Extraction de la phosvitine de la granule du jaune d’œuf
par le procédé à hautes pressions hydrostatiques
Mémoire
Pauline Duffuler
Maîtrise en sciences des aliments - avec mémoire
Maître ès sciences (M. Sc.)
Extraction de la phosvitine de la granule du jaune
d’œuf par le procédé à hautes pressions
hydrostatiques
Mémoire
Pauline Duffuler
Sous la direction de :
Alain Doyen, directeur de recherche
Yves Pouliot, codirecteur de recherche
Résumé
La phosvitine du jaune d’œuf (JO) présente des propriétés techno-fonctionnelles d’intérêt. Des études récentes ont montré le transfert de l’acide folique et de la phosvitine de la granule vers le plasma du jaune d’œuf, après un traitement de pressurisation (HPH) (400 MPa/5 min). Ainsi, ce projet vise à optimiser les paramètres de pressurisation afin de déstabiliser la granule du JO et de maximiser l’extraction de la phosvitine. Une granule générée après centrifugation du JO a été pressurisée à 400 et 600 MPa pendant 5 et 10 min. Les granules contrôle et pressurisées ont été centrifugées de nouveau, générant une deuxième granule et un surnageant. Les profils protéiques des fractions ont été obtenus par électrophorèse sur gel de polyacrylamide en mode dénaturé et réduit (SDS-PAGE). L’empreinte massique de la bande correspondant potentiellement à la phosvitine a été analysée par spectrométrie de masse. Le contenu en minéraux et en acide folique a été obtenu par spectrométrie d'émission atomique à plasma à couplage inductif (ICP-OES) et par chromatographie liquide à haute performance (HPLC), respectivement. Les taux de purification et de concentration en phosvitine ont été calculés à partir d’analyses chromatographiques (FPLC). Les résultats générés par SDS-PAGE ont démontré une corrélation entre la sévérité du traitement HPH et l’intensité des bandes de phosvitine observées sur gel pour les plasmas pressurisés. La même tendance a été observée pour la teneur en fer et en phosphore, confirmant l’augmentation de la teneur en phosvitine, cette phosphoprotéine liant le fer dans le JO. Les taux de purification et de concentration maximaux en phosvitine dans le plasma étaient respectivement de 40,05 % et de 33,3 %, après un traitement HPH à 600 MPa pendant 10min. Le mécanisme de transfert de la phosvitine vers le plasma et son potentiel lien avec l’acide folique devront être étudiés dans le cadre de travaux plus fondamentaux.
Abstract
Phosvitin, present in egg yolk, has demonstrated interesting functional and technological properties. Recent studies have showed that high hydrostatic pressure (HHP) induced the transfer of folic acid and phosvitin from the granule to the plasma of egg yolk at 400 MPa for 5 min. This project aims to optimize the pressurization parameters and destabilization of egg yolk granule to maximize the phosvitin extraction. For that purpose, a granule generated after egg yolk centrifugation was firstly pressurized at 400 and 600 MPa during 5 and 10 min. The control and pressure-treated granules were centrifuged to generate a second precipitate and supernatant. The protein profiles of egg yolk fractions were obtained by par sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE). The peptide mass fingerprint of phosvitin bands was obtained by mass spectrometry. The mineral and acid folic content was obtained by induction couples plasma-optic emission spectrofluorimetry (ICP-OES) and high protein liquid chromatography (HPLC), respectively. Finally, the extraction/yield rate was calculated after fast protein liquid chromatography (FPLC) analysis. Results obtained by SDS-PAGE showed that the band intensity corresponding to the phosvitin in pressured-treated plasma fractions increased according to the HHP treatment severity. This was further confirmed by an increase of iron and phosphorus content in these same fractions since phosvitin is an iron-binding phosphoprotein. The highest phosvitin concentration and purity rate were 33.3 % and 40.05 %, respectively at 600 MPa for 10 min. The potential mechanism resulting to the transfer of phosvitin from granule to plasma and the potential interaction of phosvitin with folic acid need further investigations.
Table des matières
Résumé ... iii
Abstract ... iv
Liste des tableaux ... vii
Remerciements ... xi
Avant-propos ... xiii
Introduction ... 1
Chapitre 1: Revue de littérature ... 3
Généralités sur l’œuf ... 4
1. Le jaune d’œuf ... 5
1.1 Généralités sur le jaune d’œuf ... 5
1.2 Composition protéique de la granule et du plasma du jaune d’œuf ... 6
1.2.1 Le plasma... 7
1.2.2 La granule ... 7
2. La phosvitine ... 8
2.1 Structure et composition ... 9
2.2 Propriétés biologiques et physico-chimiques de la phosvitine et de ses dérivés peptidiques 11 2.2.1 Propriétés techno-fonctionnelles et biologiques de la phosvitine et de ses dérivés peptidiques ... 11
2.3 Les méthodes d’extraction et de purification de la phosvitine ... 17
3. Les hautes pressions hydrostatiques ... 18
3.1 Principe et fonctionnement ... 19
3.2 Avantages et inconvénients ... 21
3.2.1 Avantages ... 21
3.2.2 Limitations ... 21
3.3 Impact sur les protéines ... 22
3.4 L’utilisation des HPH pour la production de fractions enrichies en molécules d’intérêt . 23 3.4.1 Utilisation des HPH pour l’extraction des constituants du jaune d’œuf ... 25
3.4.2 Utilisation des hautes pressions hydrostatiques pour l’extraction de la phosvitine: résultats préliminaires ... 26
Chapitre 2 : Hypothèse et objectifs ... 28
1. Hypothèse ... 29
Chapitre 3: High hydrostatic pressure induced extraction and selective transfer of phosvitin from the
egg yolk granule to plasma fractions ... 30
Résumé ... 31
Abstract ... 32
1. Introduction ... 33
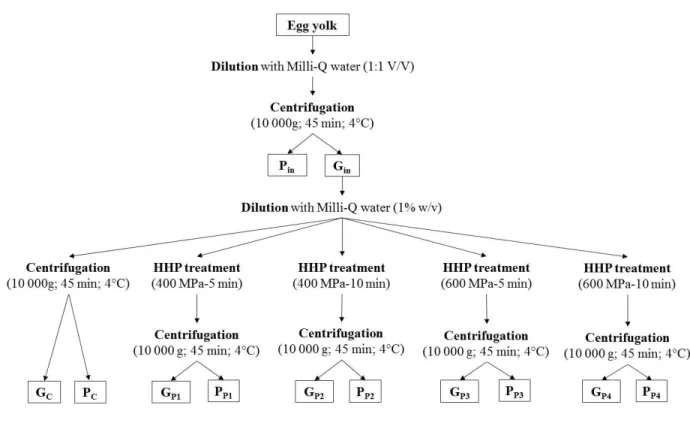
2. Materials and methods ... 35
2.1 Preparation of granules ... 35
2.2 High hydrostatic pressure treatment ... 35
2.3 Analysis ... 36
2.3.1 Protein profiles of egg yolk fractions by SDS-PAGE ... 36
2.3.2 LC–MS/MS analysis ... 37
2.3.3 Phosphorus and iron content in granule and plasma fractions ... 38
2.3.4 Fast protein liquid chromatography ... 38
2.3.5 Folic acid analysis ... 39
2.3.6 Statistical analysis ... 39
3. Results and discussion ... 39
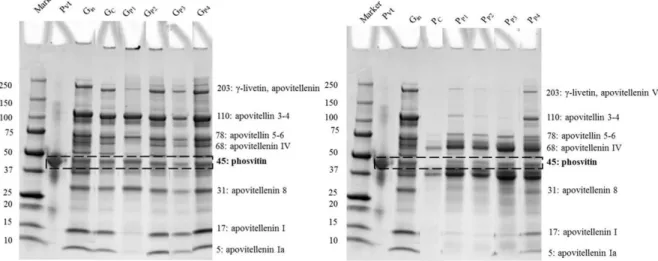
3.1 Impact of HHP treatment on protein profiles of granule and plasma fractions ... 39
3.2 Phosvitin identification by LC-MS/MS analysis ... 41
3.3 Mineral composition ... 42
3.4 Concentration and purification rate of phosvitin in plasma fractions ... 44
3.5 Folic acid content in granule and plasma fractions ... 47
4. Conclusion ... 49
Chapitre 4 : Discussion générale, conclusion et perspectives ... 50
Conclusion générale et perspectives ... 53
Liste des tableaux
Tableau 1. Composition de l'α-phosvitine et de la β-phosvitine (Anton et al. 2007). ... 9 Tableau 2. Exemples de l'utilisation des HPH pour la production de fractions enrichies en protéines
d'intérêt ... 24
Table 3. Proteomic analysis of the 45 kDa-band in GP4 and PP4 fractions. ... 42
Table 4. Phosphorus and iron content (g/100 g of dry sample) of control as well as pressure-treated
granule and plasma fractions. ... 44
Table 5. Concentration (mg phosvitin/ 100 mg of dry sample) and purification rate (%) of
phosvitin in control plasma as well as plasma generated from pressure-treated granule ... 46
Liste des figures
Figure 1. Composition des fractions granule et plasma du jaune d'œuf (Mc Cully et al., 1962; Anton
et Gandemer, 1997). ... 6
Figure 2. Les précurseurs des protéines du jaune d'œuf (Chay Pak Ting et al., 2011) ... 7
Figure 3. Représentation schématique de la granule du jaune d'œuf (Naderi et al., 2017a). ... 8
Figure 4. Représentation schématique de la structure de la phosvitine (Anton et al., 2007). ... 10
Figure 5. Système à hautes pressions hydrostatiques commercial (http://www.hiperbaric.com/fr/hautes-pressions) ... 20
Figure 6 : Principe des HPH (http://www.hiperbaric.com/fr/hautes-pressions) ... 20
Figure 7. Profil d'électrophorèse sur gel des protéines de la granule après pressurisation (Std = marqueur ; Gc = granule témoin; PC = Plasma témoin ; GHHP = granule pressurisée; PHHP = plasma pressurisé) (Naderi et al., 2017 a) ... 26
Figure 8. Experimental design for production of granule and plasma fractions ... 36
Figure 9. SDS-PAGE protein profile of granule (A) and plasma (B) fractions. ... 40
Figure 10. Elution profile of plasma fractions generated without (PC) and with (PP1 to PP4) pressurization of granule. ... 45
Liste des abréviations
5-MTHF : 5-methyltetrahydrofolate
CFU : De l’anglais « Colony Forming Unit » EDTA : Acide béthylène diamino tétra-acétique
EPA : Acide eicosapentaénoïque de l’anglais « EicosaPentaénoic acid »
DHA : Acide docosahexaénoïque de l’anglais « DocosaHexanoic acid »
FPLC : De l’anglais « Fast protein liquid chromatography »
G : Granule
HAP : Hydroxyapatite
HDL : Lipoprotéine de haute densité ou de l’anglais « high density
lipoprotein »
HPH / HHP : Haute pression hydrostatique ou de l’anglais « high hydrostatic pressure »
ICP-OES : De l’anglais “Inductively coupled plasma - optical emission
spectrometry”
kDa : KiloDalton
LC-MS/MS : Chromatographie liquide couplée à la spectrométrie de masse
LDL : Lipoprotéine de basse densité ou de l’anglais « low density lipoprotein »
M : Mole
MPa : Mégapascal
MS : Spectrométrie de masse de l’anglais « mass spectrometry »
NaCl : Sulfate d’ammonium (NH4)2SO4 : Ammonium de sulfate
P : Plasma
PAGE : De l’anglais « polyacrylamide gel electrophoresis » ou électrophorèse sur gel de polyacrylamide
pH : Potentiel Hydrogène
PTH : De l’anglais « parathyroid hormone »
SDS : De l’anglais « sodium dodecylsulfate » ou dodécylsulfate de sodium
Remerciements
Je tenais, tout d’abord, à remercier mon directeur de recherche, le Dr. Alain Doyen. Avant tout, je voulais vous remercier de m’avoir permis de réaliser ma maîtrise à l’Université Laval et de m’avoir fait confiance pour réaliser ce projet de recherche sur un sujet novateur, intéressant et stimulant à la fois. Votre disponibilité, vos précieux conseils et vos encouragements m’ont permis d’acquérir des connaissances et des compétences solides pour la poursuite de mes études et ma future carrière. Merci de m’avoir si bien accueillie dans votre équipe de recherche, une équipe stimulante, où l’entraide et la bonne humeur sont présentes quotidiennement. Ces deux dernières années furent enrichissantes autant d’un point de vue personnel que professionnel. Cette expérience m’a confortée dans l’idée de poursuivre mes études en doctorat ainsi que de m’orienter dans le domaine des Sciences des Aliments. Je voudrais également remercier mon co-directeur Dr. Yves Pouliot pour son soutien et sa bienveillance tout au long de ces deux années.
Je remercie également les évaluateurs de ce mémoire d’avoir accepté de prendre de leur temps pour me fournir une critique constructive de ce travail afin d’en améliorer la qualité. Je tenais également à remercier les Drs Alice Marciniak et Shyam Suwal. Vous avez su m’épauler dans ma réflexion scientifique et m’aiguiller lorsque je faisais face à des obstacles au cours de mon projet de recherche. Vous m’avez tous les deux transmis votre passion pour la recherche et je vous en remercie infiniment. C’est certainement grâce à vous que je souhaite continuer en doctorat aujourd’hui.
Au Dre Nassim Naderi, je tenais à te remercier pour ton aide et le soutien que tu m’as apporté au début de ce projet. Tes connaissances et ton savoir-faire furent précieux. Comme on le dit si bien en Malaisie et en Indonésie, Terima kasih banyak !
Je voudrais également remercier Mélanie Giarratano, ma partenaire dans ce projet de recherche. Nous nous sommes mutuellement épaulées et soutenues durant cette maîtrise. Ta présence, ta bonne humeur, et ta bienveillance ont illuminé mes journées. Même si nos chemins se séparent, je suis sûre qu’ils se recroiseront, que ce soit à Québec, à la Réunion ou encore à l’autre bout du monde.
Je tenais également à remercier Véronique Perreault pour son aide technique apportée au cours de ma maîtrise. Un grand merci à Diane Gagnon et Sophie Fortin, deux membres exceptionnels du département des sciences des aliments à l’Université Laval. Votre bonne humeur et votre bienveillance m’ont permis de travailler dans un cadre agréable et de m’épanouir quotidiennement dans mes recherches. Votre aide fut précieuse, vous m’avez grandement appris et je vous en suis extrêmement reconnaissante. Un grand merci également à tous les membres de mon équipe de recherche et du département des Sciences des aliments de la faculté.
Ensuite, un grand merci à ma famille et mes amis pour leur soutien. Merci d’être là dans les bons comme dans les mauvais moments.
Finalement, je tenais à remercier la Fédération des producteurs d’œufs du Canada pour le financement de ce projet de recherche.
Avant-propos
Quatre chapitres composent ce mémoire. Les travaux présentés ont été réalisés sous la direction du Dr. Alain Doyen et la co-direction du Dr. Yves Pouliot, deux professeurs du département des Sciences des aliments de l’Université Laval.
Le premier chapitre regroupe l’ensemble de l’état des connaissances actuelles sur l’œuf de poule et son jaune; la phosvitine, ses propriétés biologiques et fonctionnelles, ses méthodes d’extraction et de purification; les hautes pressions hydrostatiques et la problématique associée à ce projet de recherche
Le deuxième chapitre présente l’hypothèse de recherche ainsi que les deux principaux objectifs du projet.
Le troisième chapitre est constitué de l’article scientifique rédigé au cours de la maîtrise dont le titre est « High hydrostatic pressure induced extraction and selective transfer of phosvitin from the egg yolk granule to plasma fractions », sera soumis dans le journal Separation and Purification Technology. Pauline Duffuler est la première auteure de l’article, et a réalisé l’ensemble des expérimentations et des analyses présentées dans ce manuscrit. Les autres auteurs sont Nassim Naderi, Mélanie Giarratano, Shyam Suwal, Alice Marciniak, Guillaume Brisson, James D. House, Yves Pouliot et Alain Doyen qui ont participé à la planification des expériences ainsi qu’à la révision de l’article.
Le quatrième chapitre expose la discussion générale et les conclusions de cette étude. Finalement, les références bibliographiques seront citées à la fin de ce mémoire.
Introduction
La phosvitine du jaune d’œuf est une phosphoprotéine de la granule (fraction insoluble générée après centrifugation du jaune d’œuf), liée aux lipoprotéines de haute densité (HDLs) par des ponts phosphocalciques (Mecham & Olcott, 1949). Ces complexes phosvitine-HDLs sont responsables de la structure de la granule du jaune d’œuf (Burley & Cook, 1961; Powrie & Nakai, 1986; Radomski & Cook, 1964; Saari, Powrie, & Fennema, 1964). Sa richesse en phosphosérines et sa structure linéaire sont deux des particularités de cette protéine (Anton, 2007). De plus, la phosvitine présente un intérêt majeur en raison de ses nombreuses propriétés fonctionnelles et biologiques. En effet, elle possède d’intéressantes propriétés émulsifiantes, antioxydantes, anti-inflammatoires, antibactériennes ainsi qu’une forte capacité de chélation avec le fer, le calcium et le magnésium (Jiang & Mine, 2000; Samaraweera et al., 2014; Sattar Khan et al., 2000; Taborsky, 1963). De ce fait, diverses stratégies d’extraction et de purification de cette protéine d’intérêt ont été proposées. Ces méthodes incluent la précipitation par des sels (Ko, Nam, Jo, Lee, & Ahn, 2011), l’utilisation combinée de dilutions et de traitements thermiques (Ren & Wu, 2015), la chromatographie par échange d’anions (Castellani, Guérin-Dubiard, David-Briand, & Anton, 2004; Lei & Wu, 2012), etc. Ces méthodes possèdent cependant plusieurs désavantages. En effet, certaines utilisent des solvants organiques ou/et des produits chimiques non compatibles avec les standards des industries alimentaire et pharmaceutique comme l’utilisation de l’éthanol, de chlorure de sodium ou encore de sulfate d’ammonium (Ko et al., 2011). D’autres procédés, bien que performants concernant les rendements d’extraction et taux de pureté obtenus, sont longs (plusieurs étapes), coûteux et difficilement applicables à l’échelle industrielle telles que la chromatographie échangeuse d’anions (Castellani, Martinet, David-Briand, Guérin-Dubiard, & Anton, 2003; Lei & Wu, 2012; Ren, Li, Offengenden, & Wu, 2015; Ren & Wu, 2014) et la filtration sur gel (Abe, Itoh, & Adachi, 1982). De ce fait, il devient intéressant de développer de nouvelles stratégies d’extraction de la phosvitine du jaune d’œuf via l’utilisation de procédés efficients et durables.
Le procédé à hautes pressions hydrostatiques (HPH), également appelé pascalisation ou pasteurisation à froid, est une technologie dite novatrice puisque commercialement
disponible seulement depuis les années 1980. Basée sur le principe de Le Châtelier, cette technologie utilise une pression (comprise entre 0,1 MPa et 1000 MPa) transmise instantanément et de façon homogène sur l’ensemble d’un produit (Aertsen, Meersman, Hendrickx, Vogel, & Michiels, 2009; Hugas, Garriga, & Monfort, 2002). Déjà implantée et utilisée en industries alimentaires, cette technologie rapide et efficiente possède de nombreux avantages. En effet, la pressurisation permet généralement de conserver les propriétés organoleptiques des aliments (Martín, Barbosa-Cánovas, & Swanson, 2002; Oey, Lille, Van Loey, & Hendrickx, 2008; Rivalain, Roquain, & Demazeau, 2010) tout en assurant leur innocuité microbienne (Chawla, Patil, & Singh, 2011; Palou, López‐Malo, Barbosa‐Cánovas, Welti‐Chanes, & Swanson, 1999; Patterson, Quinn, Simpson, & Gilmour, 1995; Rivalain et al., 2010). Les HPH sont également utilisées pour extraire une large gamme de molécules d’intérêt (lycopène, polyphénols (anthocyanine, acide phénolique, etc.)) à partir de diverses matrices alimentaires (Cao et al., 2011; Jun, 2006). Récemment, Naderi et al. (2017b) ont étudié l’utilisation combinée des HPH et de la centrifugation pour la génération d’une fraction enrichie en acide folique (5-MTHF) à partir de la granule du jaune d’œuf (Naderi et al. 2017ab). Plus spécifiquement, et après pressurisation et centrifugation de la fraction granule, l’acide folique a été transféré de la granule au plasma (fraction soluble obtenue après centrifugation du jaune d’œuf). Les résultats ont montré qu’une pressurisation à 400 MPa durant 5 min a permis d’extraire 78 % de l’acide folique contenu dans la granule du jaune d’œuf. Fait intéressant, la pressurisation a également engendré le transfert de la phosvitine de la fraction granule vers la fraction plasma (Naderi et al., 2017a). Cependant, il est nécessaire d’optimiser les paramètres du procédé afin de maximiser les taux d’extraction et de purification de la phosvitine. Par conséquent, le but de ce projet de recherche était d’étudier le potentiel de déstabilisation de la granule du jaune d’œuf par les HPH afin de générer une fraction enrichie en phosvitine. Plus spécifiquement, une optimisation des paramètres de pressurisation (pression/durée) sera réalisée afin d’obtenir des rendements d’extraction et des taux de pureté maximaux en phosvitine.
Généralités sur l’œuf
L’œuf de poule est un aliment de base largement consommé dans le monde entier. Au Canada, plus de 800 000 millions d’œufs coquilles furent produits (Gouvernement du Canada, 2017). Dans cette province, 40 % de la production d’œufs est consommée sous forme d’œufs de table et 60 % sous forme d’ovoproduits (blanc, jaune ou œuf entier sous forme liquide, congelée ou en poudre, œufs cuits, etc.) (CNPO Site filière, 2019; Gouvernement du Canada, 2003). Les ovoproduits sont largement produits et utilisés pour leurs propriétés nutritionnelles et fonctionnelles au sein de l’industrie agroalimentaire (pâtisserie, plats préparés, etc.) mais également en industrie pharmaceutique et cosmétique. En effet, l’œuf est reconnu pour ses propriétés stabilisatrices, coagulantes, gélifiantes, texturantes, colorantes et antioxydantes (Burley & Cook, 1961).
L’œuf de poule est composé de plusieurs fractions : l’albumen ou le blanc d’œuf (61,5%), le vitellus ou le jaune d’œuf (29 %), les membranes coquillères et la coquille (9,5%). Chacune de ses parties confère un rôle bien spécifique à l’œuf. La coquille est la couche externe de l’œuf. Cette enveloppe minéralisée, d’environ 0,3 mm d’épaisseur est composée d’une structure ordonnée composée de 95,1 % de minéraux (98,4 % de carbonate de calcium et 1,6 % de carbonate de magnésium et de phosphate de calcium), de 3,3% de matière organique et de 1,6 % d’eau. Sa composition lui confère ses propriétés mécaniques. Grâce à ses nombreux pores, la coquille permet d’assurer les échanges gazeux entre l’embryon et le milieu extérieur, essentiel pour le bon développement de ce dernier. De plus, elle permet également de protéger l’embryon contre les agressions extérieures et les contaminations microbiennes (Anton, 2007). L’albumen, représente les deux tiers du poids de l’œuf. Le blanc d’œuf est défini comme une solution aqueuse de protéines contenant 87 % d’eau, 12,5 % de protéines et moins de 1 % de minéraux. L’ovalbumine est une protéine globulaire qui représente 54 % des protéines du blanc d’œuf. Les autres protéines composant le blanc d’œuf sont principalement l’ovotransferrine (12-13 %), l’ovomucoïde (11%), l’ovalbumine gène Y (7 %) et des lysozymes (3,5 %) (Anton, 2007; Powrie & Nakai, 1986; Stevens, 1991). Les protéines du blanc d’œuf possèdent de nombreuses propriétés nutritionnelles (haute valeur nutritive), techno-fonctionnelles (pouvoir émulsifiant de l’ovalbumine (Mine, Noutomi, & Haga, 1991), pouvoir antibactérien de
l’ovomucine et de l’ovotransferrine (Ibrahim, Sugimoto, & Aoki, 2000; Kobayashi et al., 2004)) et biologiques (protection in vivo de l’embryon contre les contaminations microbiennes) (Stevens, 1991). La troisième partie de l’œuf, le jaune d’œuf, sera présentée en détail dans la section suivante s’agissant du cœur du projet de recherche.
1. Le jaune d’œuf
1.1 Généralités sur le jaune d’œuf
Le jaune d’œuf, ou vitellus, est une matrice entourée d’une membrane appelée vitelline, empêchant le blanc de se mélanger avec le jaune de l’œuf. Le jaune d’œuf apporte l’ensemble des nutriments nécessaires au développement de l’embryon dans l’œuf. Le jaune d’œuf représente 36 % du poids de l’œuf et 50 à 52 % de sa matière sèche. Il est composé principalement de 50 % d’eau, 30,6 % de lipides, 16 % de protéines et 1,7 % minéraux, majoritairement le cuivre, le zinc, le magnésium et le fer. Il contient également en plus faible quantité, des vitamines (A, B1, B2, B9, B5, D, E), des glucides et des acides aminés (Powrie & Nakai, 1986). Le jaune d’œuf contient l’ensemble des lipides de l’œuf ainsi que 78 % de ses calories. Ainsi, celui-ci est constitué de cinq composantes majeures : 68 % de LDLs, 16 % de HDLs, 10 % de livétines, 4 % de phosvitine et de 2 % de protéines mineures. Les lipides et les protéines interagissent entre eux pour former des lipoprotéines de basse densité (LDLs) et des lipoprotéines de haute densité (HDLs). De par sa composition, le jaune d’œuf représente une source intéressante de nutriments pour le consommateur et possède ainsi des qualités nutritionnelles et des bénéfices pour la santé. En effet, les protéines contenues dans le vitellus possèdent une très bonne digestibilité (Nys & Sauveur, 2004). De plus, le jaune d’œuf est une source intéressante d’oméga 3 (acide eicosapentaénoïque (EPA), d’acide docosahexaénoïque (DHA)), de nutriments essentiels au fonctionnement du cerveau et de la vision (Maki et al., 2003). Le jaune d’œuf possède également de nombreuses propriétés antioxydantes et fonctionnelles. En effet, celui-ci est utilisé comme émulsifiant, colorant ou encore agent aromatique (Anton, 2013). Ainsi, les ovoproduits, issus du vitellus, sont beaucoup utilisés en industrie agroalimentaire dans la composition de mayonnaises, de crèmes pour stabiliser les émulsions (Burley & Cook, 1961).
1.2 Composition protéique de la granule et du plasma du jaune d’œuf
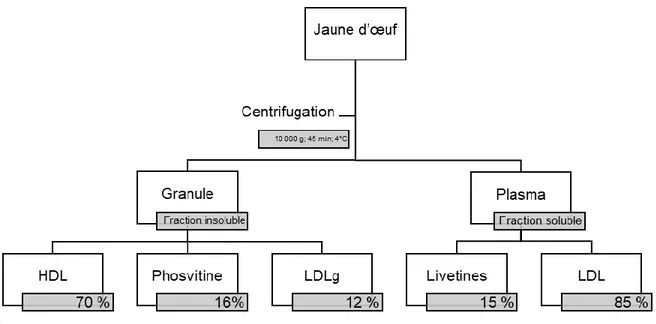
Le jaune d’œuf est une matrice complexe défini comme une émulsion de particules lipidiques, principalement composée de granules suspendues, dispersées dans une solution aqueuse de protéines appelée plasma (Powrie & Nakai, 1986). Ainsi, il est possible de séparer le jaune d’œuf par centrifugation (10 000 g x 45 min) en deux fractions spécifiques : le plasma et la granule (Figure 1) (Causeret, Matringe, & Lorient, 1991) .
Figure 1. Composition des fractions granule et plasma du jaune d'œuf (Mc Cully et al.,
1962; Anton et Gandemer, 1997).
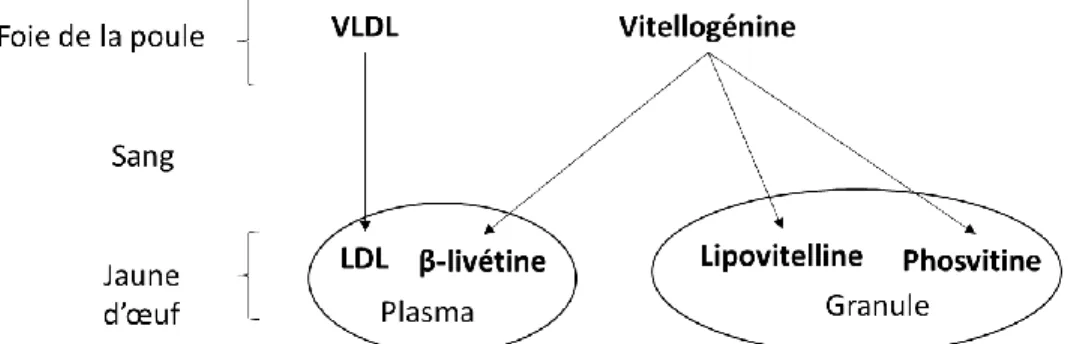
Les protéines du jaune d’œuf sont issues de deux principaux précurseurs : les VLDLs et la vitellogénine. Comme illustré dans la figure 2, ces deux précurseurs sont synthétisés dans le foie de la poule et transportés par le sang dans l’œuf. Les VLDLs sont la source des LDLs (Burley et al., 1984) tandis que la vitellogénine est le précurseur de la β-livétine contenu dans le plasma et des protéines de la granule du jaune d’œuf : la phosvitine et la lipovitélline I et II (Chay Pak Ting et al., 2011; Wallace & Morgan, 1986).
1.2.1 Le plasma
Suite à l’étape de centrifugation, le surnageant, appelé plasma, est défini comme la fraction aqueuse dans laquelle les particules du vitellus sont en suspension. Le plasma représente de 77 à 81 % de la matière sèche du jaune d’œuf. Il est composé de 53 % des protéines et 85 % des phospholipides du jaune d’œuf (Burley & Cook, 1961). De plus, il contient 73 % des lipides et de 25 % des protéines de l’œuf. Plus précisément, les deux composés principaux du plasma sont les LDLs (85 %) et les livétines (15 %). Le plasma contient également la majorité des caroténoïdes responsables de la couleur caractéristique du jaune d’œuf. Les LDLs présents dans cette fraction sont en partie responsables de ses propriétés émulsifiantes (Anton, 2013).
1.2.2 La granule
La granule, qui représente le culot de centrifugation, est la partie solide et non soluble du jaune d’œuf. Elle représente 23 % de la matière sèche du jaune d’œuf. Elle contient 7 % des lipides et 47 % des protéines du jaune d’œuf (Anton, 2013). La granule contient alors 64 % de protéines, 31 % de lipides et 5 % de cendres (Dyer-Hurdon & Nnanna, 1993). Elle est également constituée de HDLs (70 %), de phosvitine (16 %) et de LDLs résiduels (LDLg 12 %). La granule possède également de nombreux minéraux comme le calcium, le fer ou encore le magnésium.
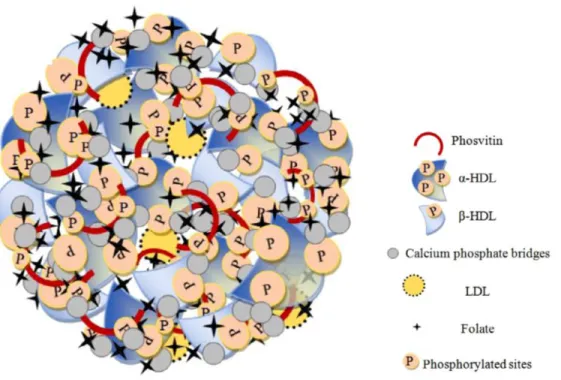
La granule est composée de nombreux complexes sphériques de diamètres compris entre 0,3 et 2,0 µm (Chang, Powrie, & Fennema, 1977). Ces derniers, responsables de la structure du granule, sont composés de phosvitine et HDLs reliés entre eux par des ponts phosphocalciques, entre le groupement phosphate des HDLs et les résidus de phosphosérines de la phosvitine (Figure 3) (Causeret, 1989). Les ponts reliant les groupes phosphates de chacune de ces protéines permettent à la granule de stabiliser sa structure. Ces liaisons sont responsables de la structure très compacte des protéines, peu hydratée et accessible aux enzymes (Causeret et al., 1991).
Figure 3. Représentation schématique de la granule du jaune d'œuf (Naderi et al., 2017a).
2. La phosvitine
La phosvitine est une protéine de la granule du jaune d’œuf. Ses particularités sont à l’origine de son étymologie. La 1ère syllabe « phos » réfère à la forte concentration en phosphore présent dans la protéine tandis que la deuxième syllabe « vitine » fait référence à
son précurseur, la vitellogénine. En effet, la phosvitine, ainsi que les HDLs, sont deux protéines issues d’un précurseur appelé vitellogénine (Taborsky, 1963). Au niveau physiologique, une stimulation d’œstrogène provoque la synthèse de ce précurseur dans le foie des vertébrés. Les HDLs, la phosvitine et d’autres protéines mineures (phosvettes) constituent alors les produits du clivage protéolytique de la vitellogénine (Finn, 2007).
2.1 Structure et composition
La phosvitine représente 3 % de la matière sèche du jaune d’œuf et 11-12 % des protéines totales du jaune d’œuf (Joubert & Cook, 1958). Cette protéine est le deuxième constituant de la granule du jaune d’œuf après les HDLs (Bourgeois-Adragna et al., 1994), avec 16% des protéines du granule du jaune d’œuf (Figure 1) (Powrie & Nakai, 1986).
La phosvitine est une phosphoglycoprotéine (Anton et al., 2007; Mecham & Olcott, 1949) ayant un poids moléculaire variant entre 35 et 45 kDa (Abe et al., 1982). Cette protéine est une association de deux isoformes : l’α-phosvitine et la β-phosvitine (Ito, Fujii, & Yoshioka, 1963). Un agrégat de plusieurs polypeptides (3-4 sous unités de 35 à 45 kDa) forme ces isoformes. La phosvitine β et α représentent 80 % des protéines phosphorylées du jaune d’œuf (Anton et al., 2007; Joubert & Cook, 1958). Chacun de ses isoformes a une composition et des propriétés propres. Les compositions de l’α-phosvitine et de la β-phosvitine sont illustrées dans le Tableau 1.
Tableau 1. Composition de l'α- et de la β-phosvitine (Anton et al. 2007). Poids moléculaire Nombre de sous-unités Composition α-phosvitine 160 kDa 3-4 sous-unités de 35-40 kDa
6 % de glucides (hexose, acide sialique, hexosamine) 3 % de phosphore β-phosvitine 190 kDa 4-5 sous unités de 45 kDa 2 % de glucides (hexoses) 10 % de phosphore
Contenant 10 % de phosphore, la phosvitine est la protéine naturelle la plus phosphorylée tous règnes confondus (Anton et al., 2007; Roach, 1991). L’ensemble du phosphore contenu dans la phosvitine représente 80 % du phosphore total contenu dans le jaune d’œuf (Joubert & Cook, 1958). À l’état natif, la phosvitine est composée de 217 acides aminés
dont 123 sont des sérines (Byrne et al., 1984; Clark, 1985). Parmi l’ensemble des résidus sérines, 90 % sont phosphorylés (Clark, 1985). L’association d’une sérine à un phosphore forme alors les phosphosérines. Le phosphore relie les sérines entre elles par des ponts calcium. La phosvitine peut contenir jusqu’à 15 enchaînements de phosphosérines consécutives, situées au centre de la protéine dans la zone hydrophile. La phosvitine contient également 10 % d’acides aminés hydrophobes (Byrne et al., 1984). Ces acides aminés sont répartis en deux segments hydrophobes situés aux extrémités de la protéine sur les fractions N-terminale et C-terminale (Damodaran & Xu, 1996). Cela confère à la protéine une large zone hydrophile centrale entourée par deux parties hydrophobes (Anton et al., 2007). Les sérines étant des acides aminés chargés négativement, elles interagissent entre elles et se repoussent, provoquant de fortes répulsions électrostatiques entre les phosphosérines. Due à la présence d’une proportion élevée de sérines dans la molécule, ces répulsions limitent le repliement de la protéine ce qui lui confère sa forme linéaire de 28 nm de long et 1,4 nm de large (Figure 4) (Anton, 2007; Joubert & Cook, 1958).
Figure 4. Représentation schématique de la structure de la phosvitine (Anton et al., 2007).
La phosvitine est une protéine résistante à de nombreux traitements physiques et chimiques grâce à sa structure désordonnée et linéaire. En effet, des traitements thermiques sévères (110 °C, 20 min) n’affectent pas sa structure puisqu’une absence de dénaturation et d’agrégation a été observée (Albright, Gordon, & Cotterill, 1984). La phosvitine est également résistante aux traitements de pressurisation hydrostatiques (600 MPa, 10 min) et
aux hydrolyses enzymatiques en raison de ses nombreux sites phosphorylés (Castellani et al., 2004).
2.2 Propriétés biologiques et physico-chimiques de la phosvitine et de ses dérivés peptidiques
La phosvitine possède de nombreuses propriétés techno-fonctionnelles et biologiques telles que des propriétés antioxydantes, antivirales, émulsifiantes, antibactériennes, anticancer ou encore chélatrice d’ions métalliques. Ces propriétés sont développées dans la section suivante.
2.2.1 Propriétés techno-fonctionnelles et biologiques de la phosvitine et de ses dérivés peptidiques
2.2.1.1 Propriétés émulsifiantes
Plusieurs équipes de recherche se sont intéressées aux propriétés émulsifiantes de la phosvitine (Dickinson, Hunt, & Horne, 1992). Cette protéine stabilise également efficacement les émulsions. Ces propriétés fonctionnelles sont dues aux particularités de la phosvitine au niveau de sa structure et de sa composition. En effet, la structure désordonnée de la phosvitine, rendant la protéine flexible et exposant ses zones hydrophobes, facilitent ainsi la formation d’une émulsion. Son efficacité en tant qu’émulsifiant est due à la présence d’acide sialique et de résidus phosphates dans la protéine (Kato, Miyazaki, Kawamoto, & Kobayashi, 1987). De plus, les nombreuses forces électrostatiques répulsives de la phosvitine contribueraient à stabiliser les émulsions (Le Denmat, Anton, & Beaumal, 2000; Petrowski, 1976).
De nombreux facteurs influencent les propriétés émulsifiantes de la phosvitine tels que la concentration en NaCl, le pH, l’état d’agrégation de la protéine, la présence ou non d’autres constituants du jaune d’œuf, etc (Castellani, David-Briand, Guérin-Dubiard, & Anton, 2005; Chung & Ferrier, 1992; Dickinson et al., 1992; Nakamura, Ogawa, Nakai, Kato, &
Kitts, 1998). En effet, une concentration en NaCl supérieure à 0,5 M impacte les forces ioniques et diminue le potentiel émulsifiant ainsi que sa stabilité. De plus, un pH compris entre 3 et 10 augmente le potentiel émulsifiant. Plus précisément, un pH entre 5 et 10 permet une émulsion plus stable (Chung & Ferrier, 1991, 1992). Ainsi, la phosvitine à pH neutre et à des faibles forces ioniques optimisent les propriétés émulsifiantes (Castellani et al., 2005). L’état d’agrégation de la phosvitine a une forte influence sur les propriétés émulsifiantes de la phosvitine. L’agrégation de la phosvitine augmente ses propriétés émulsifiantes mais diminue la stabilité de l’émulsion (Castellani et al., 2005). Par ailleurs, l’ajout de calcium affecte l’agrégation et la stabilité des émulsions. Par exemple, une concentration faible en calcium affecte la floculation (Dickinson et al., 1992). De la même manière, les propriétés émulsifiantes de la phosvitine sont inhibées en présence d’autres constituants de l’œuf tels que les LDLs, les interactions entre les gouttelettes d’huiles et les protéines hydrophobes, comme les LDLs, étant plus fortes qu’avec les protéines hydrophiles comme la phosvitine (Aluko & Mine, 1997; Halling, 1981; Parker, 1987). Le couplage, par la réaction de Maillard du polysaccharide, le galactomannane à la phosvitine améliore significativement les activités émulsifiantes et stabilisantes l’émulsion de cette protéine de l’œuf (Nakamura et al., 1998).
2.2.1.2 Propriétés antioxydantes
La phosvitine est également reconnue pour ses propriétés antioxydantes. En effet, la conformation en brin de la phosvitine et sa richesse en phosphosérines lui permettent de former des interactions avec des cations divalents et trivalents métalliques (Albright et al., 1984; Hegenauer, Saltman, & Nace, 1979). Plus spécifiquement, le calcium, le cobalt, le magnésium, le manganèse ont été particulièrement étudiés pour leur interaction avec la phosvitine (Causeret, Matringe, & Lorient, 1992; Grizzuti & Perlmann, 1973). À titre d’exemple, une molécule de phosvitine est capable de se lier avec 113 ions Mn2+ et avec 120 ions Ca2+ (Anton, 2007). Des ponts ioniques relient ces cations présents dans le jaune d’œuf aux groupes phosphates des phosphosérines de la phosvitine (Causeret et al., 1992; Grizzuti & Perlmann, 1973). Cependant, le cation le plus ciblé et étudié est le fer. En effet, l’œuf contient 2 mg de fer pour 100 g d’œuf. La phosvitine possède une grande affinité avec ce cation divalent puisque 95 % du fer contenu dans le jaune d’œuf est lié à la
phosvitine sous forme de fer ferrique Fe3+ (Albright et al., 1984). Cependant, la phosvitine n’étant pas saturée en fer, d’autres cations s’y fixent. Ainsi, à saturation, ce taux de fixation maximal correspond, pour le fer, à 115 mg fixé par gramme de phosvitine. Le ratio fer/phosvitine est alors de 0,5. Due à sa forte affinité, il est difficile de libérer le fer une fois celui-ci fixé sur la phosvitine (Albright et al., 1984). En effet, un traitement thermique (110 °C durant 20 et 40 min) ou une acidification ne permettent pas la séparation de la phosvitine et du fer (Albright et al., 1984; Hegenauer et al., 1979; Taborsky, 1963). Seule l’utilisation de chélatants, tels que l’EDTA, ayant une affinité supérieure à celle de la phosvitine pour le fer permet de dissocier les deux molécules.
La richesse en phosphore de la phosvitine est responsable de ses capacités de chélation avec les métaux. Les chélateurs métalliques représentent une catégorie d’antioxydants. Des études ont démontré que la capacité de chélation de la phosvitine avec les cations est à l’origine de ses propriétés antioxydantes (Lu & Baker, 1986; Sattar Khan et al., 2000). Seul, le fer ou le calcium catalyse l’oxydation des lipides dans des modèles de viandes crues, salées, ou cuites (Decker & Xu, 1998). Cependant, en présence de phosvitine, ces ions métalliques se couplent à la protéine, inhibant ainsi l’oxydation lipidique dans des émulsions de phospholipides (Lu & Baker, 1986). La phosvitine est capable d’inhiber l’oxydation des lipides dans la viande hachée (Lee, Han, & Decker, 2002; Lu & Baker, 1986). Plus spécifiquement, l’ajout de 20 µM de phosvitine dans une viande cuisinée et non salée, réduit de 11 % à 39 % l’apparition d’un des marqueurs de l’oxydation (thiobarbituric acid reactive substances (TBARS)). Lorsque la viande est crue et salée, 60 mM de phosvitine inhibe jusqu’à 20 % cette oxydation (Lee et al., 2002). Les propriétés antioxydantes de la phosvitine sont améliorées lorsque celle-ci est couplée au galactomannane généré lors de la réaction de Maillard. En effet, cela permet de diminuer de 25 % la peroxydation lipidique (dans un système modèle d’acide linoléique) (Nakamura et al., 1998). De plus, la phosvitine pressurisée (600 MPa durant 5 min) est capable de stabiliser l’oxydation des lipides et des protéines dans les produits carnés (Jung, Nam, Ahn, Kim, & Jo, 2013). La phosvitine couplée à un polyphénol, le resveratrol, peut inhiber l’oxydation des lipides dans des émulsions (Duan et al., 2016). L’activité antioxydante de la protéine va être dépendante de deux conditions : le pH et la température. L’activité antioxydante de la phosvitine est maximale à pH neutre et diminue à un pH acide ainsi qu’à
des températures élevées (Lee et al., 2002).La phosvitine ne perd que de 2 à 15 % de ses propriétés antioxydantes quand celle-ci est chauffée à 60 ou 100 °C (Lu & Baker, 1986; Sattar Khan et al., 2000). Cependant, la phosvitine n’est pas encore utilisée en tant qu’antioxydant dans des produits carnés à cause de sa faible teneur dans l’œuf et de son coût de production élevé (Jung et al., 2013).
La phosvitine, seule ou associée à d’autres antioxydants, pourrait être utilisé en tant qu’inhibiteur de l’oxydation lipidique dans des modèles de viande, et jouer ainsi le rôle d’un chélatant naturel pour améliorer la durée de conservation de ces produits.
La phosvitine est également une source de phosphopeptides bioactifs. Le clivage de la phosvitine par une hydrolyse trypsique entraîne la libération de peptides (1-3 kDa), possédant diverses propriétés biologiques et fonctionnelles (Jiang & Mine, 2000; Katayama, Ishikawa, Fan, & Mine, 2007). Ainsi, les propriétés antioxydantes des peptides issus de la phosvitine pourraient être utilisés en industrie agroalimentaire (Katayama et al., 2007). Ajoutée à la formulation de produits alimentaires, des peptides permettraient d’améliorer la qualité du produit, notamment de limiter ses altérations.
2.2.1.3 Propriétés antimicrobiennes
Les peptides bioactifs, issus de la phosvitine, peuvent être utilisés comme agent antimicrobien contre les bactéries gram négatives telle Escherichia coli. Li et al. (2016) ont démontré les propriétés antibactériennes de PT5e, un phosphopeptide dérivé de la phosvitine contre E. coli et Staphylococcus aureus (Li, Wang, Jiang, Cui, & Zhang, 2016). Khan et al. (2000) ont montré qu’E. coli était inhibée en présence 0,1mg/mL de phosvitine lors d’une incubation à 50°C durant 20 min (Sattar Khan et al., 2000). L’activité antimicrobienne de la phosvitine, est due à l’effet combiné 1) de ses propriétés tensioactives, 2) de son pouvoir de chélation avec les ions métalliques et 3) du stress thermique appliqué. En effet, un stress thermique affaiblirait la paroi cellulaire de la bactérie et déstabiliserait les ions métalliques présents sur sa paroi. Les interactions électrostatiques entre la bactérie et la protéine favoriseraient leur rapprochement. Enfin, les
sites phosphorylés de la phosvitine se lieraient aux ions métalliques de la bactérie provoquant la rupture de la membrane bactérienne et ainsi la mort bactérienne (Sattar Khan et al., 2000). Jung et al. (2013) ont démontré une baisse drastique de la contamination microbienne d’une viande de bœuf hachée contenant de la phosvitine pressurisée à partir de 300 MPa. La présence de phosvitine combinée à une pressurisation d’une viande de bœuf hachée à 600 MPa a permis de réduire la contamination microbienne de 7,96 log CFU/g à des niveaux indétectables (<1 log CFU/g) (Jung et al., 2013). Ainsi, la phosvitine et ses phosphopeptides dérivés pourraient potentiellement être utilisés agent antibactérien.
2.2.1.4 Propriétés anticancer
La phosvitine possède des effets potentiellement bénéfiques au niveau de la prévention des cancers de la peau. La mélanine est une protéine synthétisée par la peau, la protégeant notamment des dommages causées par les radiations ultraviolets (Chang, 2012). S’accumulant dans les tissus de la peau, une surproduction de mélanine provoque potentiellement une hyperpigmentation de la peau, l’apparition de tâches de vieillesse, deux dommages associés à l’augmentation du risque d’un cancer de la peau. Des études précédentes ont démontré que plusieurs molécules (la quercétine, l’acide kojique, etc.) qui possèdent des propriétés de chélations avec les métaux, inhibaient l’action de la tyronase, une enzyme à l’origine de la mélanogénèse (Cabanes, Chazarra, & Garcia‐Carmona, 1994; Kubo & Kinst-Hori, 1999). Possédant une forte affinité avec les métaux, l’impact de la phosvitine a été étudié sur l’activité de la tyrosine, et sur la synthèse de mélanine. Selon l’étude de Jung et al. (2012), l’ajout de 50 µg/ml de phosvitine a diminué l’activité de la tyrosinase de 42% et la synthèse de mélanine de 17% en inhibant le facteur de transcription MITF dans les cellules cancéreuses melanoma (B16F10). Ainsi, la phosvitine pourrait potentiellement inhiber la production des pigments, dans les cellules de la peau, notamment, les cellules cancéreuses melanoma. La phosvitine serait alors potentiellement un inhibiteur naturel de la mélanogénèse (Jung et al., 2012; Ren et al., 2015; Samaraweera, 2012; Samaraweera, Zhang, Lee, & Ahn, 2011). L’étude in vitro de Moon et al. (2014) a démontré l’action cytotoxique de la phosvitine sur diverses lignées cellulaires cancéreuses. En effet, la phosvitine aurait des effets protecteurs contre le stress oxydatif induit par les
dommages causés à l’ADN sur les leucocytes humains (Moon et al., 2014). De ce fait, la protéine pourrait trouver des applications dans le secteur pharmaceutique comme agent anticancéreux ou encore dans le secteur cosmétique afin d’inhiber la mélanogénèse et donc de réduire les tâches de la peau dues à la vieillesse.
2.2.1.5 Impact sur le calcium osseux
L’hydrolyse trypsique de la phosvitine génère des peptides ayant la capacité de former des complexes stables avec le calcium. Le calcium est un minéral important dans le fonctionnement de l’organisme puisqu’il participe à de nombreuses fonctions cellulaires (pression sanguine, la fonction neuronales et musculaires) et prévient certaines maladies non transmissibles (cancer colorectal, maladies cardiovasculaires, etc). Des études ont démontré les effets bénéfiques des peptides bioactifs issus de la phosvitine sur la biodisponibilité et l’absorption au niveau osseux du calcium, notamment grâce à leurs propriétés de chélation avec les métaux (Choi, Jung, Choi, Kim, & Ha, 2005; Ren et al., 2015; Samaraweera, 2012; Samaraweera et al., 2011). La minéralisation osseuse est due à la réaction biologique suivante. Lorsque la concentration en calcium est faible dans le sang, l’hormone parathyroide (PTH) est sécrétée entrainant la libération du calcium des os. Cependant, lorsque la concentration en calcium est élevée dans le sang, l’hormone PTH est inhibée et le calcium est alors transformé en hydroxyapatite (HAP), un précurseur de la minéralisation osseuse. Selon Zhang et al. (2015), la conversion du calcium en HAP dans l’intestin est accélérée par un facteur 12, en présence de phosvitine, passant de 6h à 0,5 h (Zhang, Geng, Huang, & Ma, 2015). Choi et al. (2005) ont démontré l’effet bénéfique d’une diète enrichie en phosphopeptides dérivés de la phosvitine sur des jeunes rats. La présence de ses phosphopeptides dans la diète augmentait l’absorption et l’accumulation du calcium dans les os des jeunes rats, comparé à un groupe control. Ainsi, les peptides de la phosvitine amélioreraient la biodisponibilité du calcium et ainsi son incorporation dans les os (Choi et al., 2005).
Les différentes sections précédentes montrent que la phosvitine est une protéine à très haute valeur ajoutée possédant des propriétés fonctionnelles et bioactives d’intérêt pour les secteurs alimentaires, nutraceutiques et pharmaceutiques. De ce fait, plusieurs équipes de recherche se sont intéressées à la mise en place de stratégies d’extraction et de purification de la phosvitine.
2.3 Les méthodes d’extraction et de purification de la phosvitine
Plusieurs recherches ont porté sur l’utilisation de solvants organiques pour l’extraction et la purification de la phosvitine. Ainsi, à partir de la granule du jaune d’œuf, l’utilisation de chlorure de sodium (NaCl 10 % à pH 4) ou du sulfate d’ammonium ((NH4)2SO4 10% à pH 4) ont permis l’extraction de la phosvitine. Plus spécifiquement, un taux de récupération de 97 % de la phosvitine avec un taux de pureté de 85 % ont été obtenu via l’utilisation de NaCl. Pour le sulfate d’ammonium, un rendement de 72 %, avec un taux de pureté de 85 % ont été calculés (Ko et al., 2011; Ren & Wu, 2014). La majorité des méthodes utilisaient également les outils chromatographiques. Castellani et al. (2003) ont étudié et développé deux méthodes afin de purifier la phosvitine : la chromatographie par échange d’anions et la chromatographie par interactions hydrophobes. La chromatographie échangeuse d’anions, séparant les composés selon leurs charges, est le procédé permettant l’obtention des meilleurs rendements d’extraction mais des taux de purification variés. Ainsi, une fraction purifiée en phosvitine a été générée avec un rendement d’extraction de 85 % et un taux de pureté de 98 % (Castellani et al., 2003). Lei & Wu (2012) ont également étudié l’utilisation de la chromatographie par échange d’anions obtenant ainsi un taux de pureté plus élevé, soit 92,6 %, mais un rendement d’extraction faible de seulement 35,4 % (Lei & Wu, 2012).
Hormis la chromatographie, d’autres stratégies telles que la filtration sur gel (Abe et al., 1982), les traitements thermiques (Ren & Wu, 2015), les techniques de filtration baromembranaires (l’ultrafiltration, la diafiltration) (Ting et al., 2011) et la modification du pH (Ren & Wu, 2014), ont été appliquées pour maximiser l’extraction et la concentration de la phosvitine. Ainsi, Ren & Wu (2015) ont étudié l’extraction de la phosvitine par l’utilisation combinée du NaCl (100 g.L−1) et d’un traitement thermique (90°C). Le taux de pureté de la phosvitine dans la fraction générée, a augmenté de 75,7 % à 80,1 % lorsque le
pH testé variait de 8 à 5. La température du traitement thermique impacte également la pureté et le rendement de l’extraction. Ainsi, le taux de pureté de la phosvitine augmente jusqu’à 90 °C puis diminue à 95 °C. L’augmentation de la pureté pourrait être associée à la dénaturation et à la précipitation des contaminants de faibles poids moléculaires présents dans la fraction générée. De ce fait, un traitement thermique à 90 °C combinée à l’utilisation du NaCl a permis d’extraire 58,8 % de la phosvitine et a généré une fraction enrichie à 88 % en phosvitine (Ren & Wu, 2015). Cette méthode est aussi efficace que les techniques précédentes et n’utilise pas de solvants organiques. Cependant, ce procédé utilise une quantité excessive de NaCl. Ren et Wu (2014) ont également étudié l’impact du pH (valeurs de 8 et 5,5) sur l’extraction de la phosvitine à partir de la granule du jaune d’œuf. Le taux de récupération de la phosvitine dans la fraction finale était de 54,5 % (Ren & Wu, 2014). Ting et al. (2011) ont combiné l’ultrafiltration à la diafiltration pour concentrer la phosvitine. La phosvitine était préalablement extraite par précipitation avec le NaCl. L’ultrafiltration a été réalisée via l’utilisation d’une membrane en polyéthersulfone de 10 et 30 kDa. La phosvitine ayant un poids moléculaire de 35 kDa, elle a été concentrée dans le rétentat. Ce procédé a permis une récupération sélective de la phosvitine à 84 % et une maximisation de sa pureté.
Pour l’ensemble des méthodes d’extractions étudiées, les taux d’extraction, les rendements et les taux de pureté obtenus pour la phosvitine sont intéressants. Cependant, plusieurs de ces stratégies (chromatographie, utilisation de NaCl) utilisent des solvants non compatibles avec le domaine alimentaire, utilisent beaucoup d’intrants et génèrent des extrants ayant un impact environnemental significatif. Par conséquent, il est nécessaire de développer des méthodologies innovantes et durables afin de minimiser l’impact environnemental tout en générant des fractions enrichies en phosvitine. La stratégie priorisée dans le cadre de ce projet est l’utilisation des hautes pressions hydrostatiques (HPH).
3. Les hautes pressions hydrostatiques
Le procédé utilisant les HPH, appelé également pasteurisation à froid ou pascalisation, est utilisé principalement pour la conservation des aliments. Cette technologie est considérée
comme émergente et alternative aux procédés de conservation alimentaires conventionnels. Les premières études en laboratoire sur la pressurisation datent de la fin du XIXème siècle, où Hite a proposé d’inactiver les pathogènes du lait par HPH afin de retarder son acidification et ainsi augmenter sa durée de vie. D’abord utilisé dans le secteur de la géologie et de la chimie, l’application des HPH à l’échelle commerciale sur les produits alimentaires ont débuté à la fin des années 1980 au Japon sur des boissons et des confitures. Le procédé de pressurisation s’est par la suite développé dans le monde entier à partir du début des années 1990.
3.1 Principe et fonctionnement
La pressurisation est un procédé basé sur le principe de Le Châtelier (Aertsen et al., 2009) dont la définition est la suivante : si un système chimique en équilibre subit une modification de pression, de température ou encore de concentration, le système évolue vers un nouvel état d'équilibre de manière à faire face aux changements. L’impact de la pression va provoquer la diminution du volume du produit (Lullien-Pellerin & Balny, 2002). Le volume est le principal paramètre affecté par la pression. Les HPH sont régis par trois paramètres distincts :
• Les pressions généralement utilisées varient généralement entre 0,1 (correspondant à la pression atmosphérique) et 1 000 MPa; 600 MPa étant la pression la plus communément appliquée en industries alimentaires (Gonzalez & Barrett, 2010).
• Le temps de maintien à la pression cible dépendra du produit alimentaire initial, de son taux de contamination initial et des applications finales.
• La température est un paramètre optionnel (Koutchma, 2014a).
La pressurisation se déroule selon les étapes suivantes. Le produit, sous forme liquide ou solide, est inséré dans un emballage souple, étanche et pouvant résister à des pressions importantes. Cet emballage est alors scellé hermétiquement. Par la suite, l’aliment à traiter est transféré dans une enceinte close remplie d’eau, transmettant la pression au produit (Figure 5).
Figure 5. Système à hautes pressions hydrostatiques commercial
(http://www.hiperbaric.com/fr/hautes-pressions)
La pression appliquée est isostatique c’est-à-dire qu’elle est appliquée de manière instantanée et homogène en tous points du produit (Hugas et al., 2002). Finalement, une décompression, généralement instantanée est réalisée et le produit est récupéré pour être stocké aux températures de réfrigération (Figure 6).
3.2 Avantages et inconvénients
3.2.1 Avantages
En industries alimentaires, le procédé à HPH est exclusivement utilisé en tant que technologie de conservation des aliments afin d’en assurer l’innocuité par une inactivation des microorganismes pathogènes et d’altération sous forme végétative (Palou et al., 1999; Patterson et al., 1995). Les risques de toxi-infections d’origine alimentaires (toxi-infection, intoxication) sont alors considérablement réduits. La durée de vie des aliments est généralement augmentée (Chawla et al., 2011; Rivalain et al., 2010). Des études ont également démontré qu’un traitement de pressurisation pouvait diminuer l’allergénicité de certains composants (Peñas, Préstamo, Luisa Baeza, Martínez-Molero, & Gomez, 2006). La pressurisation est un procédé préservant globalement les propriétés organoleptiques d’un produit (flaveur, saveur couleur, etc.) et sa valeur nutritionnelle (Oey et al., 2008; Rivalain et al., 2010). Ainsi, les molécules de faibles poids moléculaires comme les vitamines, les arômes volatils restent stables à la pressurisation (Martín et al., 2002). En effet, il a été démontré que la vitamine B (B1, B2, B3, B6) contenue dans un jus d’orange est stable vis-à-vis d’un traitement de pressurisation entre 200 et 500 MPa durant 1 min à température ambiante (Donsi, Ferrari, & Di Matteo, 1996). Néanmoins, cela entraînait une perte mineure en vitamine C. L’acide folique (5-MTHF) contenue dans un jus d’orange est aussi stable face à un traitement de pressurisation (Oey et al., 2008). Les HPH peuvent être aussi utilisées comme un pré-traitement pour améliorer l’extraction de composants bioactifs de certains aliments (Galanakis, 2013; Jun, 2006).
3.2.2 Limitations
Tel que mentionné précédemment, les HPH permettent d’inhiber les pathogènes végétatifs. Cependant, les HPH, aux valeurs de pressions appliquées en industries alimentaires (600 MPa) n’ont pas d’effet sur les spores qui peuvent résister à des pressions extrêmes de 1000 MPa (Garcia-Gonzalez et al., 2007; Rivalain et al., 2010). Cependant, et tout dépendamment du microorganisme à l’étude, l’utilisation combinée des HPH et d’un
traitement thermique permettent une destruction d’une partie des spores (Martín et al., 2002). De plus, un traitement de pressurisation n’est pas efficace sur des produits à faible teneur en eau. L’important coût d’achat de l’équipement est également un frein majeur à la démocratisation du procédé. De plus, les traitements de pressurisation se font par lot (batch), limitant ainsi leurs applications au niveau industriel (Naderi et al., 2017a) . Finalement, bien que le procédé impacte peu les propriétés physico-chimiques/sensoriels des aliments, certaines matrices, en lien avec l’effet des HPH sur les protéines alimentaires, sont relativement sensibles à la pressurisation. À titre d’exemples, la couleur du lait et de la viande crue sont modifiées du fait, respectivement, de la déstabilisation des micelles de caséine et de la dénaturation de la myoglobine induisant une oxydation du fer. La section suivante présente plus en détail l’impact des HPH sur les protéines.
3.3 Impact sur les protéines
La pressurisation affecte la structure et la conformation des protéines (Silva, Foguel, & Royer, 2001). Ainsi, les liaisons covalentes sont généralement peu affectées par les HPH ce qui permet un maintien de la structure primaire de la protéine (Oey et al., 2008). Cependant, les structures secondaires, tertiaires et quaternaires sont impactées par les hautes pressions (Balny & Masson, 1993; Lullien-Pellerin & Balny, 2002; Rivalain et al., 2010; Sancho et al., 1999). Globalement, à partir de 50 MPa, les protéines vont subir des changements de structures réversibles. À partir de 400 MPa, ces changements sont généralement irréversibles (Huppertz, Kelly, & Fox, 2002). Plus spécifiquement, les liaisons ioniques et hydrophobes impliquées dans le maintien de la structure tertiaire et quaternaire des protéines sont déstabilisées par les HPH engendrant la dissociation de la protéine en polymères ou monomères. Concernant les liaisons électrostatiques, les HPH engendrent la dissociation des paires d’ions provoquant le phénomène d’électrostriction, responsable de la diminution de volume des protéines et pouvant induire leur dénaturation (Rivalain et al., 2010). L’impact des HPH sur les liaisons hydrogènes dépend des pressions utilisées. À des pressions faibles (<400 MPa), le nombre de ponts hydrogènes de la protéine augmente. À des pressions plus élevées (>400 MPa), les liaisons hydrogènes sont
dissociées. Si la structure secondaire est affectée, la dénaturation protéique peut être irréversible (Lullien-Pellerin & Balny, 2002; Sancho et al., 1999) .
Affectant la structure de la protéine, un traitement de pressurisation impactera également sa cinétique enzymatique (Dufour, Hervé, & Haertle, 1995). En effet, l’utilisation combinée de la pressurisation et de l’hydrolyse enzymatique augmente le degré d’hydrolyse de l’enzyme et ainsi la quantité de peptides générés sur diverses protéines (ovalbumine, protéines du lactosérum) (Peñas et al., 2006; Quirós, Chichón, Recio, & López-Fandiño, 2007). À titre d’exemple, l’utilisation combinée d’une hydrolyse enzymatique et de la pressurisation sur la β-lactoglobuline (400 MPa; 10 min) permet l’augmentation du rendement peptidique et de la génération de certains peptides bioactifs (VAGTWY, ALPMHIR) (Boukil, 2018). En revanche, la pressurisation peut inactiver également les enzymes dans certains produits alimentaires (jus de fruits) (Bayındırlı, Alpas, Bozoğlu, & Hızal, 2006; Hendrickx, Ludikhuyze, Van den Broeck, & Weemaes, 1998; Sampedro, Geveke, Fan, & Zhang, 2009). En effet, la polyphénol-oxydase (PPO) et la pectine méthyl estérase (PME) ont été inactivées après un traitement HPH (500/600 MPa ; 5-25 min; 25°C) dans un jus de pêche permettant ainsi de retarder les brunissements enzymatiques et d’augmenter la durée de vie du produit (Rao et al., 2014).
3.4 L’utilisation des HPH pour la production de fractions enrichies en molécules d’intérêt
L’impact de la pressurisation a été étudié sur de nombreuses matrices alimentaires. Les HPH, seules ou combinées à d’autres procédés (hydrolyse enzymatique, procédés baromembranaires) ont été utilisés pour améliorer l’extraction des composants bioactifs dans des matrices alimentaires (Galanakis, 2013) et ainsi générer des fractions enrichies en molécules d’intérêt (Tableau 2).
Tableau 2. Exemples de l'utilisation des HPH pour la production de fractions enrichies en
protéines d'intérêt
Études et composés ciblés (en gras) Paramètres de pressurisation
Références
Effects of high hydrostatic pressure on enzymes, phenolic
compounds, anthocyanins, polymeric color and color of
strawberry pulps
400-600 MPa; 5-25 min
Cao et al., 2011
Application of high hydrostatic pressure processing of food to extracting lycopene from tomato paste waste
100–600 MPa; 1-15 min
Jun, 2006
High hydrostatic pressure extraction of flavonoids from propolis
100–600 MPa; 1-10 min
Shouqin, Jun, & Changzheng,
2005 Extraction of anthocyanins from grape skins assisted by high
hydrostatic pressure
200- 600 MPa; 30-90 min;
20-70°C
Corrales, García, Butz, & Tauscher, 2009) High hydrostatic pressure extraction of phenolic compounds
from Maclura pomifera fruit
250-500 MPa, 10 min
Altuner, Işlek, Çeter, & Alpas,
2012 A comparison between high hydrostatic pressure extraction and
heat extraction of ginsenosides from ginseng (Panax ginseng CA Meyer)
80MPa; 12h; 30◦C
H.-S. Lee et al., 2011
Characterization of polyphenols from green tea leaves using a high hydrostatic pressure extraction
100-600 MPa; 1-10 min
Xi et al., 2009
The use of high hydrostatic pressure to modulate milk protein interactions for the production of an alpha-lactalbumin
enriched-fraction
600 MPa/300 s Marciniak,
Suwal, Britten, Pouliot, & Doyen, 2018 High hydrostatic pressure-assisted enzymatic hydrolysis
improved protein digestion of flaxseed protein isolate and generation of peptides with antioxidant activity
100300 MPa; 5 -10 min
Franck et al., 2019
Ultrafiltration performance and recovery of bioactive peptides after fractionation of tryptic hydrolysate generated from
pressure-treated β-lactoglobulin 400-600 MPa; 10 min Boukil, Suwal, Chamberland, Pouliot, & Doyen, 2018
Impact of a high hydrostatic pressure pretreatment on the separation of bioactive peptides from flaxseed protein hydrolysates by electrodialysis with ultrafiltration membranes
400 MPa; 20 min Cecile Urbain Marie et al.,
2019
High hydrostatic pressure effect in extraction of
5-methyltetrahydrofolate (5-MTHF) from egg yolk and granule
fractions
400 MPa; 5 min (Naderi, Pouliot, et al., 2017b)
3.4.1 Utilisation des HPH pour l’extraction des constituants du jaune d’œuf 3.4.1.1 Extraction de l’acide folique (5-MTHF)
L’acide folique (5-methyltetrahydrofolate (5-MTHF)), également appelé folate ou vitamine B9, est présente dans la granule du jaune d’œuf. Cette vitamine a un rôle primordial dans la prévention des maladies cardiovasculaires, des cancers. Chez la femme enceinte, la consommation de folate (400 µg/j) au cours de la grossesse a été corrélée à une baisse du nombre de l’occurrence de spina bifida chez le nouveau-né. Ainsi, plusieurs recherches sont disponibles sur des techniques d’extraction et de purification de l’acide folique, dont celles de Naderi et al. (2014 – 2017 ab). En effet, ces études ont porté sur l’utilisation des HPH comme procédé de déstabilisation du jaune d’œuf et du granule afin de générer une fraction enrichie en acide folique (Naderi et al., 2017 ab) après centrifugation de la fraction pressurisée. Plusieurs paramètres de pressurisation ont été testés soient 200, 400 et 600 MPa pendant 5 et 10 min.
Les résultats ont montré que les HPH engendrent la désintégration de la structure compacte de la granule ainsi que le transfert de l’acide folique de la fraction insoluble, la granule à la fraction soluble, le plasma, permettant la génération d’une fraction enrichie en acide folique. Les paramètres de pressurisation ont également été optimisés afin de maximiser les taux de pureté et de rendement en acide folique. Ainsi, une pressurisation de la granule du jaune d’œuf à 400 MPa pendant 5 min a permis d’extraire 78 % de l’acide folique présent dans la granule. Le taux d’extraction de phosvitine issu de la granule était 5,5 plus important comparé à celui issu du jaune d’œuf. En parallèle à l’extraction de l’acide
folique, Naderi et al. (2017) ont également évalué l’impact des HPH sur les constituants protéiques de la granule et du plasma du jaune d’œuf. Ces résultats sont présentés dans la section suivante.
3.4.2 Utilisation des hautes pressions hydrostatiques pour l’extraction de la phosvitine: résultats préliminaires
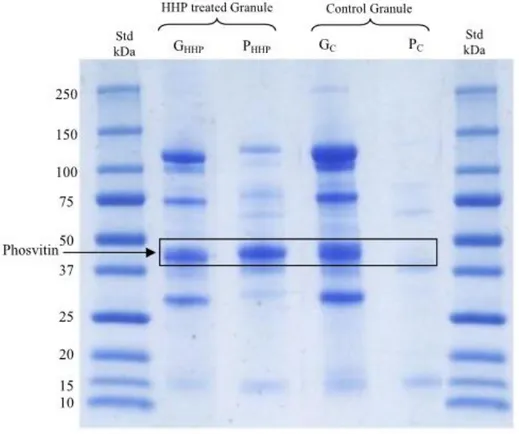
Suite à une précédente étude réalisée sur l’acide folique, Naderi et al. (2017) ont déterminé le profil protéique (SDS-PAGE) de la granule et du plasma obtenus après pressurisation et centrifugation (Figure 7) (Naderi et al., 2017 a).
Figure 7. Profil d'électrophorèse sur gel des protéines de la granule après pressurisation (Std = marqueur ; Gc = granule témoin; PC = Plasma témoin ; GHHP = granule pressurisée; PHHP =
plasma pressurisé) (Naderi et al., 2017 a)
À l’état natif, la phosvitine et l’acide folique sont, tous deux, contenus dans la granule du jaune d’œuf. Cependant, après pressurisation à 400 MPa pendant 5 min, ces deux composés