... vi
Liste des tableaux ... vii
Remerciements ... ix 1. Introduction ... 1 1.1 Problématique ... 1 1.2 Revue de littérature ... 2 1.2.1 Biologie du chiendent ... 2 1.2.2 Nuisibilité en agriculture ... 7 1.2.3 Méthodes de lutte ... 9 1.3 Hypothèses de travail ... 15 1.4 Objectifs de la recherche ... 15 2. Matériel et méthodes ... 16 2.1 Sites expérimentaux ... 16 2.2 Dispositifs expérimentaux ... 18
2.2.1 Matériel utilisé et mise en place des traitements ... 18
2.2.2 Variables mesurées ... 22
2.3 Analyses statistiques ... 23
3. Résultats ... 27
3.1 Biomasse sèche des rhizomes de chiendent ... 27
3.2 Décompte de bourgeons sur les rhizomes de chiendent ... 31
4. Discussion ... 40
4.1 Effet des bâches sur la maîtrise du chiendent ... 40
4.2 Faisabilité de la méthode ... 47
v
6. Références ... 51
7. Annexes ... 54
Annexe 1. Fiche technique des caractéristiques de la bâche utilisée ... 54
Annexe 2. Coefficients des comparaisons simples à un degré de liberté ... 62
Annexe 3. Présentation des graphiques des résidus pour la biomasse sèche des rhizomes de chiendent ... 63
Annexe 4. Présentation des graphiques des résidus pour le nombre de bourgeons des rhizomes de chiendent ... 64
Annexe 5. Résultats des comparaisons simples pour le nombre de bourgeons sur les rhizomes du chiendent (valeurs non transformée) à T0 ... 65
Annexe 6. Résultats des comparaisons simples pour le nombre de bourgeons sur les rhizomes du chiendent (valeur non transformée) à T6 ... 66
Annexe 7. Résultats des comparaisons simples pour le nombre de bourgeons sur les rhizomes du chiendent (valeur non transformée) à T12 ... 67
Annexe 8. Nombre moyen de bourgeons sur les rhizomes du chiendent selon le moment d’échantillonnage et le traitement ... 68
vi
Liste des figures
Figure 1. Dispersion du chiendent par les rhizomes ... 4 Figure 2. Croissance des différents biotypes de chiendent commun au Minnesota ... 5 Figure 3. Sites expérimentaux de 2015 (site 1) et 2016 (site 2) à la station Agronomique de Saint-Augustin-de-Desmaures ... 16 Figure 4. Plan détaillé des parcelles et sous-parcelles, leurs dimensions ainsi que des zones d’échantillonnage du sol dans chaque parcelle... 18 Figure 5. Étapes de mise en place de la bâche dans les parcelles des traitements 1 à 7 ... 19 Figure 6. Étapes pour la réalisation des échantillonnages du sol dans les sous parcelles à l’aide des quadrats ... 23 Figure 7. Comparaison simple GLY vs TB illustrée dans le temps pour le nombre de bourgeons sur les rhizomes du chiendent commun ... 37 Figure 8. Pourcentage de maîtrise des rhizomes de chiendent selon la date d'échantillonnage ... 40 Figure 9. Contraste B18 vs autres Bâches illustré dans le temps pour le nombre de bourgeons sur les rhizomes du chiendent commun ... 43 Figure 10. Contraste fauche vs travail de sol illustré dans le temps pour le nombre de bourgeons sur les rhizomes du chiendent commun ... 46
vii
Liste des tableaux
Tableau 1. Protocole expérimental pour 2015 et 2016 avec le numéro des traitements, leurs descriptions et abréviations ainsi que les dates de mise en place de ces traitements ... 21 Tableau 2. Dates des interventions de travail de sol et de fauche aux 2 sites pour les traitements 4 à 7 ... 21 Tableau 3. Conditions lors de l'application du traitement de glyphosate (8) ... 22 Tableau 4. Description des comparaisons simples et de leurs traitements associés ... 25 Tableau 5. ANOVA pour les données de biomasse sèche des rhizomes du chiendent commun selon les traitements disposés selon un plan en blocs complets aléatoire ... 27 Tableau 6. ANOVA pour les données de biomasse sèche des rhizomes du chiendent commun selon les traitements disposés selon un plan en blocs complets aléatoire sans les interactions non significatives ... 28 Tableau 7. Résultat des comparaisons simples pour la biomasse sèche des rhizomes de chiendent (valeurs non transformées) ... 29 Tableau 8. Biomasse sèche des rhizomes en fonction des traitements ... 30 Tableau 9. Biomasse sèche moyenne des rhizomes en fonction des deux sites étudiés de St-Augustin-de-Desmaures ... 30 Tableau 10. Biomasse sèche moyenne des rhizomes en fonction des temps d'échantillonnage ... 31 Tableau 11. ANOVA pour les données de décompte de bourgeons sur les rhizomes du chiendent commun selon les traitements disposés selon un plan en blocs complets aléatoire ... 32 Tableau 12. ANOVA pour les données de décompte de bourgeons sur les rhizomes du chiendent commun selon les traitements disposés selon un plan en blocs complets aléatoire sans les interactions non significatives ... 33 Tableau 13. Interaction Traitement*Temps décomposé par moment d'échantillonnage pour la variable nombre de bourgeons sur les rhizomes ... 33 Tableau 14. Résultat des comparaisons simples à un degré de liberté à 18 semaines après la mise en place des traitements pour le nombre de bourgeons sur les rhizomes de chiendent commun ... 35
viii
Tableau 15. Comparaisons simples à un degré de liberté pour les échantillons pris à 56 semaines après la mise en place des traitements pour le nombre de bourgeons sur les rhizomes du chiendent commun ... 36 Tableau 16. Nombre de bourgeons sur les rhizomes de chiendent ainsi que les pourcentages de la population initiale selon les traitements et le moment de l’échantillonnage ... 38 Tableau 17. Degrés jours cumulés sous les bâches et dans un traitement témoin sans bâche ... 41 Tableau 18. Degrés jours cumulés sous la bâche selon les différentes périodes durant lesquelles elle se retrouve au sol en 2016 ... 45
ix
Remerciements
Je tiens à remercier tous ceux et celles qui ont contribué de près ou de loin à la réalisation de ce mémoire. Tout d’abord, je veux remercier mon directeur, le Dr Gilles D. Leroux professeur au Département de phytologie de l’Université Laval pour sa disponibilité durant tout mon cheminement au deuxième cycle, son savoir et ses idées inspirantes. Au départ, il m’a transmis une passion pour la recherche dans le domaine de l’agriculture au moment où j’ai effectué mon stage professionnel à ses côtés. Puis, j’ai appris énormément au cours de cette maîtrise grâce à sa grande expertise.
Je tiens aussi à remercier Monsieur David Miville, professionnel de recherche au Département de phytologie de l’Université Laval, pour sa disponibilité, sa patience et son assistance technique. Aussi, je remercie Messieurs Francis Gagnon et Pascal Tessier pour leur assistance technique sur le terrain ainsi que tous les étudiants du premier cycle qui ont participé grandement aux travaux réalisés au champ.
1
1. Introduction
1.1 Problématique
Une plante adventice est « une plante qui pousse spontanément dans une culture et dont la présence est plus ou moins nocive à celle-ci » (Larousse, 2015). En effet, les plantes adventices font concurrence aux plantes cultivées, puisqu’elles peuvent germer au même moment que celles-ci et donc, empêcher le bon établissement de la culture. De plus, certaines cultures n’offrent pas une grande compétition aux adventices, par exemple le maïs (Zea mays), le lin (Linum usitatissimum) et le soya (Glycine max). Dans ce cas, une période critique d’absence de mauvaises herbes a été déterminée afin d’assurer un rendement optimal de la culture. Par exemple, les mauvaises herbes, sous certaines conditions d’humidité du sol, peuvent réduire les rendements de maïs et de soya de 18 et 23% respectivement (OMAFRA, 2009).
Le chiendent commun (Elymus repens) est une adventice qui réduit le rendement des cultures. En effet, c’est une graminée vivace qui se propage peu par ses graines, beaucoup par ses rhizomes, lesquels ont de la facilité à se régénérer et à produire de nouvelles pousses lorsque présents dans les dix premiers centimètres de sol. Le chiendent commun peut, en effet, diminuer les rendements des cultures de maïs, de soya et de blé (Triticum
aestivum) de respectivement 25 à 85%, de 19 à 55% et de 57% (OMAFRA, 1993).
Plusieurs méthodes de lutte contre le chiendent commun spécifiques pour chaque grande culture sont disponibles dans le Guide de lutte contre les mauvaises herbes publié par l’OMAFRA (2015). En effet, l’herbicide glyphosate semble une méthode efficace de lutte chimique au chiendent dans le maïs et le soya Roundup Ready®. Il offre une efficacité attendue de 90 à 95 %. Aussi, un travail de sol fait avec une charrue à versoirs ou un chisel à l’automne peut diminuer la quantité de rhizomes de chiendent viables dans le sol. Dans ce cas, les rhizomes sont enfouis dans les horizons de sol plus profonds que leur horizon naturel (0 à 10 cm de profondeur); la germination des bourgeons est ainsi diminuée (OMAFRA, 1993).
2
La solarisation pourrait être un moyen supplémentaire pour la maîtrise du chiendent. Cette méthode consiste à chauffer le sol à l’aide de bâches de plastique déposées sur le sol. La température sous la bâche et dans le sol est augmentée à des températures létales pour les racines et semences des mauvaises herbes (Horowitz et al., 1983). C’est une approche peu connue ni étudiée en ce qui concerne la maîtrise du chiendent, qui pourrait aider à diminuer le nombre de bourgeons viables sur les rhizomes ainsi que la biomasse des rhizomes. Ce procédé a été étudié aux États-Unis sur d’autres adventices soit, la lépidie à feuilles larges (Lepidium latifolium) (Hutchinson et Viers, 2011) et le kudzu (Pueraria
montana) (Newton et al., 2008). Il s’agit de plantes appartenant respectivement à la famille
des brassicacées et à la famille des fabacées. Ce sont toutes deux des mauvaises herbes vivaces introduites en Amérique du Nord et envahissantes qui se propagent par leurs rhizomes. Les bâches, mises en place dans différentes conditions, ont aidé à maîtriser l’expansion des colonies. De plus, dans les deux cas, l’effet des bâches était encore visible un an après l’arrêt des traitements. Aucune étude n’a été réalisée sur l’effet que pourraient avoir des bâches de plastique sur la croissance du chiendent commun sous les conditions du Québec.
1.2 Revue de littérature
1.2.1 Biologie du chiendent 1.2.1.1 Historique
Elymus repens (Linnaeus) Gould est le nom accepté du chiendent commun, mais peut
aussi être référé comme Agropyron repens (Linnaeus) P. Beauvois ou Elytrigia repens (Linnaeus) Desvaux ex Nevski.
Le chiendent commun est une espèce introduite en Amérique du Nord qui provient de l’Europe et de l’Asie Occidentale (Vascan, 2015). Les premiers écrits parlant de cette plante herbacée datent de 1860. Son implantation se serait produite lorsque les Européens ont introduit la culture de céréales au Canada. Aujourd’hui, le chiendent est présent partout au Canada et aux États-Unis, mais présente un problème plus au nord des États-Unis et au Canada. Dès son établissement au Canada, il était considéré comme l’une des trois pires mauvaises herbes de l’Est canadien (OMAFRA, 1993).
3
1.2.1.2 Caractéristiques
Le chiendent commun est une graminée herbacée vivace qui peut se disséminer par ses graines et se propager par ses rhizomes. Son système aérien est composé de feuilles qui ont une largeur de 9 à 10 mm et une longueur de 6 à 20 cm (Werner et Rioux, 1977). Les feuilles sont glabres, excepté les premières feuilles qui peuvent être pubescentes. À la base du limbe de la feuille, les oreillettes ont la forme de crochet entourant la tige et la ligule est membraneuse et très courte. Le plant adulte a un port dressé et peut atteindre une hauteur de 40 à 100 cm (Bouchard et Néron, 2007).
Les rhizomes servent à la propagation végétative du chiendent et peuvent atteindre une longueur de 1,5 m en une saison de croissance. Ils sont minces, blanchâtres et écailleux (Werner et Rioux, 1977). De plus, ils sont munis d’une pointe dure à leur extrémité (Bouchard et Néron, 2007).
L’inflorescence du chiendent est caractérisée par un épi allongé et mince de 5 à 20 cm de longueur (Bouchard et Néron, 2007). Les épillets sont sessiles et sont alternés sur deux rangs. Ils comprennent de 3 à 8 fleurs et mesurent de 10 à 20 mm de longueur (Werner et Rioux, 1977).
1.2.1.3 Croissance
Les graines de chiendent germent tôt au printemps et les plants commencent à produire des talles au stade 4 à 6 feuilles et des rhizomes au stade 6 à 8 feuilles. À l’opposé, les plants qui se développent suite à la germination d’un bourgeon provenant d’un rhizome commencent à produire des rhizomes au stade 3 à 4 feuilles (Figure 1). Avant, les plants utilisent leurs réserves nutritives pour croître. Les rhizomes se développent surtout durant les mois de juin, juillet et août au Québec (Werner et Rioux, 1977). Ils croissent horizontalement dans le sol à une profondeur dépassant rarement 10 cm (Laganière, 1994). En effet, lorsque des rhizomes de chiendent sont placés plus profondément que 10 cm, leur croissance est affectée. Par exemple, des rhizomes portant de 1 à 5 bourgeons, placés à une profondeur supérieure à 10 cm, s’épuisent avant même de pouvoir atteindre la surface (Vengris, 1962). À l’automne, les rhizomes sortent de terre pour former une première feuille. Par contre, sous le climat du Québec, cette feuille périra durant l’hiver. La floraison
4
a lieu vers la fin de juin et le début de juillet au Canada. Les graines deviennent matures du début d’août jusqu’au début de septembre et se séparent du plant-mère vers la fin septembre (Werner et Rioux, 1977).
Figure 1. Dispersion du chiendent par les rhizomes Tirée de : OMAFRA, 1993
Le chiendent peut croître dans tous les types de sols, mais préfère les sols bien drainés ayant une bonne structure. En effet, la croissance des rhizomes est diminuée dans les cas où le sol est compacté. Aussi, il peut croître dans un intervalle de pH allant de 4,5 à 8,0 et possède une grande tolérance aux sols salins (Werner et Rioux, 1977). L’étude de Westra et Wyse (1981), réalisée au Minnesota, rapporte une grande variation dans la croissance et le développement du chiendent due aux différents biotypes retrouvés en nature (Figure 2). En effet, l’étude a été réalisée sur 10 biotypes provenant du Minnesota qui avaient tous des caractéristiques différentes. Par exemple, le biotype 1 produit des feuilles qui restent près du sol, des rhizomes de petit diamètre et c’est ce biotype qui produit le moins d’épillets. Au contraire, la majorité des feuilles du biotype 2 se retrouvent dans les 45 premiers cm au-dessus du sol. Le biotype 6 a des feuilles mesurant de 2 à 3 mm de plus en largeur que les autres biotypes et produit le plus d’épillets par épi. Aussi, le biotype 10 est celui qui a la plus grande croissance aérienne, de rhizomes et de plants filles, mais le plus faible nombre
5
d’épis par plant. Bref, une grande variabilité dans la croissance et le développement peut être observée selon les biotypes étudiés. L’étude a aussi indiqué que si le chiendent n’est pas perturbé pendant une période allant de 2 à 3 mois, le nombre de bourgeons sur les rhizomes pouvait augmenter de 300 à 400 fois par rapport à son nombre initial.
Figure 2. Croissance des différents biotypes de chiendent commun au Minnesota Tirée de : Westra et Wise (1981)
1.2.1.4 Reproduction
Le chiendent peut assurer sa reproduction par voie sexuée, soit par la production de graines, mais il peut aussi se disperser par voie asexuée, soit par la croissance de ses
6
rhizomes. En effet, le chiendent peut produire de 15 à 400 graines par plant, mais en produit en moyenne de 25 à 40. Les graines germent à une température diume variant de 15 à 25 OC (Sagar cité par Werner et Rioux, 1977). De plus, si elles sont enfouies dans le sol, elles peuvent demeurer dormantes pendant trois ans et conserver leur viabilité pendant quatre ans. Les graines ne possèdent aucun moyen de se disséminer sur de longues distances, elles ne font que tomber sur le sol à partir du plant mère (Werner et Rioux, 1977). Aussi, dans l’étude de Westra et Wyse (1981), les plants de chiendent produisent des épis environ 2 mois et demi après leur plantation au mois de mai. Ensuite, des graines viables sont formées seulement 10 jours après l’anthèse.
Comme la production de graines par plant est relativement faible et qu’elles n’ont aucun moyen de se disperser sur de longues distances, le mode de dispersion par les rhizomes est beaucoup plus important. En effet, le diamètre de propagation des rhizomes du plant mère peut aller jusqu’à 3 m dans une saison de croissance (Werner et Rioux, 1977). De plus, selon l’étude de Westra et Wyse (1981), la majorité des biotypes ont montré un diamètre de propagation d’environ 1,5 m autour du plant mère 2,5 mois après la plantation au mois de mai. Aussi, lors d’une autre récolte réalisée 3,5 mois après la plantation, le diamètre de propagation des rhizomes de chiendent avait augmenté de 63 % en comparaison avec la première récolte.
Chaque rhizome porte un grand nombre de bourgeons, mais ce ne sont pas tous les bourgeons présents sur les rhizomes qui sont aptes à germer et à produire des plants. En effet, ces bourgeons peuvent rester dormants pendant presque toute la vie du rhizome. Cette dormance peut être expliquée par la dominance qu’exerce le bourgeon apical sur les bourgeons axillaires. Cette dominance est maintenue grâce à l’effet qu’exercent certaines substances de croissance sur les bourgeons. Les substances de croissance responsables de cette dominance apicale sont l’auxine et les cytokinines (Su Y.-H. et al., 2011). L’auxine, lorsqu’elle se retrouve en grande concentration (1 ppm) dans une tige ou un rhizome, stimule la croissance du bourgeon apical, mais inhibe la croissance des bourgeons axillaires. Par contre, à de faibles concentrations (0,01 ppm), l’auxine favorise la croissance des bourgeons latéraux. Donc, lorsque l’apex est brisé, les concentrations d’auxines diminuent, ce qui favorise la croissance des bourgeons latéraux. Aussi, les cytokinines
7
favorisent la translocation des substances nutritives vers les bourgeons. Elles ont donc un rôle antagoniste aux auxines, mais elles agissent seulement en présence d’auxines. Donc, si le rapport cytokinines/auxines est faible, il y aura de la dominance apicale, mais si ce rapport est élevé, il y aura levée de la dominance apicale (Rioux, 2015).
1.2.2 Nuisibilité en agriculture 1.2.2.1 Présence dans les cultures
Le chiendent est nuisible en agriculture puisqu’il peut causer des diminutions de rendement dans plusieurs types de cultures comme le soya, le maïs, le canola (Brassica
napus) et les céréales s’il n’est pas maîtrisé.
En effet, Young et al. (1982) ont mesuré l’effet que peut avoir le chiendent sur le soya. Lorsque le chiendent est présent à de fortes densités (520 et 910 tiges/m2), la hauteur, le nombre de branches, de gousses et de grains par plant ont diminué significativement lors de la récolte du soya. De plus, le rendement du soya a diminué selon différentes densités de chiendent et selon la quantité de précipitations qu’ont reçues les parcelles. En effet, plus le sol est humide, plus il est nécessaire d’avoir de grandes densités de chiendent pour réduire le rendement du soya. Aussi, lorsque le chiendent était présent plus de 4 semaines avec le soya, la hauteur, le nombre de branches produisant des gousses et le rendement en grains étaient diminués. Par contre, les chercheurs ont conclu qu’il existe une assez longue période de temps entre le semis du soya et la levée du chiendent pour que soient utilisés des herbicides en post-levée, sans affecter le rendement du soya. Par exemple, des applications d’ASSURE II à raison de 0,75 L/ha sont recommandées contre le chiendent commun en post-levée dans le soya par le Guide de Lutte contre les Mauvaises Herbes 2016-2017 de l’OMAFRA, avec une efficacité attendue de 70 à 85 %.
Les mêmes auteurs (1984) ont mesuré l’effet du chiendent sur les rendements de maïs à deux sites différents. Une densité de 745 tiges de chiendent/m2 a réduit la croissance du maïs et son rendement. De plus, les populations les plus élevées de chiendent pour les deux sites, soit de 695 et 645 tiges/m2, ont réduit les rendements respectivement de 29 et 58 %. Par contre, les densités minimales de chiendent requises pour diminuer significativement le rendement du maïs varient selon les précipitations reçues durant la saison de croissance. Par
8
exemple, des densités de 600 et de 900 tiges/m2 ont diminué les rendements de 13 % lorsque les précipitations étaient abondantes. Des résultats similaires ont été observés avec des densités de chiendent de 55 et 75 tiges/m2 lorsque les précipitations étaient plus faibles.
Le canola est une autre culture qui est influencée par la présence du chiendent. En effet, O’Donovan (1991) a déterminé que 100 tiges de chiendent /m2
pouvaient diminuer le rendement du canola de 27 à 36 %. Aussi, plus les populations de chiendent augmentent, plus les pertes de rendement sont importantes jusqu’à l’obtention d’une population maximale de chiendent, ensuite les pertes de rendement ne changent plus significativement.
Bref, lorsqu’il est présent à une faible densité, le chiendent ne cause pas beaucoup de pertes de rendement des cultures. Par contre, plus sa population augmente, plus il diminue les rendements de la majorité des cultures.
1.2.2.2 Allélopathie du chiendent
Les rhizomes et les feuilles de chiendent en décomposition dans le sol produisent une toxine qui a un effet allélopathique sur la germination et la croissance des plantes cultivées. Il a été démontré que les feuilles du chiendent en décomposition dans le sol sont plus toxiques que les rhizomes à certains stades de croissance et à certaines concentrations. En effet, les feuilles récoltées au stade végétatif ont un effet plus marqué sur les rendements de la luzerne (Medicago sativa) que celles récoltées à des stades ultérieurs. Ainsi, l’activité allélopathique peut causer des diminutions de rendement dans les cultures comme la luzerne. Par exemple, le rendement de la luzerne a été influencé par les résidus de chiendent incorporés au sol. Par contre, le rendement a été plus affecté lorsque le chiendent était incorporé 1 semaine avant le semis que lorsqu’il était incorporé 3 semaines avant le semis de la luzerne (Touchette, Leroux et Deschênes, 1988).
Une autre expérience a montré que les extraits aqueux de feuilles et de rhizomes de chiendent possèdent au moins deux inhibiteurs qui sont des composés flavonoïdes. Ces composés réduisent la croissance aérienne et racinaire des plantes cultivées et des adventices. De plus, ils réduisent la nodulation des légumineuses et la fixation de l’azote par Rhizobium. Aussi, ces auteurs ont confirmé que les feuilles de chiendent sont plus
9
toxiques que les rhizomes, car ils contiennent une plus grande quantité de ces composés (Weston, Burke et Putnam, 1986).
1.2.3 Méthodes de lutte 1.2.3.1 Chimique
Plusieurs herbicides sont homologués au Canada pour lutter contre le chiendent adéquatement. Par exemple, le foramsulfuron (OPTION 2.25 OD©), le rimsulfuron et le nicosulfuron (ULTIM© et ACCENT©) et le glyphosate (ROUNDUP WEATHERMAX©) sont tous des herbicides efficaces contre le chiendent (OMAFRA, 2015).
Le glyphosate est un herbicide systémique du groupe 9, inhibiteur de la synthèse des acides aminés aromatiques, qui a un large champ d’activité (OMAFRA, 2000). Il est principalement utilisé dans les cultures de maïs et de soya « Roundup Ready » pour éliminer les adventices. Lorsqu’il est appliqué sur le chiendent, il est rapidement absorbé et transloqué dans toute la plante. Dans l’étude de Sprankle et al. (1975) pour aider à l’absorption du produit, un adjuvant était ajouté au mélange. L’absorption du glyphosate se faisait alors dans les quatre heures suivant le traitement. Par contre, si une pluie survient de 0 à 8 heures après l’application, l’effet du glyphosate diminue dû au lessivage du produit. De plus, un labour peut affecter la translocation du glyphosate dans le chiendent. En effet, le labour sépare les rhizomes du plant-mère et empêche donc la translocation du glyphosate vers les bourgeons. Labourer un jour après une application de glyphosate peut permettre de mieux maîtriser le chiendent. Au contraire, lorsque le labour est fait le jour du traitement, le produit n’a pas le temps d’être transloqué dans toutes les parties de la plante.
De même, des applications de glyphosate peuvent réduire la densité de tiges de chiendent, peu importe le degré d’infestation initial. Par contre, la maîtrise est plus efficace lorsque les populations de chiendent sont plus faibles. En effet, en comparaison avec des traitements témoins sans herbicide, le nombre de tiges de chiendent est diminué de 63, 74 et 35 % pour respectivement une faible, moyenne et forte population traitée avec 0,45 kg/ha de glyphosate. Lorsque les populations sont traitées avec 0,90 kg/ha de glyphosate, la maîtrise du chiendent est meilleure. Bref, lorsque l’infestation initiale est élevée (180 tiges/m2) une maîtrise maximale du chiendent est obtenue avec une dose de 0,90 kg/ha de
10 glyphosate combiné à de l’ethylan (Frigate®
) ou encore avec 1,80 kg/ha de glyphosate seul. De plus, lorsque l’infestation est à un degré moyen (72 tiges/m2), la maîtrise avec 0,45 kg/ha de glyphosate combiné à de l’ethylan est aussi efficace que les traitements avec de plus fortes doses (Leroux, 1993).
Le glyphosate peut aussi affecter la biomasse des rhizomes de chiendent. En effet, lorsque le chiendent est traité avec 0,90 ou 1,80 kg/ha de glyphosate, il n’y a pas de différence entre les biomasses de rhizomes, mais celles-ci sont plus élevées lorsque le chiendent est traité avec 0,45 kg/ha. Le glyphosate maîtrise mieux les rhizomes lorsque ceux-ci sont en plus faibles populations, peu importe la dose utilisée. Par contre, en tout temps, la maîtrise des rhizomes est moins importante que la maîtrise des parties aériennes (Leroux, 1993).
1.2.3.2 Non chimique
1.2.3.2.1 Méthodes culturales
Le travail du sol, les rotations de cultures ainsi que le type de cultures utilisé sont tous des moyens pour aider à lutter contre le chiendent. Le labour, effectué à l’aide d’une charrue à versoir, peut aider à maîtriser la croissance du chiendent en comparaison avec un système où aucun travail du sol ne serait fait. Un labour conventionnel peut diminuer de 83 % le nombre de bourgeons viables sur les rhizomes du chiendent, contrairement à un travail du sol réduit qui diminue le nombre de bourgeons viables sur les rhizomes de 35 % (Chandler, Murphy et Swanton, 1994). Donc, plus de bourgeons demeurent viables à la suite du travail réduit du sol qu'à la suite du labour. En effet, le labour déplace les rhizomes de chiendent à des profondeurs supérieures à 10 cm, soit celles qui sont optimales pour leur croissance. Les pousses s’épuiseront alors avant d’atteindre la surface (Duval, 2004).
Par ailleurs, le moment où le labour est pratiqué a de l’importance sur la maîtrise du chiendent. Un labour effectué à l’automne aurait tendance à mieux maîtriser le chiendent qu’un labour effectué au printemps. À l’automne, les rhizomes sont exposés à des températures plus faibles et sont à risque de dessèchement durant l’hiver (Chandler, Murphy et Swanton, 1994). Il a été démontré, au contraire, dans d’autres travaux qu’un labour effectué au printemps contribuerait mieux à détruire les rhizomes de chiendent. En
11
effet, dans l’étude effectuée par Majek, Erickson et Duke (1984), c’est lors du labour de printemps que les bourgeons présents sur les rhizomes ont le moins survécu. En effet, tous les rhizomes présents dans le sol à ce moment sont des rhizomes qui ont été produits l’année précédente. À l’opposé, lorsque le sol est travaillé en juin ou en août, les rhizomes présents dans le sol sont une composition de rhizomes de l’année précédente ainsi que de nouveaux rhizomes. Plusieurs de ces nouveaux rhizomes ont tendance à survivre au labour tandis que les plus vieux ne survivent pas. Bref, pour réduire la viabilité des bourgeons des rhizomes de chiendent, il faudrait prévenir la croissance de nouveaux rhizomes. Donc, le travail du sol devrait être fait en mai ou avant que le chiendent n’atteigne 6 feuilles, soit au moment où il commence sa production de rhizomes.
De plus, les différents types de travail du sol peuvent affecter la distribution des rhizomes de chiendent dans le profil du sol. En effet, lorsque le sol n’est pas travaillé ou travaillé au printemps avec un chisel, 83 % des rhizomes se retrouvent dans les premiers 7,5 cm de sol. Au contraire, lorsque le sol est travaillé à l’automne ou au printemps avec une charrue à versoirs, 56 et 47 % des rhizomes respectivement se retrouvent dans les premiers 7,5 cm de sol, le reste étant enfouis plus profondément. Lorsque les rhizomes sont enfouis à des profondeurs supérieures à celles où on les retrouve normalement (0-10 cm), ils s’épuisent avant d’atteindre la surface (Chandler, Murphy et Swanton, 1994).
Une autre technique utilisée pour éliminer le chiendent est appelée épuisement. Cette technique est décrite par Duval (2004) et consiste à segmenter finement et à répétition les rhizomes du chiendent. Une herse à disque ou un rotoculteur peuvent être utilisés pour cette opération. Tout d’abord, un premier travail de sol est effectué à une profondeur de 5 à 15 cm suite à une récolte de céréale par exemple. Les rhizomes sont donc coupés une première fois. Le deuxième passage peut être effectué à nouveau lorsque le chiendent atteint un stade de 3 à 4 feuilles, soit au moment où ses réserves sont au plus bas, ou effectué 21 jours après le premier passage. Plus les rhizomes sont segmentés souvent, plus la germination des bourgeons sur les rhizomes sera stimulée et ceux-ci s’épuiseront avant de pouvoir atteindre la surface du sol.
Le type de culture utilisée dans la rotation peut aussi affecter la croissance du chiendent. En effet, plusieurs cultures sont sensibles au chiendent et peuvent avoir des
12
rendements réduits (maïs, soya et canola). D’autres cultures sont très compétitives au chiendent et peuvent nuire à sa croissance. Par exemple, le fait d’utiliser une céréale d’automne, en remplacement d’une céréale de printemps dans la rotation, diminue le développement du chiendent. La céréale d’automne couvrira le sol durant la fin de l’été et l’automne et entrera en compétition avec le chiendent. De plus, la céréale d’automne reprend sa croissance tôt au printemps, entrant directement en compétition avec la croissance du chiendent. Par contre, si le blé d’automne ne s’établit pas correctement, dû aux conditions de sécheresse et de température par exemple, la croissance du chiendent ne sera pas affectée. Bref, le type de culture utilisé, plus particulièrement dans un système de travail réduit du sol, peut aider à maîtriser le chiendent (Loeppky et Derksen, 1994).
L’utilisation d’engrais verts pourrait aider à réduire la croissance du chiendent. En effet, l’engrais vert d’automne est particulièrement efficace pour maîtriser le chiendent. Plus précisément, le sarrasin (Fagopyrum esculentum) peut être utilisé comme engrais vert durant l’été pour limiter la croissance du chiendent (Jobin et Douville, 1997). En effet, celui-ci germe et croît très rapidement, entrant directement en compétition pour la lumière avec le chiendent. Par contre, cette compétition est de courte durée puisque, lorsque les fleurs sont formées, la lumière recommence à atteindre le sol. Le chiendent peut alors recommencer à se développer (Duval, 2004).
1.2.3.2.2 Solarisation
D’autres méthodes de lutte peuvent être utilisées pour maîtriser les adventices. En effet, la solarisation est un moyen de réchauffer le sol à l’aide de bâches de plastique placées à la surface du sol qui captent l’énergie solaire. Cette méthode sert à augmenter la température du sol afin de diminuer la viabilité des semences retrouvées dans le sol. Différents types de plastiques peuvent être utilisés. Une étude réalisée par Horowitz, Regev et Herzlinger en Israël (1983) a montré la différence d’effet entre les plastiques transparents et les plastiques opaques noirs. Premièrement, les chercheurs ont remarqué une maîtrise efficace des mauvaises herbes dans les parcelles qui ont reçu des traitements avec une bâche de plastique noire ou transparente. Par contre, la bâche de plastique transparente semblait être plus efficace que la noire. En effet, la proportion d’adventices annuelles ayant émergé deux mois après le retrait des bâches, par rapport au traitement témoin enherbé a été de 15 % et
13
3,4 % pour respectivement les plastiques noir et transparent. En effet, certaines adventices comme le liseron des champs (Convolvulus arvensis), le pourpier potager (Portulaca
oleracea) et les amarantes (Amaranthus spp.), ont été réprimées seulement par la bâche
transparente. Aussi, la germination des semences de mauvaises herbes a été plus faible suite à l’application et le retrait de la bâche transparente qu’avec la bâche noire. Suite au retrait des bâches, l’effet de la bâche de plastique noire s’est maintenu pendant moins de quatre mois tandis que celui de la bâche transparente s’est maintenu pendant un an. La température obtenue sous les bâches a augmenté d’environ 9 OC et 18 OC respectivement avec les bâches noires et transparentes, à une profondeur de 5 cm dans le sol.
Rubin et Benjamin (1983) ont montré que certaines mauvaises herbes vivaces (Cyperus
rotundus, Sorghum halepense, Cynodon dactylon) sont seulement partiellement maîtrisées
lorsqu’une bâche transparente est utilisée pendant une courte période, soit de 4 et 5 semaines. Par contre, une bâche installée de 8 à 10 semaines améliore significativement la maîtrise de ces adventices, mais ces espèces demeurent présentes grâce à leurs organes de régénération végétative, situés dans des horizons de sol plus profonds (supérieur à 10 cm). Au contraire, l’utilisation d’une bâche de plastique opaque avait, en général, un effet moindre sur les adventices vivaces. Par contre, une bâche opaque mise sur le sol pendant 7 semaines a produit de meilleurs résultats que tous les autres traitements. L’ombrage pourrait avoir eu un effet additionnel à l’augmentation de température du sol.
Une étude réalisée aux États-Unis par Hutchinson et Viers dans les années 2000 portait sur l’effet des bâches de plastique noire sur la lépidie à feuilles larges (Lepidium
latifolium). Cette espèce vivace est considérée comme envahissante dans l’ouest des
États-Unis. Elle se disperse surtout par ses graines et ses racines qui lui permettent de s’établir dans presque tous les environnements. Dans cette étude, les auteurs ont voulu comparer l’effet des bâches noires à l’effet d’herbicides connus, soit le glyphosate et le chlorsulfuron. L’application des bâches était précédée d’une tonte seulement ou d’une tonte suivie d’un labour. Les bâches sont demeurées en place durant deux ans.
Les résultats pour le traitement de tonte suivie de la bâche ont été variables et n’ont pas affecté significativement la lépidie à feuilles larges. Par contre, le traitement où un labour a été fait avant la mise en place de la bâche a réduit la densité des tiges de 94 % mesurée
14
deux ans après le retrait de la bâche. Les résultats de l’étude suggèrent que la tonte suivie du labour a mieux maîtrisé les rhizomes de la mauvaise herbe que la tonte seule avant la pose de la bâche. Aussi, l’efficacité de la bâche dépendrait de la densité initialement présente de lépidie à feuilles larges où celle-ci serait plus variable au fur et à mesure que la densité de la population augmente. En effet, ces traitements pourraient être utilisés lorsque des populations initiales faibles menacent d’infester de plus grandes superficies (Hutchinson et Viers, 2011).
De plus, un an et deux ans après la mise en place des traitements, les résultats étaient semblables à ceux obtenus avec les traitements de glyphosate, de chlorsulfuron ou de tonte + labour + bâche. Donc, là où l’utilisation des herbicides est restreinte, le traitement de tonte + labour + bâche pourrait être une bonne option. Par contre, il est vrai que la mise en place des bâches et le labour demandent plus de temps qu’une application d’herbicides. De plus, cette méthode est limitée par la confection des bâches, la méthode d’application sur le terrain et le sol qui est laissé à nu, le laissant exposé à des infestations futures par d’autres mauvaises herbes envahissantes (Hutchinson et al., 2011).
Une étude de Newton et al. (2008) a été réalisée sur Pueraria montana (kudzu), une espèce vivace envahissante aux États-Unis et dans le nord de l’Ontario. L’objectif de l’étude était de vérifier l’effet des bâches transparentes sur le nombre de tiges vivantes et mortes de kudzu. En premier lieu, on voulait savoir s’il existait des différences entre l’utilisation de bâches transparentes et de bâches opaques, ceci afin de déterminer laquelle utiliser. Newton et al. (2008) ont pu conclure que les températures moyennes sous les deux types de bâches ne différaient pas significativement. Les auteurs ont décidé d’utiliser la bâche transparente puisque les résultats suggéraient une plus grande uniformité des températures sous cette bâche. Par la suite, la bâche a été installée soit tout l’été, soit à des intervalles de 1, 2 et 4 semaines. Par exemple, pour l’intervalle de 4 semaines, la bâche était installée pendant 1 semaine puis le sol était à découvert pendant 4 semaines. Pour l’intervalle de 2 semaines, la bâche était installée pendant 1 semaine puis retirée pour les 2 semaines suivantes. Les intervalles ont été répétés durant toute la saison de croissance. Suite à l’expérimentation en 2005, l’étude a permis de conclure que le traitement pour lequel la bâche est demeurée durant toute la saison de croissance a eu l’effet le plus néfaste
15
sur le nombre de tiges de kudzu, suivi par le traitement à intervalle de 1 semaine puis par celui à intervalle de 4 semaines. Les taux de mortalité des tiges étaient respectivement de 44, 36 et 22 %. En comparaison, dans la parcelle où le kudzu n’a pas été traité, seulement 2 % de mortalité a été enregistré. Les auteurs ont recommencé les mêmes traitements en 2006 sur les mêmes parcelles et ont enregistré un taux de mortalité en moyenne de 97% pour tous les traitements avec bâche. Bref, deux années consécutives avec une bâche seraient nécessaires pour éradiquer complètement le kudzu.
1.3 Hypothèses de travail
1. L’application d’une bâche de plastique opaque sur un sol infesté de chiendent procure une maîtrise efficace de l’adventice;
2. L’application d’une bâche de plastique opaque durant toute la saison de croissance procure une maîtrise supérieure du chiendent comparativement à l’application de bâche durant une plus courte période de temps;
3. L’installation de la bâche sur une courte période de temps aurait une efficacité différente sur le chiendent selon qu’elle est installée au printemps ou à l’automne; 4. Le travail du sol est plus efficace à maîtriser le chiendent que la fauche
lorsqu’effectué avant la mise en place d’une bâche.
1.4 Objectifs de la recherche
1. Montrer que l’application d’une bâche de plastique opaque diminue la biomasse sèche des rhizomes et des bourgeons sur les rhizomes du chiendent;
2. Déterminer s’il existe une durée de temps minimale nécessaire pour l’application des bâches afin d’obtenir une efficacité maximale;
3. Déterminer s’il existe un moment préférable durant la saison pour la mise en place la bâche;
4. Vérifier l’effet du travail du sol ou de la fauche préalablement à la mise en place d’une bâche.
16
2. Matériel et méthodes
2.1 Sites expérimentaux
Le projet de recherche a eu lieu sur deux champs infestés par le chiendent à la station agronomique de l’Université Laval à Saint-Augustin-de-Desmaures. Ces sites n’avaient reçu aucun traitement herbicide de désherbage chimique contre le chiendent depuis 10 ans. Le site 1 est le lieu de l’expérimentation pour l’année 2015 et le site 2, celui pour l’année 2016 (Figure 3). À ces deux sites, le sol est caractérisé par une texture de sol loam argileux. Avant la mise en place des traitements, une herse à disques, ajustée à une profondeur de 10 cm, ainsi qu’un rouleau Brillion© ont été passés dans le sens nord-sud, à deux reprises sur la totalité des sites.
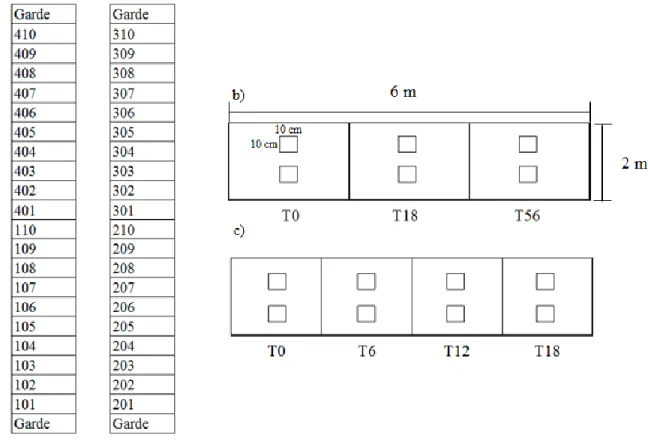
Figure 3. Sites expérimentaux de 2015 (site 1) et 2016 (site 2) à la station Agronomique de Saint-Augustin-de-Desmaures
(46°44'03.7"N 71°30'23.9"W) Tirée de : Google maps Adaptée par : Amélie Picard
17
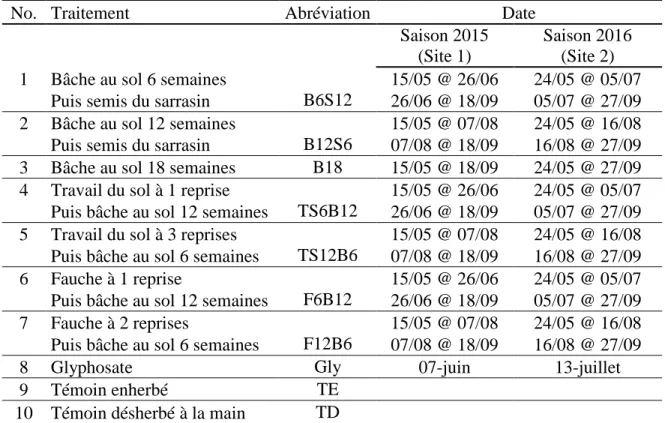
Le site 1 était caractérisé, le 5 mai 2015, par un nombre moyen de 140 tiges de chiendent dans 0,25 m2 et un recouvrement moyen du chiendent de 45 %. D’autres adventices étaient présentes, mais en plus petit nombre, soit le tussilage pas-d’âne (Tussilago farfara), l’asclépiade (Asclepias syriaca), le pissenlit (Taraxacum officinale) et le laiteron des champs (Sonchus arvensis). Un total de dix traitements ont été répartis selon un plan en bloc complet aléatoire avec quatre répétitions pour un total de 40 parcelles expérimentales. Les parcelles ont été disposées de façon à recouvrir la presque totalité du site et les blocs ont été placés de façon à regrouper dans un même bloc, les populations semblables de chiendent. Les parcelles avaient une dimension de 2 mètres de largeur par 6 mètres de longueur et ont été divisées en 3 sous-parcelles de 2 par 1,5 mètre pour permettre l’échantillonnage des parcelles. Un espace de 50 cm au début et à la fin de la parcelle a été éliminé de la zone d’échantillonnage pour éviter les biais. L’échantillonnage des rhizomes de chiendent s’est effectué avant la mise en place des traitements (T0), 18 semaines après la mise en place des bâches (T18) et l’année suivant le retrait des bâches (T56). Aussi, des parcelles de garde ont été placées aux extrémités nord et sud des répétitions (Figure 4). Le site 2 était recouvert par le chiendent à 65%, le 18 mai 2016, et comptait en moyenne 135 tiges de chiendent dans 0,25 m2. Aussi d’autres adventices étaient présentes en plus petits nombres, soit le pissenlit, le laiteron des champs, le chardon des champs (Cirsium
arvense), la moutarde des champs (Sinapis arvensis) et le tussilage pas-d’âne. De la même
façon que pour 2015, dix traitements ont été répartis selon un plan en blocs complets aléatoires avec quatre répétitions des traitements pour un total de 40 parcelles expérimentales. Les parcelles avaient une dimension de 2 par 6 mètres et, tel qu’en 2015, un espace de 50 cm au début et à la fin de la parcelle a été retiré de la zone d’échantillonnage. Les parcelles ont été séparées en quatre sous-parcelles de 2 par 1 mètres pour l’échantillonnage au temps zéro (T0) ainsi qu’à 6 (T6), 12 (T12) et 18 (T18) semaines après la mise en place des bâches. Des parcelles de garde ont été disposées aux extrémités nord et sud des parcelles (Figure 4).
18
T0 T6 T12 T18
Figure 4. Plan détaillé des parcelles et sous-parcelles, leurs dimensions ainsi que des zones d’échantillonnage du sol dans chaque parcelle
2.2 Dispositifs expérimentaux
2.2.1 Matériel utilisé et mise en place des traitements
La bâche utilisée est une bâche de polyéthylène opaque avec un côté blanc et un côté noir. Elle est utilisée en agriculture pour l’ensilage de plantes fourragères. Les caractéristiques de la bâche sont présentées à l’annexe 1. Cette bâche a une masse volumique apparente de 580 kg/m3 et une durée de vie d’environ 5 ans. La bâche a été découpée à un format de 1,50 par 6,10 mètres pour convenir à la taille des parcelles.
a) Présentation de la disposition des parcelles aux sites 1 et 2; b) plan d'une parcelle en 2015, de ses divisions en sous-parcelles et des zones d’échantillonnage du sol dans chaque division; c) plan d'une parcelle en 2016, de ses divisions en sous-parcelles et des zones d’échantillonnage dans chaque division
19
Pour s’assurer du maintien au sol des bâches durant toute la saison, des œillets ont été installés à tous les mètres sur le sens de la longueur de la bâche. Par la suite, une tranchée a été creusée au champ sur le périmètre de chaque parcelle à une profondeur de 20 cm. Cette tranchée servait à enfouir les rebords de la bâche, mais aussi à offrir une barrière physique au chiendent. La bâche a été disposée dans cette tranchée et des clous de 10 cm de longueur ont été insérés dans les œillets et dans le sol pour maintenir la bâche en place. Par la suite, les parties de la bâche qui se retrouvaient dans la tranchée ont été recouvertes de terre (Figure 5).
Figure 5. Étapes de mise en place de la bâche dans les parcelles des traitements 1 à 7
La bâche a été installée dès le début de la saison de croissance pour les traitements 1 (Bâche au sol 6 semaines, puis semis du sarrasin), 2 (Bâche au sol 12 semaines, puis semis du sarrasin) et 3 (Bâche au sol 18 semaines) (Tableau 1). Les bâches sont restées en place pour des périodes respectives de 6, 12 et 18 semaines. Par la suite, pour les traitements 1 et 2, lorsque la bâche était retirée, un semis de sarrasin a été effectué. Les semences de sarrasin utilisées ont été conservées à température de la pièce jusqu’au semis et une dose de semis de 90 kg de graines viables par hectare a été utilisée. Pour obtenir un
a) b) c)
a) une tranchée de 20 cm de profondeur est creusée sur le périmètre de la parcelle; b) la bâche est mise en place dans la tranchée et demeure en place grâce à des œillets installés sur la bâche et à des clous de 10 cm de longueur; c) les rebords de la bâche sont recouverts de terre.
20
lit de semences adéquat, un rotoculteur a été passé à deux reprises dans les parcelles avant le semis de sarrasin, travaillant le sol à une profondeur de 5 cm. Ensuite, le semis a été fait manuellement à la volée et un râteau a été passé dans les parcelles pour enfouir les graines et assurer une levée uniforme. Le sarrasin offrait une couverture du sol lorsque les bâches étaient retirées.
Les traitements 4 (Travail du sol à 1 reprise, puis bâche au sol 12 semaines) et 5 (Travail du sol à 3 reprises, puis bâche au sol 6 semaines) consistaient en un travail de sol à respectivement une et trois reprises avec un rotoculteur et, à chaque fois, le rotoculteur effectuait deux passages dans chaque parcelle (Tableau 2). La bâche a été installée sur ces parcelles par la suite et est demeurée en place pour des durées respectives de 12 et 6 semaines (Tableau 1).
La végétation a été fauchée à une et à deux reprises dans respectivement les parcelles des traitements 6 (Fauche à 1 reprise, puis bâche au sol 12 semaines) et 7 (Fauche à 2 reprises, puis bâche au sol 6 semaines). La végétation était fauchée à une hauteur de 5 cm du sol (Tableau 2). Les bâches ont été installées par la suite et sont restées en place pour des durées respectives de 12 et 6 semaines (Tableau 1).
Les traitements 8, 9 et 10 constituaient les traitements témoins de l’expérience. Le traitement 8 a reçu une application de 900 grammes par hectare de glyphosate (Roundup Weathermax© 540 SN) dans un volume d’application de 200 l/ha. Le traitement a été effectué à l’aide d’un pulvérisateur à dos relié à une bonbonne de CO2 comportant une
rampe avec quatre buses TJ-11002 DG espacées de 50 cm. Les conditions lors de la pulvérisation sont présentées dans le tableau 3. Le traitement 9 n’a reçu aucune intervention durant toute la saison de croissance, ce qui constituait le traitement témoin enherbé (Tableau 1). Dans les parcelles du traitement 10, le chiendent a été coupé au niveau du sol avec un coupe bordure à chaque fois que le regain arrivait au stade de 2 feuilles. Ce traitement 10 constituait le traitement témoin désherbé à la main.
21
Tableau 1. Protocole expérimental pour 2015 et 2016 avec le numéro des traitements, leurs descriptions et abréviations ainsi que les dates de mise en place de ces traitements
No. Traitement Abréviation Date
Saison 2015 (Site 1)
Saison 2016 (Site 2)
1 Bâche au sol 6 semaines 15/05 @ 26/06 24/05 @ 05/07
Puis semis du sarrasin B6S12 26/06 @ 18/09 05/07 @ 27/09
2 Bâche au sol 12 semaines 15/05 @ 07/08 24/05 @ 16/08
Puis semis du sarrasin B12S6 07/08 @ 18/09 16/08 @ 27/09 3 Bâche au sol 18 semaines B18 15/05 @ 18/09 24/05 @ 27/09 4 Travail du sol à 1 reprise 15/05 @ 26/06 24/05 @ 05/07 Puis bâche au sol 12 semaines TS6B12 26/06 @ 18/09 05/07 @ 27/09 5 Travail du sol à 3 reprises 15/05 @ 07/08 24/05 @ 16/08 Puis bâche au sol 6 semaines TS12B6 07/08 @ 18/09 16/08 @ 27/09
6 Fauche à 1 reprise 15/05 @ 26/06 24/05 @ 05/07
Puis bâche au sol 12 semaines F6B12 26/06 @ 18/09 05/07 @ 27/09
7 Fauche à 2 reprises 15/05 @ 07/08 24/05 @ 16/08
Puis bâche au sol 6 semaines F12B6 07/08 @ 18/09 16/08 @ 27/09
8 Glyphosate Gly 07-juin 13-juillet
9 Témoin enherbé TE
10 Témoin désherbé à la main TD
Tableau 2. Dates des interventions de travail de sol et de fauche aux 2 sites pour les traitements 4 à 7
No. Traitement
Date de l’intervention (travail de sol ou fauche) Saison 2015
(Site 1)
Saison 2016 (Site 2) 4. Travail du sol à 1 reprise 04-juin 10-juin
Puis bâche au sol 12 semaines
5. Travail du sol à 3 reprises 04-juin 10-juin Puis bâche au sol 6 semaines 26-juin 12-juillet
29-juillet 15-août
6. Fauche à 1 reprise 04-juin 10-juin
Puis bâche au sol 12 semaines
7. Fauche à 2 reprises 04-juin 10-juin
22
Tableau 3. Conditions lors de l'application du traitement de glyphosate (8)
Paramètre Année d'application 2015 (Site 1) 2016 (Site 2)
Date / Heure 6 juillet / 6h du
matin
13 juillet / 6h du matin Humidité du sol à 5 cm Très humide Très humide
Ennuagement Faible Faible
Température air (°C) 16 18,5
Température sol à 10 cm (°C) 7 19
Humidité relative (%) 91,3 99
Condition du couvert Forte rosée Légère rosée
Vent (km/h) 0 0 - 5
Stade de croissance du
chiendent Épiaison 3-4 Feuilles
2.2.2 Variables mesurées
Les variables dépendantes mesurées dans cette étude étaient la biomasse sèche des rhizomes sains ainsi que le nombre de bourgeons présents sur ces rhizomes. Pour ce faire, des quadrats de 10 cm x 10 cm ont été utilisés et ont été enfoncés dans le sol à une profondeur de 10 cm (Figure 6). Ces quadrats ont permis d’obtenir des échantillons de sol qui contenait les rhizomes de chiendent. À chaque moment d’échantillonnage (T0, T6, T12, T18 et T56), deux échantillons de sol ont été prélevés par parcelle. Les échantillons ont été conservés dans un réfrigérateur maintenu à 4 oC jusqu’au nettoyage. Le nettoyage servait à trier les rhizomes sains du sol. Une fois les rhizomes bien nettoyés, chaque bourgeon présent sur les rhizomes était compté. Par la suite, les rhizomes ont été séchés à 65 oC jusqu’à un poids constant puis pesés pour obtenir la biomasse sèche des rhizomes.
Des sondes de températures de type HOBO® ont été installées à la surface du sol sous la bâche du traitement 3 (bâche au sol 18 semaines) et dans le traitement témoin enherbé sans bâche aux deux sites. Les huit sondes sont restées en place du 10 juin au 18 septembre 2015 et du 24 mai au 26 septembre 2016.
23
a) b) c)
Figure 6. Étapes pour la réalisation des échantillonnages du sol dans les sous parcelles à l’aide des quadrats
a) Le quadrat de 10 X 10 X 10 cm est placé à la surface du sol vis-à-vis l’endroit où l’échantillonnage aura lieu; b) le quadrat est enfoncé dans le sol à l’aide d’une masse; c) le quadrat est retiré du sol et l’échantillon est recueilli.
2.3 Analyses statistiques
Les données de biomasse sèche de rhizomes et de décompte des bourgeons ont été traitées séparément à l’aide du logiciel SAS University Édition©. À titre de rappel, il y avait dix traitements répétés quatre fois dans un dispositif en bloc complet aléatoire. La procédure MIXED de SAS a été utilisée pour traiter les données, puisqu’il s’agissait d’un plan en bloc complet aléatoire avec des mesures répétées dans le temps. Comme le site 1 n’a pas été échantillonné au T6 et au T12, et que le site 2 n’a pas été échantillonné au T56, il s’agit d’un design incomplet. Donc, lorsque des moyennes sont calculées dans les tests des effets globaux, celles-ci ne prennent en compte que les sites où il y a des observations. Les données ont été soumises à l’Analyse de Variance (ANOVA) pour vérifier s’il existait des interactions entre les différents facteurs à l’étude (traitements, temps, sites). Les interactions possibles sont :
24
- Traitements * Temps : le temps correspond aux échantillons pris dans une même parcelle à différentes dates sur le même site. Si cette interaction est significative, cela signifie que les traitements ont eu un effet différent sur le chiendent selon le moment où l’échantillonnage a été effectué;
- Traitements * Sites : si cette interaction est significative, cela indique que les traitements ont des effets différents selon le site à l’étude;
- Temps * Sites : si cette interaction est significative, cela indique que les données à chaque date d’échantillonnage ont été différentes selon le site à l’étude;
- Traitements * Sites * Temps : une interaction triple significative démontrerait des interactions entre les traitements, les sites et le temps sur les variables à l’étude.
Les données ont dû subir une transformation racine carrée pour améliorer l’homogénéité de la variance et la normalité des résidus. Par la suite, si les tests globaux des traitements étaient significatifs, ils ont été soumis à des comparaisons simples établies a priori. Ainsi, ces comparaisons permettront de confirmer ou de réfuter les hypothèses de départ. La description des comparaisons simples est présentée au tableau 4 et les coefficients des comparaisons simples à un degré de liberté sont décrits à l’annexe 2.
25
Tableau 4. Description des comparaisons simples et de leurs traitements associés
No. Comparaison simple1 Abréviation Traitements comparés
1 Glyphosate Gly vs 8
vs traitements de bâche TB vs 1, 2, 3, 4, 5, 6 et 7
2 Traitement témoin enherbé TE vs 9
vs traitements de bâche TB vs 1, 2, 3, 4, 5, 6 et 7
3 Traitement témoin désherbé TD vs 10
vs traitements de bâche TB vs 1, 2, 3, 4, 5, 6 et 7
4 Bâche 18 semaines B18 vs 3
vs autres bâches Autres B vs 1, 2, 4, 5, 6 et 7
5 Travail de sol TS vs 4 et 5
vs fauche F vs 6 et 7
6 Travail de sol 6 semaines TS6 vs 4
vs travail de sol 12 semaines TS12 vs 5
7 Fauche 6 semaines F6 vs 6
vs fauche 12 semaines F12 vs 7
8 Bâche 6 semaines installée au printemps 6P vs 1
vs 6 semaines installée l’été 6A vs 5 et 7
9 Bâche 12 semaines installée au printemps 12P vs 2
vs 12 semaines installée l’été 12A vs 4 et 6
1
: Voir les tableaux 1 et 2 pour la description détaillée des traitements et de leurs abréviations.
Les pourcentages de la population initiale de chiendent ont été calculés en utilisant la formule :
𝑃 =𝐷1− 𝐷0 𝐷0
× 100
où :
P = Pourcentage de la population initiale
D0 = Biomasse initiale de rhizomes ou nombre initial de bourgeons (T0)
D1 = Biomasse de rhizome ou nombre de bourgeons suite à un échantillonnage (T6,
T12, T18 et T56)
De plus, avec les valeurs de température à la surface du sol mesurées sous les bâches et dans le traitement témoin enherbé, un calcul des degrés-jours a été effectué avec la formule suivante :
26
𝐷𝐽 𝑞𝑢𝑜𝑡𝑖𝑑𝑖𝑒𝑛 = (𝑇𝑚𝑎𝑥+ 𝑇𝑚𝑖𝑛)
2 − 𝑇𝑏𝑎𝑠𝑒
où :
DJ = Degrés-jours
Tmax = Température maximale obtenue dans une journée
Tmin = Température minimale obtenue dans une journée
Tbase = Température de référence (ici nous avons utilisé celle du maïs, soit 6oC)
La somme de tous les degrés-jours pour une saison de croissance a été calculée pour ainsi obtenir un cumul des degrés-jours sous la bâche et dans le traitement témoin enherbé (sans bâche).
27
3. Résultats
3.1 Biomasse sèche des rhizomes de chiendent
Les données de biomasse sèche des rhizomes ont dû subir une transformation racine carrée pour convenir au postulat de l’homogénéité des variances de l’ANOVA. Par contre, toutes les valeurs de biomasse sèche moyenne présentées dans les tableaux et graphiques sont des valeurs non transformées. Les graphiques des résidus sont présentés à l’annexe 3. Les résultats de l’ANOVA sont présentés dans le tableau 5.
Tableau 5. ANOVA pour les données de biomasse sèche des rhizomes du chiendent commun selon les traitements disposés selon un plan en blocs complets aléatoire
Tests des effets fixes de type 3 Source de variation d.l. Numérateur Valeur F Pr > F Traitement 9 4,15 <,0001 Temps 4 44,00 <,0001 Traitement*Temps 36 1,13 0,3058 Site 1 11,48 0,0012 Traitement*Site 9 0,89 0,5401 Temps*Site 1 0,88 0,3513 Trait*Temps*Site 9 0,62 0,7770
Les interactions Traitement*Temps et Traitement*Site ne sont pas significatives. Cela signifie que les biomasses, selon les traitements, ne sont pas différentes en fonction de la date où l’échantillon était pris et du site expérimental à l’étude. Autrement dit, l’effet des traitements n’a pas changé dans le temps ni aux deux sites à l’étude. L’interaction Temps*Site n’est pas significative et nous indique que les biomasses, à chaque moment d’échantillonnage, ne sont pas différentes selon le site à l’étude. Comme aucune interaction n’est significative, nous avons pu les sortir du modèle d’analyse; les degrés de liberté associés à ces effets non significatifs se retrouvent alors au terme d’erreur. La précision du test est ainsi augmentée. Le tableau 6 présente le nouveau tableau de l’ANOVA.
28
Tableau 6. ANOVA pour les données de biomasse sèche des rhizomes du chiendent commun selon les traitements disposés selon un plan en blocs complets aléatoire sans les interactions non significatives
Tests des effets fixes de type 3 Source de variation d.l. Numérateur Valeur F Pr > F Traitement 9 2,42 0,0158 Temps 4 48,70 <,0001 Site 1 10,99 0,0015
Les effets simples nous indiquent que les traitements ont eu un effet significatif sur la biomasse des rhizomes. Les biomasses diffèrent selon les temps d’échantillonnage et diffèrent en moyenne entre les deux sites. Par la suite, les comparaisons simples indiquent quels traitements sont significativement différents entre eux. Dans le tableau 7, seule la comparaison traitement témoin enherbé (TE) vs les traitements de bâches (TB) est significative. En effet, la biomasse sèche des rhizomes de chiendent est plus élevée pour le traitement témoin enherbé que la moyenne des traitements avec une bâche. Toutes les autres comparaisons ne sont pas significativement différentes et donc les traitements utilisant les bâches ne diffèrent pas entre eux. Selon le contraste herbicide glyphosate (GLY) vs les traitements de bâche (TB), il n’y a pas de différence significative entre les biomasses de rhizomes de ces traitements.
29
Tableau 7. Résultat des comparaisons simples pour la biomasse sèche des rhizomes de chiendent (valeurs non transformées)
Contrastes Abréviations1 Biomasse moyenne sèche des rhizomes (g/1000cm3) d.l. Numérateur Valeur F Pr > F 1. GLY vs TB 3,61 3,80 1 0,00 0,9848 2. TE vs TB 6,00 3,80 1 13,58 0,0004 3. TD vs TB 4,42 3,80 1 0,67 0,4145 4. B18 vs Autres B 3,51 3,84 1 0,76 0,3858 5. TS vs Fauche 3,64 4,37 1 2,05 0,1551 6. TS6 vs TS12 3,83 3,45 1 0,05 0,8213 7. F6 vs F12 4,16 4,57 1 0,50 0,4794 8. 6P vs 6A 3,91 4,01 1 0,01 0,9042 9. 12P vs 12A 3,12 4,00 1 3,00 0,0865 1
: Voir le tableau 4 pour la description des comparaisons simples
Plus précisément, les contrastes 1 (Gly vs TB) et 3 (TD vs TB) ne sont pas significatifs. Le traitement de glyphosate et le traitement témoin désherbé à la main ont donc eu un effet similaire aux bâches sur la biomasse moyenne sèche des rhizomes. Le contraste 4 (B18 vs autres B) n’est pas significatif; donc, si on laisse la bâche durant 18 semaines ou moins longtemps, l’effet est le même sur la biomasse des rhizomes de chiendent. Le contraste TS vs Fauche n’est pas significatif; cela signifie que travailler le sol ou faucher la végétation avant de mettre en place une bâche n’offre pas de différence vis-à-vis la biomasse sèche de rhizomes. Par contre, les biomasses moyennes des rhizomes sont légèrement plus élevées pour les traitements de fauche que les traitements de travail de sol. Les contrastes 6 (TS6 vs TS12) et 7 (F6 vs F12) servent à comparer si l’on devait faucher ou travailler le sol à une ou trois reprises pour avoir le plus grand impact sur la biomasse sèche des rhizomes. Comme ces contrastes ne sont pas significatifs, le type et le nombre d’interventions
30
n’affectent pas la biomasse sèche des rhizomes. Les contrastes 6P vs 6A et 12P vs 12A servent à comparer si la bâche doit être installée dès le début de la saison de croissance ou si elle doit être installée au courant de l’été. Comme les deux contrastes ne sont pas significatifs, le moment où la bâche est installée a peu d’impact sur la biomasse sèche des rhizomes. Le tableau 8 vient confirmer ces affirmations. On remarque que les biomasses individuelles de chaque traitement qui a reçu une bâche (1 à 7) ne diffèrent pas.
Tableau 8. Biomasse sèche des rhizomes en fonction des traitements
Traitements1 Biomasse sèche des rhizomes (g/1000cm3) Erreur type 1. B6S12 3,91 0,58 2. B12S6 3,12 0,58 3. B18 3,51 0,58 4. TS6B12 3,83 0,58 5. TS12B6 3,45 0,58 6. F6B12 4,16 0,58 7. F12B6 4,57 0,58 8. Gly 3,61 0,58 9. TE 6,00 0,58 10. TD 4,42 0,58 1
: Voir le tableau 1 pour la description complète des traitements
Pour chaque traitement, il y a 2 échantillons par parcelle répétés à 7 temps d’échantillonnage dans 4 blocs et sur 2 sites, donc n=112.
Tableau 9. Biomasse sèche moyenne des rhizomes en fonction des deux sites étudiés de St-Augustin-de-Desmaures Site Biomasse sèche des rhizomes (g/1000 cm3) Erreur type 1 3,47 0,41 2 4,65 0,38
Au site 1, il y a 2 échantillons par traitement répété pour 10 traitements, répété dans 4 blocs à T0, T18 et T56, donc n=240. Au site 2, il y a 2 échantillons par traitement répété pour 10 traitements, répété dans 4 blocs à T0, T6, T12 et T18, donc n=320.