PUBLICATIONS ET COMMUNICATIONS
Travaux de thèse publiés (revues internationales à comité de lecture) :
Hourcade B, Münz T, Sandoz JC, Rössler W et Devaud JM «Long-term memory leads to synaptic reorganization in the mushroom bodies: a memory trace in the insect
brain?» (2010), Journal of Neurosciences, May 5 ; 30(18) : 6461-5.
Hourcade B, Perisse E, Devaud JM et Sandoz JC «Long-term memory shapes the primary olfactory centre of an insect brain» (2009) Learning & Memory, Sep 30 ; 16(10) : 607-15.
Travaux de thèse présentés lors de congrès scientifiques nationaux et
internationaux :
Hourcade B (septembre 2009) Neural changes associated with long-term memory in the olfactory pathway of Apis mellifera. ApiMondia, Montpellier, France.
Hourcade B, Perisse E , Devaud JM and Sandoz JC (juin 2009) Glomerular plasticity associated with olfactory long-term memory in the honeybee antennal lobe. Workshop Learning and Memory in insects, Roscoff, France.
Hourcade B (juillet 2009) Neural changes associated with long-term memory in the mushroom bodies of Apis mellifera. Atelier de travail organisé à lʼoccasion de la présence exceptionnelle de Martin Heisenberg “From rule learning to gene expression in honeybee”, Toulouse, France.
Hourcade B (mai 2008) Plasticités neurales liées à la mémoire à long terme dans la voie olfactive de lʼabeille. Club de Neurobiologie des Invertébrés, Toulouse, France.
Hourcade B, Perisse E (septembre 2007) Plasticité glomérulaire et mémoire olfactive - Approche structurale et fonctionnelle au niveau du lobe antennaire de l'abeille. Section française de Union internationale pour l'étude des insectes sociaux, Toulouse, France.
Hourcade B, Perisse E, Devaud JM and Sandoz JC (juillet 2007) Glomerular plasticity associated with olfactory long-term memory in the honeybee antennal lobe. The 8th Congress of the international Society for the Neuroethology, Vancouver, Canada.
RESUME
Il est communément accepté que le stockage de mémoires à long terme repose sur des modifications des propriétés fonctionnelles et/ou de la connectivité synaptique du réseau neural. Toutefois, ces remaniements ne sont pas facilement identifiables, étant donné la complexité des procédures dʼapprentissage et des circuits cérébraux étudiés. Une telle recherche peut être facilitée en étudiant la mémoire de stimuli spécifiques dʼune modalité sensorielle, et se concentrant sur des réseaux dʼorganisation modulaire et relativement simple. Nous nous sommes donc attachés à lʼétude de mémoires associatives dʼodeurs uniques, et aux possibles changements neuraux associés, au niveau de deux neuropiles de la voie olfactive de lʼabeille mellifère : les lobes antennaires et les corps pédonculés. Nous avons tiré profit de lʼorganisation modulaire de ces deux centres nerveux pour chercher dʼéventuelles modifications structurales qui pourraient être corrélées avec la formation dʼune mémoire à long terme (MLT) spécifique pour une odeur. Des abeilles en contention étaient entraînées à former une MLT spécifique avec un protocole de conditionnement appétitif pavlovien. La stabilité et la spécificité des performances mnésiques étaient testées comportementalement trois jours après le conditionnement. À ce moment, les cerveaux étaient préparés pour la visualisation des lobes antennaires et des corps pédonculés. Puisque les lobes antennaires sont organisés en unités morphofonctionnelles bien définies, les glomérules, nous avons cherché dʼéventuels remaniements structuraux (par analyse volumétrique) et fonctionnels (par imagerie calcique) au sein de ces unités. Nous avons montré que la MLT est associée avec lʼaugmentation du volume dʼun sous-ensemble spécifique des glomérules, déterminé par lʼodeur apprise, sans modification de la densité des réponses glomérulaires aux odeurs. Puis, nous avons voulu tester la possibilité que la MLT olfactive soit associée avec un remaniement de lʼarchitecture synaptique, de la même manière que chez les mammifères ou dʼautres espèces, chez qui la consolidation dʼune mémoire stable est accompagnée de variations du nombre et de la forme des synapses. Pour cela, nous avons profité de lʼorganisation modulaire des corps pédonculés, où les contacts synaptiques entre lʼafférence olfactive et les cellules intrinsèques sont ségrégés en complexes synaptiques, les microglomérules (MG), qui peuvent être facilement visualisés et dénombrés. Nous avons ainsi montré que la MLT olfactive est associée avec une augmentation de la densité microglomérulaire, dépendante de la transcription. Ensemble, ces résultats fournissent la première preuve de réarrangements structuraux stables et spécifiques du réseau neuronal, incluant la genèse de nouvelles synapses, accompagnant la MLT dans le cerveau dʼun insecte. Nous discutons les mécanismes sous-jacents à de tels changements, suggérant ainsi la conservation, des mammifères aux insectes, des mécanismes impliqués dans le stockage à long terme des mémoires.
ABSTRACT
The storage of stable memories is generally considered to rely on changes in the functional properties and/or the synaptic connectivity of neural networks. However these changes are not easily tractable given the complexity of the learning procedures and brain circuits studied. Such a search can be narrowed down by studying memories of specific stimuli in a given sensory modality, and by working on networks with a modular and relatively simple organization. We have therefore focused on associative memories of individual odors and possibly related changes at two levels of the honeybee olfactory pathway: the antennal lobes (ALs) and the mushroom bodies (MBs). We took advantage of the modular organization of both brain centers to look for structural changes that could be correlated with the formation of an odor-specific, olfactory long-term memory (LTM). Restrained bees were trained to form an odor-specific long-term memory in an appetitive Pavlovian conditioning protocol. The stability and specificity of this memory was tested behaviorally three days after conditioning. At that time, we performed brain processing of ALs and MBs. As ALs are organized in well-identified morpho-functional units, the glomeruli, we looked for evidence of structural (with volumetric analysis) and functional (with calcium imaging) plasticity in these units. We show that olfactory LTM is associated with volume increases in a specific subset of glomeruli, determined by the odor learned, without any change in the density of odor-evoked activity. Then, we addressed the question of whether olfactory LTM could be associated with changes in the synaptic architecture, in a similar way as in mammals and other species, where the consolidation of stable memories is accompanied by structural plasticity involving variations in synapse number and/or size. For this, we took benefit of the modular architecture of the MBs, where synaptic contacts between olfactory input and MB neurons are segregated into discrete synaptic complex(microglomeruli) which can be easily visualized and counted. We thus reveal a transcription-dependent increase in the microglomerular density correlated with the formation of LTM. Taken together, these results provide the first evidence of specific and stable structural rearrangements of neural networks, including the synaptic growth, within the insect brain as specific and stable memories are formed. We discuss the mechanisms underlying such rearrangements, which seem to be a common property of insect and mammalian brain networks involved in the storage of stable memory traces.
Mots clés : plasticité structurale, plasticité synaptique, plasticité fonctionnelle, mémoire à
Table des matières
...
I. INTRODUCTION GENERALE
!
12
...
I.A. La trace mnésique
!
12
... I.A.1) Racines historiques de la notion de trace mnésique! 12
... I.A.2) La consolidation : stabilisation progressive de lʼengramme! 15
... I.A.3) Conception contemporaine de la trace mnésique! 18
... I.A.4) Modifications neurales associées à la mémoire! 19
I.B. L
ʼ
abeille mellifère, modèle insecte de la neurobiologie de la
...
mémoire olfactive
!
25
... I.B.1) Comment étudier lʼapprentissage et la mémoire chez lʼabeille ?! 25
I.B.1)a) Diversité des protocoles
I.B.1)b) Le conditionnement du réflexe dʼextension du proboscis
...
I.B.2) Bases neurales de la mémoire olfactive! 30
I.B.2)a) Architecture fonctionnelle du cerveau de lʼabeille I.B.2)b) Traitement du stimulus olfactif
I.B.2)c) Traitement du stimulus gustatif I.B.2)d) Un minicerveau plastique
...
II. OBJECTIFS DES TRAVAUX DE THESE
!
50
...
III. MATERIELS ET METHODES
!
52
...
III.A. Conditionnement olfactif
!
53
...
III.A.1) Animaux! 53
...
III.A.2) Préparation des abeilles! 54
... III.A.3) Injection de lʼinhibiteur de la transcription! 54
... III.A.4) Procédures de conditionnement et de test de rappel! 56
III.A.4)a) Les stimuli conditionnel et inconditionnel III.A.4)b) Dispositif de stimulation olfactive
III.A.4)c) Phase de conditionnement
III.A.4)d) Soins apportés aux abeilles entre les phases de conditionnement et de test de rappel
III.A.4)e) Phase de test de rappel
...
III.A.5) Réponses mesurées! 63
...
III.A.6) Sélection des animaux! 63
...
III.A.7) Analyse statistique! 63
...
III.B. Analyses anatomiques des neuropiles
!
64
...
III.B.1) Estimation des volumes neuropilaires! 64
III.B.1)a) Étude des volumes glomérulaires III.B.1)b) Etude du volume glial des glomérules III.B.1)c) Etude du volume des lèvres des calices
III.B.2) Estimation de la densité microglomérulaire des calices des corps ...
pédonculés! 71
...
III.B.3) Analyses statistiques! 75
III.B.3)a) Comparaisons des mesures de volume et de densité de MG entre groupes III.B.3)b) Prédiction des modifications glomérulaires
...
III.C. Approche fonctionnelle sur le lobe antennaire
!
77
...
III.C.1) Opération des animaux! 78
...
III.C.2) Sonde fluorescente! 80
...
III.C.3) Procédure dʼenregistrement! 80
...
III.C.4) Stimulations olfactives! 81
... III.C.5) Obtention des cartes de niveaux dʼactivation glomérulaire! 82
...
III.C.6) Cartographie glomérulaire! 83
... III.C.7) Analyse des signaux calciques du lobe antennaire! 83
...
...
IV. RESULTATS
!
87
IV.A. Chapitre 1. Caractérisation comportementale de la mémoire à
...
long terme
!
87
IV.A.1) La mémoire mesurée à 72 heures est associative et spécifique de ...
lʼodeur apprise! 88
IV.A.2) Les abeilles pseudoconditionnées ne développent pas de mémoire ...
associative! 92
IV.A.3) La mémoire mesurée à 72 h est une MLT tardive : elle dépend dʼune ...
vague de transcription! 94
...
IV.A.4) Principales conclusions du chapitre 1! 98
IV.B. Chapitre 2. Plasticité des lobes antennaires en relation avec la
...
MLTt
!
99
IV.B.1) La MLTt est associée à une augmentation du volume de certains ...
glomérules! 99
IV.B.2) La MLTt nʼest pas associée à une modification de la densité des ...
réponses glomérulaires aux odeurs! 106
...
IV.B.3) Principales conclusions du chapitre 2! 114
IV.C. Chapitre 3. Caractérisation des modifications glomérulaires
...
liées à la MLT
!
115
... IV.C.1) La plasticité glomérulaire est indépendante de la transcription! 115
IV.C.2) La plasticité liée à la MLTt affecte autant le coeur et le cortex des ...
glomérules! 124
IV.C.3) La MLTt nʼinduit pas dʼaugmentation du volume de lʼenveloppe gliale ...
des glomérules! 127
IV.C.4) Les modifications des volumes glomérulaires nʼaffectent pas les lobes ...
droit et gauche de la même manière.! 130
IV.C.5) La MLTt nʼaffecte pas la symétrie des patrons dʼactivation glomérulaire ...
! 134
...
IV.C.6) Principales conclusions du chapitre 3! 136
IV.D. Chapitre 4. Plasticité des calices des corps pédonculés en
...
relation avec la MLTt olfactive
!
137
IV.D.1) Lʼarchitecture synaptique des lèvres des corps pédonculés est ...
IV.D.2) Le volume neuropilaire des lèvres des corps pédonculés nʼest pas ...
affecté par la MLTt! 142
... IV.D.3) La plasticité des lèvres liée à la MLTt nʼest pas asymétrique! 144
...
IV.D.4) Principales conclusions du chapitre 4! 146
...
V. DISCUSSION GENERALE
!
147
...
V.A. Principales mises en évidence
!
147
...
V.B. Relations entre modifications neurales et MLTt
!
148
V.B.1) La mémoire mesurée est associative, spécifique de lʼodeur, et ...
dépendante de la transcription! 148
V.B.2) Correspondance entre changements neuraux et MLTt de lʼassociation ...
spécifique SC-SI! 152
V.B.3) Distinction avec les remaniements neuraux liés à lʼâge et à lʼexpérience ...
sensorielle! 159
...
V.C. Origine synaptique de la réorganisation associée à la MLTt
!
161
... V.C.1) Implication potentielle de nouvelles synapses et de la glie! 161
...
V.C.2) Rôle de la signalisation inhibitrice! 165
.... V.C.3) Rôle de la signalisation cholinergique dans les corps pédonculés! 168
V.D. Des mécanismes de plasticité à long terme apparemment
...
fortement conservés
!
169
... V.D.1) Les remaniements structuraux sous-tendent la mémoire! 169
...
V.D.2) La synapse, substrat de la plasticité! 171
...
V.D.3) Rôle peu probable de la neurogenèse! 174
V.D.4) Des sites de potentielle plasticité liée à la mémoire communs entre ...
lobe antennaire et bulbe olfactif ?! 174
... V.D.5) Conservation des bases moléculaires de la plasticité! 175
... V.D.6) Une latéralisation de la trace mnésique chez les insectes ?! 176
...
V.E. Trace mnésique candidate au maintien de la MLTt
!
177
...
V.E.1) Une trace mnésique ?! 177
...
V.E.2) Caractère distribué de la trace! 179
... V.E.3) Mise en place, transformation et devenir de la trace! 180
...
VI. CONCLUSIONS ET PERSPECTIVES
!
182
...
VII. REFERENCES BIBLIOGRAPHIQUES
!
186
...
ABREVIATIONS
AM : Acétoxy-méthyl ActD : actinomycine D CG : cellule gliale CK : cellule de Kenyon CL: calice latéral CM: calice médian CP: corps pédonculéDLT : dépression à long terme IL : interneurone local
LA : lobe antennaire LV: lobe vertical LM: lobe médian
MCT: mémoire à court terme MG : microglomérule
MLT : mémoire à long terme
MLTp : mémoire à long terme précoce
MLTt : mémoire à long terme tardive
NA: nerf antennaire
NP : neurone de projection NRO : neurone récepteur olfactif NV : neurone VUMmx1
PBS : « phosphate buffer salin » PLT : potentialisation à long terme PCL : potentiels de champs locaux REP : réflexe d’extension du probosis SC : stimulus conditionné
SI : stimulus inconditionné TAC: tractus antenno-cérébral
I. INTRODUCTION GENERALE
I.A. La trace mnésique
I.A.1) Racines historiques de la notion de trace mnésique
# Lʼidée selon laquelle les stimuli de notre environnement produisent des traces organiques au sein de notre cerveau, et que ce type de changement est à la base de notre mémoire, a été proposée très précocement. Dès 400 ans avant notre ère, Platon, dans Theaetecus , fait lʼanalogie entre la formation dʼune mémoire et la gravure dʼun texte sur une tablette de cire. Dans cette métaphore, Platon propose que de plus ou moins grandes facultés de mémorisation correspondent à des tablettes de tailles variables. Il illustre aussi lʼoubli par lʼeffacement des caractères gravés dans la cire. Cette notion a été reprise, il y a une centaine dʼannées, par le savant allemand Richard Semon, qui nomma « engramme » (Semon, 1904) cet enregistrement organique que nos tissus extraient des stimuli de notre environnement1. La racine étymologique du terme engramme est grecque, et signifie « quelque chose converti en version écrite ». Semon pensait que le stockage de ces traces dépendantes de lʼexpérience était réalisé dans tous les tissus de lʼorganisme. Dans son ouvrage, The mneme , il énonça deux lois majeures. La première stipulait que les excitations de différents tissus de lʼorganisme constituent un complexe dʼexcitation, un complexe cohérent dʼengrammes interconnectés. La seconde loi précise que lʼengramme doit être « mis en éveil », c.-à-d. quʼil y a nécessité dʼun passage dʼun état de latence à un état dʼactivité (comme nous le verrons plus loin, ces idées font encore écho dans les discussions modernes sur la neurobiologie de la mémoire).
# Toutefois, les minces connaissances anatomiques et les limitations techniques de lʼépoque ont retardé la caractérisation des bases biologiques de ces engrammes, ou traces mnésiques. En revanche, dès le début du 20e siècle, diverses approches psychologiques ont permis, en étudiant les effets de variations de lʼenvironnement sur le
1 Dans la suite du manuscrit, nous utiliserons indifféremment engramme ou trace mnésique, pour désigner les modifications neurales qui permettent de stocker lʼinformation acquise au cours dʼun apprentissage.
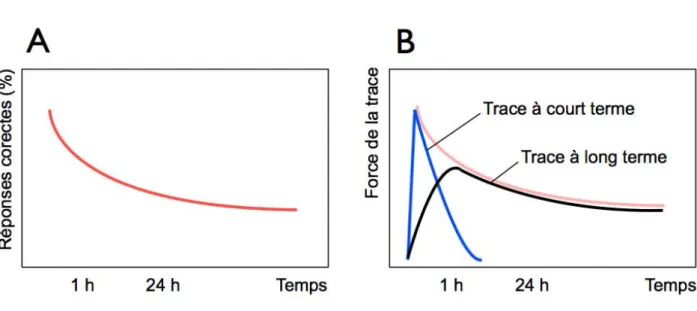
comportement, de mieux comprendre les relations existant entre les traces mnésiques et le comportement. Par exemple, les travaux dʼHermann Ebbinghaus (sur lui-même ; il était à la fois lʼexpérimentateur et son sujet) ont permis de mettre en évidence des caractéristiques majeures du stockage mnésique (Ebbinghaus, 1913). Il montra que les souvenirs ont des durées de vie différentes et que le processus de répétition rend les souvenirs plus durables. En effet, certains souvenirs, de courte durée, sont retenus pendant quelques heures tandis que dʼautres, de longues durées, peuvent persister pendant des jours, des mois, voire des années. Cet auteur a aussi produit la première courbe dʼoubli (fig. 1A). Le rappel était excellent lorsque le test était réalisé juste après lʼapprentissage, mais les performances chutaient de manière importante dans lʼheure qui suivait. Par contre, les performances se stabilisaient par la suite (dès 1 h). Pour expliquer lʼaspect logarithmique de cette courbe, il a été proposé quʼelle résulte de 2 types de trace mnésique (fig. 1B) : une trace à court terme dʼabord très puissante puis qui sʼaffaiblit très rapidement, et une trace à long terme, dʼapparition plus lente, mais plus stable dans le temps.
#
Figure 1 : Courbe d’oubli d’Ebbinghaus et notions de traces à court et long terme. (A) Courbe
d’oubli d’Ebbinghaus. (B) L’existence de deux traces mnésiques distinctes a été proposée, pour expliquer le profil logarithmique de la courbe d’oubli (adapté de Rudy, 2008).
# Rapidement, les travaux dʼautres auteurs ont étayé lʼexistence de ces deux types de traces. Cʼest le cas de lʼétude des pathologies de la mémoire par le psychologue français Théodule Ribot. Il observa que les mémoires les plus récentes étaient les premières à être perdues et que les plus anciennes étaient plus résistantes aux perturbations. Il posa donc une des questions centrales dans lʼétude des mécanismes de la mémoire : qu'est-ce qui rend les souvenirs anciens plus résistants aux perturbations ? Quelques années plus tard, le psychologue allemand Georg Müller et son étudiant Alfons Pilzeckzer suggéraient que cette trace se consolide avec le temps, et ont donc introduit le terme de consolidation mnésique, qui correspond à la période de stockage des informations à long terme (Müller et Pilzecker, 1900).
# La mise en place de protocoles expérimentaux adaptés et contrôlés a permis dʼétudier la dynamique de mémorisation à travers des observations comportementales chez différentes espèces animales. Cʼest notamment le cas du conditionnement associatif classique, qui permet un apprentissage élémentaire simple, robuste et précis, que lʼon peut étudier sur différents modèles animaux, vertébrés ou invertébrés. Ce conditionnement a été mis en évidence et caractérisé pour la première fois par Ivan Pavlov (1927), qui étudiait les réflexes de salivation chez le chien. Ce conditionnement consiste en lʼassociation de deux stimuli : un stimulus initialement neutre, en lʼoccurrence un son, qui devient stimulus conditionnel (SC) au cours du conditionnement, et un stimulus inconditionnel (SI), la nourriture. Le SI se caractérise par sa capacité, avant le conditionnement, à déclencher une réponse réflexe (dans ce cas, la salivation) lors de sa présentation, alors que le son (SC) nʼa pas cet effet initialement. En revanche, si les stimuli conditionnel et inconditionnel sont présentés conjointement et de manière antérograde (de telle sorte que le SC précède le SI), au cours dʼessais successifs de conditionnement, le SC pourra déclencher une réponse comportementale de salivation, appelée réponse conditionnée (RC) en lʼabsence du SI (Pavlov, 1927). À la suite de Pavlov, de nombreux travaux ont permis de caractériser les paramètres du processus de mémorisation chez de nombreuses espèces, et ont abouti à une conception unifiée de ce processus, qui peut être décrit comme la succession de trois étapes clés :
- la première, correspondant lʼapprentissage proprement dit, est la phase dʼacquisition, qui permet la détection et lʼencodage de stimuli informatifs pertinents, et éventuellement lʼintégration dʼinformations sur leurs relations (comme dans le cas du conditionnement classique). Pour reprendre le protocole de Pavlov, cette phase dʼacquisition se déroule lors des présentations successives du SC apparié au SI.
- Ensuite vient la phase de stockage, permettant la conservation de ces informations au sein du système nerveux central, pendant une durée variable en fonction de nombreux paramètres (nature des stimuli et des relations qui les unissent, conditions expérimentales, état de lʼindividu, espèce animale considérée, etc.). Cette phase nʼest généralement pas directement observable au niveau comportemental, mais peut être déduite des effets de perturbations réalisées après lʼacquisition (cf. plus loin).
- Enfin, la phase de rappel correspond à la restitution différée des informations, permettant ainsi lʼexpression dʼun comportement modifié par une expérience antérieure. Selon le type de trace mnésique que lʼon veut évaluer, le test de rappel est réalisé à différents délais. Dans le cadre du conditionnement auditif du réflexe de salivation présenté plus haut, le test de rappel consiste en une présentation du son (SC) seul. Lʼobservation dʼune salivation en réponse au son indiquerait que lʼanimal a acquis, stocké et retenu lʼassociation SC-SI.
I.A.2) La consolidation
: stabilisation progressive de l
ʼ
engramme
# Juste après lʼapprentissage, la mémoire est particulièrement labile et en proie aux interférences : par exemple la présentation de nouveaux stimuli, des stress divers ou les médicaments (Müller et Pilzecker 1900; Lechner et al., 1999 ; McGaugh 2000). Durant cette période initiale du stockage, on assiste à un processus de consolidation, terme qui définit la stabilisation à long terme de la mémoire, à différents niveaux dʼorganisation du cerveau : les consolidations cellulaire et systémique, correspondant respectivement à des changements synaptiques dans un réseau donné et à une réorganisation à plus grande échelle des réseaux abritant la trace (Dudai, 2002). La version cellulaire a été retrouvée
chez tous les modèles animaux étudiés : des vertébrés aux insectes, en passant par les mollusques (Dudai, 2002). Elle est réalisée dans les quelques heures suivant lʼapprentissage (p. ex. Montarolo et al., 1986) et est définie, en pratique, par la fenêtre temporelle pendant laquelle la mémoire peut être altérée par lʼinhibition de la transcription ou de la traduction (ou par une autre interférence : choc électrique, refroidissement...) (pour revue, Davis et Squire, 1984). Les progrès récents de la neurobiologie moléculaire et cellulaire ont permis de comprendre comment lʼapprentissage déclenchait lʼexpression de novo de gènes et la synthèse protéique. Pendant la consolidation cellulaire, lʼactivation de diverses voies de signalisation cellulaire (p. ex. AMPc/PKA, CAMKII ou MAPK) aboutit à des modifications posttraductionnelles de facteurs de transcription (p. ex. CREB ou Elk1). Ces facteurs de transcription activés induisent une ou plusieurs vagues de transcription, et les nouveaux ARN messagers formés pourront participer à la synthèse de nouvelles protéines nécessaires à la stabilisation de la trace. Mais la consolidation ne semble pas se limiter à lʼéchelle cellulaire. Plusieurs travaux, en particulier chez les rongeurs, ont montré une consolidation dite systémique qui lui fait suite, au cours de laquelle la trace dʼune mémoire, initialement stockée dans lʼhippocampe, est transférée au sein des aires corticales (Dudai, 2002). Ce processus de consolidation à grande échelle sʼétale sur des échelles de temps largement supérieures à celles de la consolidation cellulaire, c.-à-d. de quelques heures à quelques années après lʼapprentissage (Bontempi et al., 1999 ; Haist et al., 2001).
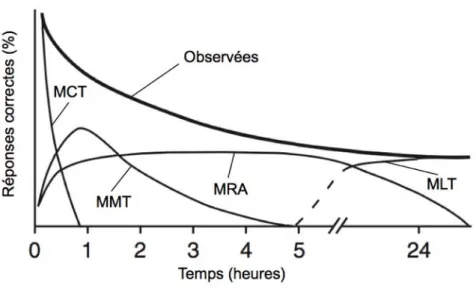
# Les mémoires non consolidée et consolidée2 correspondent aux deux principales phases de mémoire : les mémoires à court et à long terme (MCT et MLT, respectivement). Cette dichotomie a été observée chez tous les animaux étudiés, ainsi que chez lʼhomme. La MCT dure généralement une ou quelques heures, alors que la MLT, dépendante de la synthèse protéique, peut être observée au bout de plusieurs jours, jusquʼà plusieurs années chez certaines espèces (Dudai, 2002). Toutefois, quelques phases de mémoire supplémentaires ont été identifiées chez certaines espèces. Par exemple, chez la drosophile, une dissection génétique à grande échelle a permis de proposer 4 phases fonctionnelles distinctes, permettant dʼexpliquer les performances mnésiques de mouches entraînées à éviter une odeur associée à un choc électrique (fig. 2). Les phases de MCT et de MLT sont bien conservées, mais deux phases supplémentaires ont été identifiées
pour des délais intermédiaires : la mémoire à moyen terme (MMT) et la mémoire résistante à lʼanesthésie (MRA) (Dubnau et Tully, 1998 ; Heisenberg 2003).
Figure 2 : Les phases de la mémoire : exemple de Drosophila melanogaster. Quatre phases
fonctionnelles distinctes ont été proposées, pour expliquer la courbe de mémoire observée chez des mouches soumises à un conditionnement olfactif aversif (MCT : mémoire à court terme ; MMT : mémoire à moyen terme ; MRA : mémoire résistante à l’anesthésie ; MLT : mémoire à long terme). (D’après Dubnau et Tully, 1998 ; Heisenberg 2003.)
I.A.3) Conception contemporaine de la trace mnésique
## Les progrès dans lʼétude de la neurobiologie de la mémoire ont permis de mieux comprendre quels peuvent être les substrats physiologiques de la trace mnésique. Celle-ci est en effet envisagée actuellement comme un ensemble de modifications, plus ou moins stables et durables, survenant dans certains circuits nerveux à la suite dʼun apprentissage. Ces modifications se déroulent à des échelles de temps variables et à de multiples niveaux dʼintégration, comme lʼont montré des études faisant appel à différentes approches : biochimie, biologie moléculaire, électrophysiologie, imagerie fonctionnelle, anatomie, lésions spécifiques, etc. (cf. ci-dessous). Toutes ces modifications constituent des manifestations diverses, dont beaucoup semblent conservées entre espèces, dʼune propriété fondamentale du système nerveux : sa plasticité – à savoir, sa capacité à se modifier de manière adaptée en fonction de lʼexpérience (Dudai, 2002 ; Rudy, 2008).
# Sʼil est généralement admis que ces formes de plasticité sont la condition sine qua non dʼune mémorisation, cette nécessité nʼest en revanche que rarement démontrée du point de vue expérimental. Du point de vue opérationnel, de nombreux auteurs considèrent ainsi comme traces mnésiques des corrélats physiologiques de la manifestation comportementale dʼune mémorisation (Davis, 2005 ; Yu et al., 2006 ; Berry et al., 2008 ; Chen et al., 2009) A cette définition de la trace mnésique, certains opposent une version plus stricte selon laquelle, pour être considérée comme une trace mnésique au sens strict, une manifestation de plasticité doit vérifier les trois critères suivants (Gerber et al., 2004) : elle doit être corrélée avec la mise en place dʼune mémoire, suffisante pour générer cette mémoire et nécessaire à son maintien. Malheureusement, il est souvent très difficile de mettre en évidence ces trois critères du point de vue expérimental (nous reviendrons sur ce point en Discussion).
I.A.4) Modifications neurales associées à la mémoire
# Si lʼon sʼen tient à la caractérisation de la trace mnésique dʼune mémoire à long terme, dʼun point de vue neurobiologique il est généralement admis que la formation et le stockage des souvenirs sʼaccompagnent de modifications durables de lʼefficacité de la transmission synaptique et de lʼarchitecture des réseaux neuronaux activés au cours de lʼapprentissage (consolidation cellulaire), qui peuvent éventuellement sʼétendre secondairement à dʼautres réseaux (consolidation systémique) (pour revues, Bailey et Kandel, 1993 ; McGaugh, 2000 ; Kandel, 2001 ; Dudai, 2004 ; Lynch, 2004). Cette relation entre les processus mnésiques et la plasticité des structures cérébrales était déjà pressentie à la fin du XIXe siècle, par les anatomistes Golgi (1873) et Ramón y Cajal (1894). Ce dernier, qui est à lʼorigine de la « doctrine neuronale », selon laquelle le cerveau est composé de neurones (Ramón y Cajal, 1894-1904), a aussi mis en évidence leurs zones de contact et de communication — les synapses — et a proposé lʼhypothèse de la plasticité synaptique. Cet auteur pensait que les synapses nʼétaient pas des structures fixes, mais quʼau contraire lʼactivité pouvait les modifier sur les plans structural et fonctionnel, et a suggéré ainsi la synapse comme lieu des modifications persistantes nécessaires au maintien de la mémoire. Cette idée est, depuis, toujours restée au cœur de la plupart des recherches sur les mécanismes de la mémoire.
# Un demi-siècle plus tard, Lashley (1950) cherchait à localiser de telles modifications neurales en utilisant la méthodologie de lʼinterférence de la fonction par la dysfonction (Gomulicki, 1953; Herrenstein et Boring, 1965; Brazier, 1988). Cet auteur a infligé des lésions anatomiques sur diverses régions des cerveaux de rats et de singes, et a testé les effets de ces interventions lors de tâches de discrimination et dʼapprentissage en labyrinthe (Lashley 1929, 1950). Toutefois, cette série dʼexpériences fut un échec, et en 1950, il déclarait : ʻ[it] has yielded a good bit of information about what and where the memory trace is not ... I sometimes feel, in reviewing the evidence on the localization of the memory trace, that the necessary conclusion is that learning just is not possible ... Nevertheless, in spite of such evidence against it, learning does sometimes occurʼ (Lashley, 1950). Néanmoins, il fut le premier à combiner des techniques de lésion avec une analyse comportementale. De plus, ses travaux ont suggéré que la trace était dispersée dans plusieurs zones cérébrales, aidant ainsi à lʼélaboration des modèles distribués de la mémoire, par exemple ceux de son étudiant, Hebb.
# Reprenant la première formulation du concept de plasticité neuronale, ce dernier a
décrit un modèle selon lequel lʼactivité électrique qui circule dans les réseaux de neurones
lors de lʼapprentissage se fraye progressivement un chemin en entraînant des
modifications cellulaires ou biochimiques des neurones activés, de sorte que la force des connexions synaptiques entre eux augmente. La plasticité dite hebbienne peut-être
résumée à la célèbre phrase de son auteur : «Cells that fire together wire together» (Hebb,
1949). Autrement dit, il y a nécessité de coactivation contingente des compartiments pré-
et postsynaptiques pour renforcer le lien entre deux neurones (Hebb, 1949). Cʼest plus de
vingt ans plus tard que Bliss et Lomo découvrent dans lʼhippocampe un tel mécanisme de
plasticité synaptique, connu sous le nom de potentialisation à long terme (PLT) (Bliss et Lomo, 1973). La PLT correspond à la propriété des synapses de se renforcer après des
décharges neuronales brèves et intenses. À lʼopposé un mécanisme de dépression à long
terme (DLT) affaiblissant une connexion synaptique a aussi été décrit (Dudek et Bear, 1992 ; Bear, 2003). La PLT est induite par la modification posttraductionnelle de composants essentiels du compartiment postsynaptique (p. ex. les récepteurs AMPA, ou
des effecteurs régulant la densité de récepteurs AMPA), améliorant ainsi lʼefficacité de la
transmission synaptique pendant environ 1 heure. Par contre, si la stimulation est
suffisante la PLT peut persister plusieurs mois (Abraham et al., 2002). Lʼinduction et le
maintien de la PLT, de la même manière que la MCT et MLT, se distinguent par lʼabsence
ou lʼexistence de synthèse de novo de protéines, cʼest-à-dire dʼune consolidation. Lors de
lʼinduction dʼune PLT de longue durée, une cascade dʼactivation est initiée et aboutit à
lʼexpression de gènes (Nguyen et Woo, 2003), dont les produits dʼexpression vont modifier
durablement les synapses stimulées (Rudy 2008). De plus, une traduction locale dʼARN
messagers au niveau des dendrites participe à lʼélaboration de nouvelles protéines
(Sutton et Schuman, 2006), qui peuvent participer à la persistance à long terme du renforcement de la connexion synaptique (plasticité fonctionnelle) et/ou à la modification structurale de la synapse (changement de forme et/ou multiplication).
Plasticité fonctionnelle liée à la mémorisation. Pour permettre un accès expérimental au fonctionnement des réseaux de neurones après lʼapprentissage, diverses approches ont été développées. Par exemple, les techniques dʼélectrophysiologie permettent dʼenregistrer lʼactivité électrique dʼun neurone (si lʼélectrode dʼenregistrement est placée dans le cytoplasme), ou dʼun ensemble de neurones (si lʼélectrode dʼenregistrement est placée dans le milieu extracellulaire). Cette dernière technique permet dʼenregistrer les potentiels de champs locaux (PCL), résultant de lʼactivité électrique des neurones
avoisinant lʼélectrode. À titre dʼexemple, dans le système olfactif des mammifères, lʼenregistrement des PCL de rats vigiles engagés dans une expérience olfactive, a révélé lʼexistence dʼoscillations caractéristiques de lʼodeur présentée (Gervais et al., 2007 pour revue). Ces oscillations se sont révélées modifiables par un apprentissage olfactif (Ravel et al., 2003 ; Martin et al., 2004a,b ; Martin et al., 2006). Dans le premier relais de lʼinformation olfactive, le bulbe olfactif, Ravel et collaborateurs (2003) ont montré que lʼapprentissage olfactif altérait durablement le régime dʼoscillation, notamment dans les fréquences de type gamma (60–90 Hz), qui sont réduites, et bêta (15–40 Hz), qui sont augmentées. Par ailleurs, il a ensuite été démontré que cette plasticité affectant le bulbe olfactif était couplée fonctionnellement aux structures supérieures : notamment les cortex piriforme et entorhinal (Martin et al., 2006) où la modification des oscillations bêta a aussi été détectée (Martin et al., 2004b).
# Une autre approche très utilisée, dans la recherche de corrélats fonctionnels de la mémoire, est la neuroimagerie. Chez lʼhumain, les techniques dʼimagerie par résonance magnétique fonctionnelle (IRMf) et de tomographie par émission de positrons (TEP) ont permis de localiser un grand nombre de régions cérébrales dont lʼactivité était modifiée en parallèle de la formation de différents types de mémoire. Par exemple, une métaanalyse récente, cherchant à faire ressortir les points communs parmi 120 études de neuroimagerie ayant mis en évidence des corrélats fonctionnels disparates de la mémoire sémantique (mémoire des connaissances générales), conclut au rôle de 7 régions cérébrales majeures, incluant certaines zones de lʼhippocampe, du cortex frontal et du lobe pariétal (Blinder et al., 2009). De même, le rappel de mémoires épisodiques (mémoire autobiographique) nécessite la coactivation de nombreuses régions des cortex pariétal et frontal (Rugg et al., 2002).
# Chez les animaux, dʼautres techniques ont été développées afin de visualiser le fonctionnement du cerveau lors de processus dʼapprentissage et de mémoire. Chez la drosophile par exemple, une approche dʼimagerie calcique a permis de mettre en évidence la modification du fonctionnement dʼun neuropile, dʼune population cellulaire, ou même dʼune branche axonale, en relation avec différentes phases de mémoire (cf. plus haut, fig. 2) (Berry et al., 2008 pour revue). À titre dʼexemple, en relation avec le développement dʼune MCT, Yu et collaborateurs (2004) ont détecté des activations supplémentaires de neurones de deuxième ordre de la voie olfactive. De plus, en parallèle de la MLT, la même équipe a mis en évidence une activation accrue dʼune des 2 branches axonales dʼune population cellulaire du centre supérieur majeur du cerveau de la mouche : la branche α des cellules des corps pédonculés (Yu et al., 2006). Ainsi, grâce à
des approches neurofonctionnelles comme lʼélectrophysiologie ou la neuroimagerie, il est possible de révéler des modifications de lʼactivité des réseaux neuronaux en relation avec la formation dʼune trace mnésique.
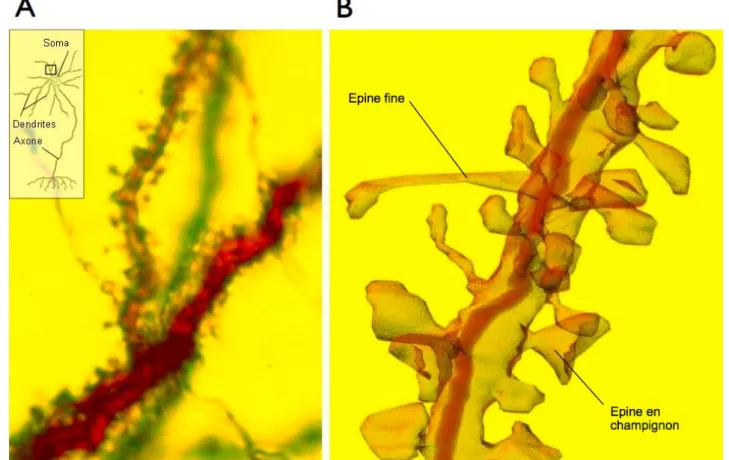
Plasticité structurale liée à la mémorisation. Depuis une trentaine dʼannées, de nombreuses études, alliant protocoles dʼapprentissage et neuroanatomie, tentent de détecter les modifications structurales engendrées par un nouvel apprentissage. Par exemple, Chang et Greenough (1982) ont tiré profit de la séparation des efférences des rétines de rats pour tester lʼeffet dʼun apprentissage visuel latéralisés. Un œil de lʼanimal était couvert afin de disposer dʼun contrôle interne. Les corps genouillés latéraux de lʼhémisphère entraîné ont montré des arbres dendritiques (fig. 3) plus développés que dans lʼhémisphère contrôle. Chez le poussin, Horn et ses collègues ont identifié une région cérébrale qui change de volume et dont la forme des dendrites est modifiée, uniquement au sein de lʼhémisphère gauche, pendant lʼapprentissage particulier de lʼempreinte (Horn et al., 1985). À partir des années 80 également, une série dʼexpériences a montré des modifications dendritiques au niveau du cortex moteur en relation avec des tâches dʼapprentissage faisant intervenir lʼhabileté naturelle des rats à utiliser une de leurs pattes antérieures pour prélever de la nourriture (Greenough et al., 1985 ; Withers et Greenough, 1989 ; Kolb et al., 1997). Ces modifications dendritiques sont depuis peu suivies en direct par microscopie multiphotonique, avant, pendant et après lʼapprentissage ; la croissance de nouvelles épines est observée dans lʼheure suivant lʼapprentissage, et ces nouveaux éléments postsynaptiques persistent à long terme (Xu et al., 2009 ; Ziv et Ahissar, 2009). De plus, il a été montré que le même apprentissage induisait également des formes de PLT (Rioult-Pedotti et al., 2000), suggérant ainsi que le renforcement synaptique consécutif à lʼapprentissage donnait lieu à la formation de nouvelles épines dendritiques, stables.
# En parallèle, de nombreuses études se sont focalisées sur un des centres majeurs de la mémoire chez les mammifères : lʼhippocampe (Leuner et Gould, 2010 pour revue). Une étude marquante a révélé que les chauffeurs de taxi londoniens présentaient de plus gros hippocampes que la moyenne des gens (Maguire et al., 2000), suggérant ainsi que les performances de navigation spatiale développées lors de la pratique de ce métier étaient sous-tendues par une augmentation de la taille du réseau impliqué. Les effets de lʼapprentissage sur lʼarchitecture synaptique de lʼhippocampe incluent des modifications du nombre dʼépines dendritiques et de leur morphologie (Moser et al., 1994 ; OʼMalley et al., 1998, 2000 ; Leuner et al., 2003 ; Knafo et al., 2004) ainsi que de la distribution des
synapses le long des dendrites (Rusakov et al., 1997 ; Andersen & Soleng 1998 ; Geinisman et al., 2001; Eyre et al., 2003 ; Miranda et al., 2006). Cette plasticité est induite par lʼactivité et éventuellement par un mécanisme proche de la PLT. En effet, des études dʼimagerie multiphotonique in vivo ont montré que le développement dʼune PLT était associé à la croissance de nouvelles épines, ainsi quʼà une modification de la forme des épines dendritiques existantes (Lamprecht et LeDoux 2004 ; De Roo et al., 2008 pour revues). Par ailleurs, de nombreux apprentissages régulent la neurogenèse ou la survie des nouveaux neurones du gyrus denté de lʼhippocampe (Trouche et al., 2009 ; Leuner et Gould, 2010 pour revue). Par exemple, lʼapprentissage spatial augmente le nombre de nouvelles cellules du gyrus denté (Ambrogini et al., 2000), et accélère la maturation de leur arbre dendritique (Tronel et al., 2010).
Figure 3 : Les épines dendritiques. (A) Vue au microscope photonique d’une partie de l’arbre
dendritique d’un neurone pyramidal d’hippocampe de rongeur, c’est-à-dire au niveau d’une région spécialisée dans la réception d’afférences de la part d’autres neurones. (B) Reconstruction tridimensionnelle des épines dendritiques d’une portion de dendrite (cadre en (A)), à partir d’images de microscopie électronique à transmission. Une grande diversité d’épines dendritique y apparaît, par exemple de larges épines en champignon, et d’autres, longues et fines. (Crédit image : John C. Fiala, synapse-web.org.)
# Même si la synapse est au centre de la plasticité neurale liée à la mémorisation, quelques données récentes proposent un rôle critique des cellules gliales astrocytaires dans une telle plasticité (Bains et Oliet, 2007 ; Henneberger et al., 2010). Ces travaux indiquent que, pendant le développement de la PLT, les astrocytes proches dʼune synapse activée régulent la densité des récepteurs AMPA de lʼépine dendritique, via la libération de gliotransmetteurs (Bain et Oliet, 2007). Il vient dʼailleurs dʼêtre montré que la libération de D-sérine par les astrocytes était nécessaire à lʼétablissement dʼune PLT à long terme dans la région CA1 de lʼhippocampe (Henneberg et al., 2010). Les cellules gliales semblent donc fortement impliquées dans les phénomènes de plasticité associés à la mémorisation.
# Malgré lʼaccumulation dʼexemples de corrélats de la mémorisation, la plupart de ces études sont limitées par deux principaux aspects. Dʼune part, les protocoles dʼapprentissage utilisés impliquent souvent de former des représentations complexes comprenant de nombreux stimuli (p. ex. apprentissages spatiaux ou contextuels). Une éventuelle modification neurale ne peut donc pas être spécifiquement associée à la mémorisation dʼun stimulus clairement identifié. Dans ce travail de thèse, pour rechercher des modifications neurales liées à la mémoire, nous avons choisi dʼutiliser un conditionnement Pavlovien simple (ou « absolu »), dans lequel un seul stimulus, une molécule odorante, sera associé à une récompense (cf. plus loin, I.B.1)). Dʼautre part, les réseaux neuronaux dans lesquels les modifications liées à la MLT sont recherchées montrent un degré de complexité particulièrement important. Par exemple, lʼhippocampe a été impliqué dans de nombreux types dʼapprentissages (p. ex. le conditionnement de trace, le conditionnement de peur au contexte, la transmission sociale de préférence alimentaire, la navigation spatiale ou la reconnaissance dʼobjet), sans pour autant que lʼon connaisse précisément les sous-circuits hippocampiques impliqués dans chacune de ces tâches. De plus, les circuits sous-tendant une tâche semblent être distribués de manière hétérogène dans les différentes sous-régions de lʼhippocampe (Rolls et Kesner, 2006) et le long de lʼaxe septotemporal (Bannerman et al., 2004). Dans ces conditions, la localisation précise dʼune trace neurale est extrêmement difficile. Pour contourner cet obstacle dans ce travail de thèse, nous avons tiré profit de lʼorganisation relativement simple du cerveau dʼun insecte, lʼabeille mellifère, et avons concentré nos efforts sur des régions cérébrales connues pour traiter spécifiquement les stimuli olfactifs (cf. plus loin, I.B.2)). De plus, lʼorganisation modulaire et la stéréotypie des réponses aux stimuli olfactifs du premier relais synaptique (le lobe antennaire) nous ont permis de tester lʼeffet du type de molécule odorante apprise sur la plasticité potentielle.
I.B. L
ʼ
abeille mellifère, modèle insecte de la
neurobiologie de la mémoire olfactive
I.B.1) Comment étudier l
ʼ
apprentissage et la mémoire chez l
ʼ
abeille
?
I.B.1)a) Diversité des protocoles
# Grâce à un répertoire comportemental riche et varié, lʼabeille sʼest imposée comme un modèle de choix pour lʼétude de lʼapprentissage et de la mémoire (Menzel, 1990, 1999, 2001 ; Giurfa 2007). Depuis les premières expériences de Karl von Frisch sur des abeilles en libre vol entraînées à discriminer des couleurs récompensées par une gouttelette de solution sucrée (von Frisch, 1914), ce sont dʼabord des conditionnements appétitifs qui ont été développés chez lʼabeille, exploitant ainsi le cadre naturel de la récolte de nourriture (nectar) dans les fleurs. Quand une butineuse découvre une source de nectar, elle apprend à associer les caractéristiques de la fleur (visuelles, tactiles et olfactives) au renforcement de nectar (glucides) ou de pollen (protéines) (von Frisch 1967). Les associations réalisées lors du butinage sont étudiées en libre vol, dans des protocoles de conditionnement visuel (Menzel et al., 1968 : Giurfa et al., 1996 ; 2001 ; Benard et Giurfa 2004) ou plus rarement olfactif (Koltermann, 1969 ; Kriston 1971 ; Laska et al., 1999, Laloi et al. 2000). De plus, pour contrôler au maximum le niveau dʼexpérience des animaux, ces procédures ont été adaptées à lʼenceinte du laboratoire en soumettant des abeilles en contention à des conditionnements visuel (Hori et al., 2006 ; 2007 ; Niggebrügge et al., 2009), tactile (Erber et al., 1998 ; Scheiner et al., 2005 ; Giurfa et Malun 2004 ; Dacher et Gauthier, 2008) ou olfactif (Kuwabara, 1957 ; Takeda, 1961 ; Bitterman et al., 1983 ; Menzel et Müller, 1996 ; Menzel, 1990).
# Toutefois, le conditionnement aversif a récemment été introduit chez lʼabeille (Vergoz et al., 2007 ; Carcaud et al., 2009). Il consiste en lʼassociation dʼune stimulation odorante à un léger choc électrique (Vergoz et al., 2007 ; Roussel et al., 2009). La mise au point dʼune telle procédure, sur un principe utilisé depuis longtemps chez la drosophile (Quinn et al., 1974), permet dʼentamer la dissection des mécanismes permettant au cerveau de différencier, traiter et stocker les informations provenant dʼexpériences appétitives et aversives (Schwärzel et al., 2003 ; Roussel et al., 2009).
I.B.1)b) Le conditionnement du réflexe dʼextension du proboscis
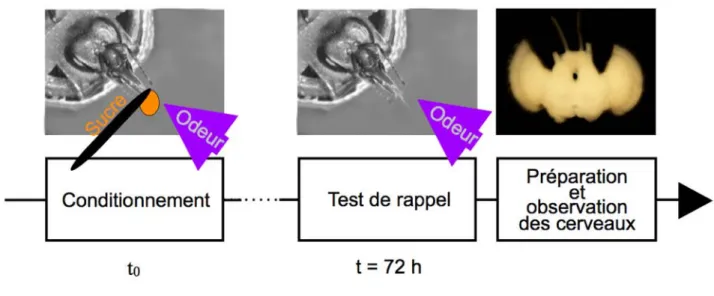
# Comme nous lʼavons vu au paragraphe I.A., le conditionnement de type Pavlovien permet une approche réductionniste des questions touchant aux mécanismes de la mémoire. Parmi les protocoles de conditionnement utilisés chez lʼabeille, le conditionnement olfactif du réflexe dʼextension du proboscis (REP, Takeda et al., 1961 ; Bitterman et al., 1983, fig. 4) est la procédure la plus couramment employée (Menzel 1999 ; Giurfa 2007 ). Il sʼagit dʼun conditionnement appétitif de type Pavlovien, permettant à des animaux en contention dʼapprendre lʼassociation entre une odeur3 neutre (le SC) et une récompense sucrée (le SI). Une des associations que les abeilles réalisent lors du butinage (fig. 4A) — lʼassociation entre lʼarôme et le nectar délivrés par les fleurs butinées — est donc réalisée en conditions contrôlées au laboratoire. Lorsquʼune solution sucrée est appliquée sur les antennes dʼune abeille affamée placée en contention, celle-ci étend son proboscis de façon réflexe (fig. 4B ; Frings, 1944 ; Kuwabara, 1957). Avant le conditionnement, lʼodeur ne déclenche pas (ou peu de) réponse. Au cours du conditionnement, la présentation conjointe des deux stimuli (fig. 4B) selon un appariement précis induit une association entre lʼodeur et la récompense sucrée (cf. Matériels et Méthodes, III.A.4)). Ainsi, lors dʼune présentation ultérieure de lʼodeur (au cours de lʼessai de conditionnement suivant, ou lors dʼun test de rappel) lʼabeille qui a appris lʼassociation répondra par une réponse conditionnée : elle montrera une extension de son proboscis en réponse à lʼodeur (fig. 4C). Cʼest cette réponse conditionnée que nous mesurons lors du test de rappel pour évaluer le niveau de performances mnésiques des abeilles.
3 Conscients du sens du mot «odeur», i.e. la perception par lʼodorat dʼune molécule odorante, nous lʼutilisons dans ce manuscrit pour désigner le stimulus olfactif, par souci de simplicité.
Figure 4 : Le conditionnement du réflexe d’extension du proboscis permet de reproduire
l’association odeur-sucre que l’abeille réalise pendant le butinage. (A) Pendant le prélèvement du nectar, l’abeille perçoit le parfum de la fleur qu’elle butine. Cette association peut être reproduite, dans l’enceinte du laboratoire, lors du conditionnement du réflexe d’extension du proboscis (REP) d’abeilles en contention. (B) La stimulation des récepteurs gustatifs des antennes avec une solution sucrée (SI) déclenche le REP. (C) Pendant le conditionnement, la présentation appariée d’une odeur (SC) et de la solution sucrée (SI) permet à l’abeille d’apprendre l’association SC-SI. (D) Après le conditionnement, et notamment lors du test de rappel, la présentation de l’odeur seule déclenche la réponse conditionnée, c’est-à-dire l’extension conditionnée du proboscis.
# Après un conditionnement du REP en plusieurs essais espacés, le pourcentage dʼindividus présentant des réponses conditionnées reste élevé pendant plusieurs jours, voire plusieurs semaines (Sandoz et al. 1995 ; Menzel, 2001 ; Menzel et al., 2001). Lʼabeille étant en contention, son cerveau est facilement accessible pour la réalisation de manipulations pharmacologiques. La sensibilité des performances mnésiques à différents agents pharmacologiques et à différents délais après le conditionnement a permis de mettre en évidences plusieurs phases de mémoire (Hammer et Menzel, 1995; Menzel, 2001) (fig. 5). Après une phase très précoce de MCT, durant quelques minutes, la consolidation mène à une phase de MTM, durant environ 24 h, et caractérisée par une augmentation de lʼactivité de la protéine kinase C (PKC) dépendante du complexe Ca2+/ calmoduline (Grünbaum et Müller 1998). Ensuite, les performances mnésiques sont sous-tendues par deux phases de MLT : dʼune part une phase précoce de MLT (MLTp) dépendante de la traduction et déclinant vers 48 h, et dʼautre part une phase tardive de MLT (MLTt) dépendante de la transcription et sous-tendant les performances mnésiques les plus tardives, généralement mesurée 72 h ou 96 h après le conditionnement (Grünbaum et Müller, 1998; Wüstenberg et al., 1998; Menzel et al., 1999; Schwärzel et Müller, 2006; Perisse et al., 2009). Pendant le conditionnement, lʼorientation vers une forme de MLT ou lʼautre est fortement influencée par la succession temporelle des essais de conditionnement. Lʼutilisation dʼun protocole en essais massés (avec un intervalle entre essais court, classiquement 1 min) mène principalement à une MLTp, alors quʼun protocole en essais espacés (avec un intervalle plus long entre essais, typiquement entre 5 et 10 min) permet de générer en majorité la MLTt (Menzel et al., 2001 ; Schwärzel et Müller, 2006).
# Comme les autres modèles animaux (cf. I.A.), les abeilles développent dʼabord une MCT labile qui doit être consolidée en MLT pour maintenir les performances mnésiques à long terme. Toutefois, cette espèce présente lʼoriginalité de produire deux types de MLT : la MLTp et la MLTt, dépendantes respectivement de la traduction et de la transcription. Dans ce travail, nous nous concentrerons sur la forme de mémoire la plus classiquement retrouvée chez les autres modèles animaux, et chercherons à mettre en évidence des modifications neuronales stables sous-tendant la MLTt, dépendante de lʼexpression de novo de gènes.
Figure 5 : Phases de mémoire sous-tendant les performances mnésiques induites par le
conditionnement du REP en plusieurs essais espacés. Des essais de conditionnement, multiples et espacés, induisent un niveau important de performances mnésiques, perdurant pendant plusieurs jours (courbe noire). En couleurs sont indiquées les différentes phases putatives de mémoire sous-jacentes aux performances observées à différents temps après l’apprentissage. Après une MCT initiale, la mémoire est consolidée en une MMT, puis en une mémoire à long terme (MLT) précoce, dépendante de la traduction et enfin en MLT tardive, dépendante de la traduction. (Adapté de Sandoz et Pham-Delègue, 2004 ; Schwärzel et Müller, 2006).
I.B.2) Bases neurales de la mémoire olfactive
I.B.2)a) Architecture fonctionnelle du cerveau de lʼabeille
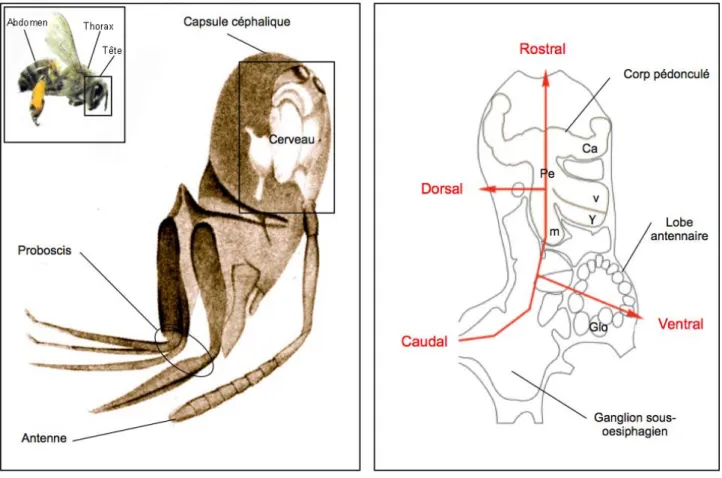
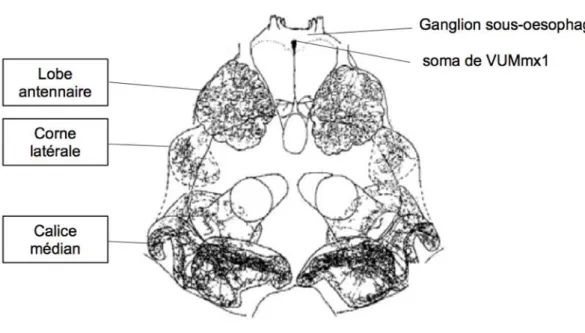
# Le système nerveux des insectes consiste en un cerveau situé dans la capsule céphalique (fig. 6A), et en une chaîne ganglionnaire paire qui sʼétend dans la partie ventrale du thorax et de lʼabdomen. À lʼextrémité antérieure de la chaîne ganglionnaire ventrale, le ganglion sous-œsophagien fait le lien avec le cerveau par le collier périœsophagien (fig. 6B). Le cerveau de lʼabeille, dʼun volume inférieur à 1 mm 3, comprend environ 960 000 neurones (Witthöft, 1967). Un atlas tridimensionnel standard, dorénavant disponible (Brandt et al., 2005 ; www.neurobiologie.fu-berlin.de/beebrain), recense les principaux neuropiles du cerveau de lʼabeille, ainsi que leurs positions et leurs morphologies respectives.
# De nombreuses études ont révélé lʼimportance des neuropiles traitant lʼinformation olfactive — notamment les lobes antennaires et les corps pédonculés (fig. 6B) — dans les processus dʼapprentissage et de mémoire olfactive (Erber, 1976 ; Hammer, 1993 ; Hammer et Menzel, 1995 ; Hammer, 1997 ; Grünbaum et Müller, 1998 ; Hammer et Menzel, 1998 ; Müller, 2002 ; Komischke et al., 2005 ; Devaud et al., 2007). Ce sont également des zones de convergence entre les voies du SC et du SI (voir description détaillée ci-dessous), et par conséquent des régions cérébrales dans lesquelles la plasticité induite par la mémorisation dʼune association odeur-sucre pourrait se produire (Lavond et al., 1993). Cʼest donc dans ces neuropiles que lʼon sʼattachera à caractériser les manifestations de plasticité en lien avec lʼapprentissage et la mémorisation lors dʼun conditionnement associatif appétitif olfactif (cf. I.B.1) ; Menzel, 2001). Avant dʼexaminer lʼétat des connaissances sur ces manifestations, nous examinerons dʼabord le détail des voies de traitement de lʼodeur (SC) et de la solution sucrée (SI), ainsi que leurs relations.
Figure 6 : Situation anatomique du cerveau de l’abeille. (A) Le cerveau de l’abeille est situé
sous la partie rostro-ventrale de la capsule céphalique. (B) Vue en coupe parasagittale du cerveau de l’insecte, où les différents neuropiles et les axes rostro-caudal et dorso-ventral sont représentés . (Adapté de Dujardin, 1850 et Strausfeld, 2002.)
I.B.2)b) Traitement du stimulus olfactif
# Chez lʼinsecte, les molécules odorantes sont détectées au niveau de chaque antenne (fig. 7). Lʼinformation olfactive est ensuite acheminée par le nerf antennaire vers le cerveau, dans un premier relais synaptique, le lobe antennaire (Pareto, 1972 ; Suzuki, 1975). Ensuite, lʼinformation olfactive traitée dans le lobe antennaire est acheminée vers les corps pédonculés et la corne latérale via les tractus antenno-cérébraux (Mobbs, 1982 ; Goodman, 2003).
# Détection des molécules odorantes. Chez lʼabeille, les molécules odorantes sont détectées au niveau des antennes (fig. 6) par des récepteurs olfactifs contenus dans des structures sensorielles spécialisés, les sensilles trichodées et placodées (Esslen et Kaissling, 1976 ; Goodman, 2003; Robertson et Wanner, 2006). La molécule odorante se fixe sur les récepteurs membranaires olfactifs — protéines à 7 domaines transmembranaires, présents à la surface des dendrites des neurones sensoriels olfactifs contenus dans les sensilles — induisant ainsi un potentiel récepteur et éventuellement un train de potentiel dʼaction dans ces neurones (Kaissling, 1987). Lʼinformation olfactive est ensuite acheminée vers le cerveau par le nerf antennaire, constitué des prolongations axonales des quelque 60000 neurones sensoriels, qui fasciculent à lʼintérieur de lʼantenne (Nishino et al., 2009). Le nerf antennaire se projette sur le lobe antennaire ipsilatéral, où il se sépare en 6 tractus sensoriels (Pareto, 1972 ; Suzuki, 1975). Quatre de ces tractus (T1-T4) innervent des parties distinctes du lobe antennaire. Les deux derniers (T5, T6) contournent le lobe antennaire et se projettent vers le lobe dorsal, le ganglion sous-oesophagien et le protocérébron caudal.
Figure 7 : Représentation schématique de la voie olfactive de l’abeille. Les molécules volatiles
sont détectées au niveau des antennes par une population de neurones spécialisés, les neurones sensoriels olfactifs. L’information est ensuite canalisée dans les axones de ces neurones, au niveau du nerf antennaire (NA), qui se termine dans les lobes antennaires (LA). Ce neuropile héberge le premier relais de l’information olfactive, où les dendrites de neurones de deuxième ordre récupèrent l’information, et la transfère via le tractus antenno-cérébral (TAC), jusqu’à la corne latérale (CL) d’une part, et jusqu’aux calices des corps pédonculés (CL et CM : calices latéral et médian). Dans les calices, les dendrites des neurones intrinsèques des corps pédonculés (cellules de Kenyon) collectent l’information et la réorientent, via le pédoncule (Pé), vers les lobes de sortie des corps pédonculés (Lγ : lobe γ , LM : lobe médian, LV : lobe vertical) (CC : corps central ; LO : lobe optique). [À partir d’une représentation du cerveau de l’abeille de Dujardin (1850) et de l’atlas standard 3D (Brandt et al., 2005).]
Traitement de lʼinformation olfactive par les lobes antennaires. Au niveau du cerveau, lʼinformation olfactive est dʼabord traitée par les lobes antennaires, premier relais de lʼinformation olfactive. Cette structure, par son anatomie et sa fonction, est comparable au bulbe olfactif des vertébrés (Hildebrand et Shepherd, 1997 ; Ache et Young, 2005). Elle est composée de structures neuropilaires ovoïdes qui sont autant de sous-unités anatomiques et fonctionnelles : les glomérules. Le nombre de glomérules est beaucoup plus restreint chez les insectes que chez les vertébrés (Ache et Young, 2005). Ainsi, au lieu des 2000 glomérules présents dans le bulbe olfactif du rat, le lobe antennaire de lʼabeille ne compte que 160 glomérules environ (Arnold et al., 1985). De plus, lʼorganisation glomérulaire est stable entre individus. Contrairement aux vertébrés, la plupart des glomérules sont donc identifiables individuellement, en fonction de leur taille, de leur forme et de leur position relative (Flanagan et Mercer, 1989 ; Galizia et al., 1999b). La figure 8A présente lʼatlas glomérulaire de la face rostro-ventrale du lobe antennaire de lʼabeille — c'est-à-dire lʼhémilobe innervé par le tractus antennaire T1 (les glomérules innervés par les tractus T2-4 sont observables sur la face opposée). Cet atlas (Galizia et al., 1999b) sert de référence pour lʼidentification individuelle des glomérules (Galizia et al., 1999b ; Sandoz et al., 2003 ; Deisig et al., 2006, 2010).
#
Figure 8 : Cartographie morphofonctionnelle du lobe antennaire de l’abeille. (A) Atlas morphologique du lobe antennaire de l’abeille en vue rostro-ventrale (Galizia et al., 1999b). D’une abeille à une autre, les glomérules sont retrouvés aux mêmes positions. Ils peuvent donc être facilement identifiés et comparés entre individus. Les glomérules visibles sur cette vue appartiennent au tractus T1. (B) Chaque glomérule, principalement constitué de synapses, et excluant tous corps cellulaires, est entouré d’une couche de cellules gliales (CG). Dans un glomérule particulier, les neurones récepteurs olfactifs (NRO) exprimant un même récepteur membranaire convergent sur les dendrites de deux populations neuronales : les interneurones
locaux (IL) et les neurones de projection (NP). Ces derniers transmettent l’information aux corps pédonculés et à la corne latérale. (C) Atlas physiologique (Sachse et al., 1999). Les profils d’activation glomérulaire sont reproductibles chez un même animal, lors de présentations olfactives successives, mais aussi d’un individu à un autre. Un atlas des activations glomérulaires de la face rostro-ventrale du lobe antennaire a donc pu être réalisé, en réponse à des molécules odorantes de longueurs de chaîne différentes, et comportant des groupements fonctionnels différents.