HAL Id: hal-02596896
https://hal.inrae.fr/hal-02596896
Submitted on 15 May 2020
HAL is a multi-disciplinary open access
archive for the deposit and dissemination of
sci-entific research documents, whether they are
pub-lished or not. The documents may come from
teaching and research institutions in France or
abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est
destinée au dépôt et à la diffusion de documents
scientifiques de niveau recherche, publiés ou non,
émanant des établissements d’enseignement et de
recherche français ou étrangers, des laboratoires
publics ou privés.
To cite this version:
Muriel Gevrey, C. Provost, Christine Argillier. Développement et optimisation des méthodes de
bioindication pour les plans d’eau Indices invertébrés.. [Rapport de recherche] irstea. 2011, pp.38.
�hal-02596896�
Partenariat 2010 – Domaine Qualité des masses d’eau - Action 15 Développement et optimisation des méthodes de bioindication pour les plans d’eau
Développement et optimisation
des méthodes de bioindication
pour les plans d’eau
Indices invertébrés.
Rapport intermédiaire
M. Gevrey, C, Provost et C. Argillier
Contexte de programmation et de réalisation
Le développement des indicateurs de qualité écologique des plans d’eau a été freiné par la faiblesse des connaissances sur le fonctionnement des communautés aquatiques et par l’hétérogénéité des données disponibles qui rendait difficile les approches comparatives. De plus, le faible nombre de masses d’eau de type plans d’eau couplée à leur forte diversité morphologique et environnementale au niveau national est aussi préjudiciable aux approches de modélisation.
Cette action vise le développement de tous les indicateurs biologiques et physico-chimiques soutenant la biologie, nécessaires à l’évaluation de l’état écologique des milieux lacustres, quelle que soit leur origine.
Plus précisément, ce rapport s’intéresse au compartiment de la macrofaune benthique.
Les auteurs
Muriel Gevrey
Ingénieur de Recherche ONEMA
[email protected] Cemagref Aix en Provence Cédric Provost
Ingénieur d’étude
Cemagref Aix en Provence Christine Argillier,
Directeur de Recherche, Pôle plans d’eau,
Cemagref Aix en Provence
Les correspondants
Onema: Yorick Reyjol, DAST, [email protected] Référence du document :
Cemagref : Christine Argillier, Pôle plans d’eau, Cemagref Aix en Provence,
[email protected] Référence du document :
[Autres renseignements nécessaires à la mise sur le Portail « les documents techniques sur l’eau », à renseigner si possible]
Droits d’usage :
Accès libre
Couverture géographique : Niveau géographique [un seul
choix] :
France national
Niveau de lecture [plusieurs
choix possibles] :
Nature de la ressource :
Professionnels, experts Document 2010
Développement et optimisation des méthodes de bioindication pour les plans d’eau
Indices macroinvertébrésRapport 2010
Gevrey, M., Provost, C & C. Argillier
1. Introduction ... 6
2. Propositions de protocoles d’échantillonnage des communautés d’invertébrés benthiques en lacs naturels peu profonds ... 7
2.1. Contexte et objectif... 7
2.2. Site d’étude et méthode... 7
2.2.1. Le site d’étude ... 7
2.2.2. Protocoles d’échantillonnage ... 8
2.2.2.1. Variabilité du cortège faunistique en fonction de la profondeur... 8
2.2.2.2. Variabilité du cortège faunistique en fonction de la distance à la berge ... 9
2.2.2.3. Nature du substrat ... 10
2.2.3. Traitement des données ... 10
2.3. Résultats ... 11
2.4. Conclusions... 16
3. Développement d’outils de bioindication basés sur les communautés d’invertébrés benthiques lacustres... 18
3.1. Métriques adaptées aux plans d’eau artificiels ... 18
3.1.1. Les données disponibles ... 18
3.1.1.1. Description des plans d’eau ... 18
3.1.1.2. Données faunistiques ... 18
3.1.1.3. Paramètres environnementaux naturels ... 19
3.1.1.4. Pressions anthropiques... 20
3.1.1.5. Métriques ... 20
3.1.2. L’approche méthodologique ... 21
3.1.3. Résultats quant à la proposition de métriques candidates pour les plans d’eau artificiels 22 3.1.3.1. Prétraitement des données... 22
3.1.3.2. Première sélection de métriques ... 23
3.1.3.3. Résultats des modèles par métriques ... 24
3.1.4. Conclusions et perspectives... 28
3.2. Métriques adaptées aux plans d’eau naturels profonds... 29
3.2.1. Les données disponibles ... 29
3.2.1.1. Description des plans d’eau et données faunistiques associées ... 29
3.2.1.2. Paramètres environnementaux naturels ... 31
3.2.1.3. Pressions anthropiques... 32
3.2.1.4. Métriques ... 32
3.2.2. L’approche méthodologique ... 33
3.2.3. Résultats : propositions de métriques pour les plans d’eau naturels profonds 33 3.2.3.1. Première sélection de métriques ... 33
3.2.3.2. Résultats des modèles par métriques ... 33
3.2.4. Conclusions et perspectives... 34
4. Conclusion Générale ... 35
Développement et optimisation des méthodes de bioindication pour les plans d’eau
Indices macroinvertébrésRapport 2010
Gevrey, M., Provost, C & C. Argillier
Résumé
Résumé
Ce rapport présente les avancés dans le développement de bioindicateurs basés sur les communautés d’invertébrés benthiques lacustres pour trois types de plans d’eau : i) les naturels peu profonds, ii) les plans d’eau d’origine anthropique et iii) les lacs naturels profonds.
Pour les lacs peu profonds, les premiers essais suggèrent qu’il faut trouver un compromis entre i) l’effort d’échantillonnage qui doit garantir la prise en compte de la variabilité naturelle des substrats dont on a montré l’influence sur les communautés et ii) la qualité de l’échantillon qui doit contenir suffisamment d’individus pour informer de manière fiable sur la diversité, l’abondance et la sensibilité des espèces présentes. Définir des préconisations d’échantillonnage sur la base de ce seul essai serait hasardeux, néanmoins, nous avons montré la stabilité des échantillons de la zone centrale. Ainsi, l’exploitation des informations acquises en zone profonde en application du protocole IOBL est recommandée dans un premier temps. Si ces données ne permettaient pas d’identifier des métriques candidates, des essais complémentaires devraient être envisagés pour définir l’effort d’échantillonnage à appliquer en zone littorale et la localisation des points sur lesquels cet effort doit s’appliquer.
En ce qui concerne les plans d’eau artificiels, à partir de données provenant d’un protocole de prélèvements uniquement d’oligochètes et sur la base d’une approche de modélisation de type hindcasting, des métriques candidates ont été proposées. Le choix de cette approche repose sur le faible nombre de retenues de référence identifiées à ce jour, Quatre métriques semblent mieux répondre à la pression et en particulier la métrique « densité de tubificidés sans soies capillaires », classiquement utilisée comme indicateur de pollution.
Les travaux réalisés sur les plans d’eau naturels profonds laissent penser que des métriques candidates pourront être proposées mais il reste encore un travail important de choix de méthodes d’analyses adaptées du fait de faible nombre de plans d’eau pour lesquels des données sont disponibles. Des métriques candidates devront être à terme proposées et agrégées sous forme d’indice.
Mots clés (thématique et géographique)
Développement et optimisation des méthodes de bioindication pour les plans d’eau
– Indices macroinvertébrésRapport 2010
Gevrey, M., Provost, C & C. Argillier
1. Introduction
En France, 439 plans d’eau, naturels ou artificiels, doivent faire l’objet d’un rendu auprès des autorités européennes (superficie supérieure à 50 hectares). Un bon état général des eaux implique aussi bien un bon état chimique (normes qualité environnementales) qu’un bon état écologique (biologie, physicochimie, hydromorphologie). Pour l’évaluation de l’état écologique des plans d’eau naturels du point de vue de la biologie, seuls les indicateurs de l’élément de qualité phytoplancton avec la concentration en chlorophylle-a et l’indice planctonique IPL (Laplace-Treyture et al., 2009) sont pris en compte actuellement. Des méthodes d’évaluation conformes aux exigences de la directive cadre sur l’eau (Communauté Européenne, 2000) sont encore en phase de développement pour les autres éléments biologiques.
Ce rapport s’intéresse plus particulièrement au développement de bioindicateurs lacustres basés sur la macrofaune benthique. Les macroinvertébrés benthiques sont considérés comme de très bons indicateurs biologiques (Woodcock & Huryn, 2007) et en Europe, ce sont les éléments de qualité biologique les plus utilisés pour révéler les pressions anthropiques (Bailey, Norris, & Reynoldson, 2004; Bonada, Prat, Resh, & Statzner, 2006; Ghetti & Ravera, 1993; Rosenberg & Resh, 1993). Pourtant dans les plans d’eau, alors que ces organismes constituent une biomasse significative dans les lacs et jouent un rôle important dans la production générale de ces milieux (Free et al., 2009; Kagalou, Economidis, Leonardos, & Papaloukas, 2006; Zbikowski & Kobak, 2007), les méthodes de bioindication opérationnelles sont encore rares.
En France, trois systèmes d’évaluation de la qualité des plans d’eau basés sur ces communautés existent, mais aucun n’est utilisé dans le cadre de la DCE. En effet, l’Indice Oligochètes de Bioindication Lacustre (IOBL) (AFNOR, 2005) et l’Indice Malacologique de qualité des systèmes Lacustres (IMOL) (Mouthon, 1993) ne sont basés que sur un seul groupe de macroinvertébrés (les Oligochètes et les Mollusques respectivement) et ne répondent donc pas aux exigences de la DCE qui préconise l’emploi de l’ensemble des communautés. De plus, des réponses de type pressions/impacts n’ont jamais été mises en évidence à l’échelle nationale. Ces deux indices sont toutefois pris en compte à titre complémentaire pour accroitre le niveau de confiance de l’état évalué des masses d’eau. Le troisième indice français, l’Indice Biotique Lacustre (IBL) (Verneaux, Verneaux, Schmitt, Lovy, & Lambert, 2004), prend en compte la globalité des macroinvertébrés benthiques. Cependant, la méthode d’échantillonnage est très lourde et ne peut donc pas être utilisée à grande échelle. Une simplification (IBL simplifié) de cette méthode, encore en phase de test sur les plans d’eau naturels profonds, a donc été proposé (Mazzella, De Bortoli, & Argillier, 2009b).
Nous présentons ici des travaux réalisés sur (1) la technique d’échantillonnage de la macrofaune benthique en lacs naturels peu profonds, (2) la sélection de métriques utilisant les communautés d’Oligochètes potentiellement utilisables en bioindication dans les retenues nationales. Le choix de ces métriques est basé sur une analyse de type pressions/impacts et (3) la phase exploratoire du développement d’un indice pour les plans d’eau naturels profonds à partir des résultats d’échantillonnages de type IBL et IBL simplifiés.
2. Propositions
de
protocoles
d’échantillonnage
des
communautés
d’invertébrés benthiques en lacs naturels peu profonds
2.1. Contexte et objectif
Pour les lacs naturels, le protocole développé initialement par l’Université de Besançon (Verneaux et al., 2004) puis simplifié (Mazzella, De Bortoli, & Argillier, 2009c), préconise l’échantillonnage de la macrofaune benthique sur deux isobathes, l’un à trois mètres de profondeur, l’autre proche de la zone la plus profonde. Sur les lacs dont la profondeur est voisine ou inférieure à trois mètres, ces deux isobathes ne sont pas identifiables. Sur des plans d’eau dont la profondeur est comprise entre trois et cinq mètres, compte tenu des contraintes opératoires (dérive du bateau en particulier), les différences de profondeurs ne sont sans doute pas très significatives. Une autre méthode de prélèvement doit donc être envisagée d’autant plus que ces milieux ont un fonctionnement particulier dont on peut attendre un impact sur les communautés d’invertébrés benthiques. Tout d’abord, une stratification thermique ne peut se mettre en place dans les lacs peu profonds, même si des stratifications temporaires (à l’échelle de la journée, voire sur quelques jours) peuvent exister (Menetrey-Perrottet, 2009). De plus, la température de l’eau de ces lacs s’ajuste plus rapidement à celle de l’air, ceci à cause de leur surface généralement réduite et de leur faible volume d’eau en comparaison avec les lacs profonds. Ainsi, l’eau des lacs peu profonds peut subir des variations de températures journalières et saisonnières plus importantes que celles des grands lacs. La température de l’eau s’élève également plus rapidement et à une valeur plus élevée (jusque 20°C en été), ce qui favorise la décomposition rapide de la matière organique morte (jusqu’à deux ou trois fois plus rapide qu’au fond des lacs à 4°C) (Menetrey-Perrottet, 2009). Une fai ble profondeur implique aussi la pénétration de la lumière jusqu’au fond, et donc le développement de la végétation sur la plus grande partie de la surface du fond du lac, puisque la photosynthèse y est possible. De plus, les échanges entre l’eau et les sédiments sont plus importants dans les lacs peu profonds (Sondergaard, Jeppesen, & Jensen, 2005). En effet, le fait que ces derniers puissent geler en hiver empêche les apports atmosphériques gazeux qui favorisent l’oxygénation des eaux, ce qui incite la remise en suspension d’une partie du phosphore contenu dans le sédiment.
Ces nombreux éléments mettent en évidence les différences limnologiques importantes entre lacs peu profonds et lacs profonds, d’où la nécessité d’élaborer une méthode de qualification de l’état écologique adaptée à chaque type de système.
Cette première partie de l’étude a pour objectif d’analyser les structures des peuplements benthiques d’un lac naturel peu profond, l’étang des Aulnes (13, France), afin d’établir une stratégie d’échantillonnage adaptée à ce type de milieux .Plus précisément, les communautés ont été décrites à différentes distances à la berge et profondeurs, afin d’analyser leur variabilité « naturelle ».
2.2. Site d’étude et méthode
2.2.1. Le site d’étude
L’étang des Aulnes est situé dans le département des Bouches du Rhône (13, France) sur la plaine de la Crau, à une altitude de 10 m NGF (Figure 1). Cet étang fait parti d’un domaine localisé sur la commune de Saint Martin de Crau ; il est géré par le Conseil Général des Bouches du Rhône depuis 1988 en tant qu’Espace Naturel Sensible.
Figure 1. Localisation géographique de l’étang des Aulnes (site d’étude)
L’étang des Aulnes est d’origine naturelle : il est issu de l’effondrement de la strate de poudingue caractéristique de la plaine de la Crau. La superficie du lac a été estimée à 0,9 km² en 2009 (Agence de l'eau RMC, 2009) et la surface de son bassin versant à 8,6 km². Son périmètre est de 4,5 km et sa profondeur moyenne de 4 m avec un maximum de profondeur mesuré à 5,5 m en 2009 (Tableau 1).
Tableau 1. Principales caractéristiques du site d’étude
Références
Altitude (m NGF)) 10 (Agence de l'eau RMC, 2009)
Périmètre du lac (km) 4,5 (Agence de l'eau RMC, 2009)
Superficie du lac (km²) 0,9 (Agence de l'eau RMC, 2009)
Profondeur moyenne (m) 4 base de données PE Cemagref
Profondeur maximale (m)
5,5
base de données;
(Agence de l'eau RMC, 2009)
Volume (m3) 3340000 base de données PE Cemagref
Superficie bassin versant (km²) 8,6 (Agence de l'eau RMC, 2009)
Temps de séjour (j) 300 base de données PE Cemagref
Marnage (m) 0,6 base de données PE Cemagref
Type lacustre Alcalin
Géologie du Bassin versant Siliceux
Occupation du sol dominante Praire naturelle
Condition de formation du lac Naturelle non glaciaire/Effondrement
Loisirs Pêche depuis la berge
Chasse
Statut de protection
NATURA 2000
Crau (FR9310064)
ZICO
Crau (Pac03)
ZNIEFF
type 1 (n°13-100-150)
2.2.2. Protocoles d’échantillonnage
2.2.2.1. Variabilité du cortège faunistique en fonction de la profondeur
Trois profondeurs ont été échantillonnées : Zmax/2, 0.75 x Zmax (0.75Zm) et 0.9 x Zmax (0.9Zm), Zmax étant la profondeur maximale du lac. Ces profondeurs correspondent aux zones propectées dans deux protocoles actuellement utilisés en France : le protocole normalisé de l’Indice Oligochètes de Bioindication Lacustre (AFNOR, 2005) et le protocole développé par Verneaux et al. (2004) à l’origine de l’Indice Biologique Lacustre.
Ces profondeurs ont été échantillonnées sur cinq transects (Figure 2) de manière à respecter la stratégie d’échantillonnage de l’IBL simplifié (Mazzella et al., 2009c).
Figure 2. Schéma de la stratégie d’échantillonnage pour l’analyse de la variabilité du peuplement
benthique des échantillons en fonction de la profondeur
2.2.2.2. Variabilité du cortège faunistique en fonction de la distance à la berge
Trois distances à la berge ont été testées : 10 (AD10), 20 (AD20) et 40 mètres (AD40) (Figure 3). Comme premier point de prélèvement en Zone littorale, la distance à la berge de 10 mètres a été choisie car c’est celle considérée comme limite de la zone littorale dans le protocole de caractérisation des habitats littoraux (Rowan et al., 2006). Les distances 20 et 40 mètres ont été fixées en mesurant les distances moyennes correspondant aux points de prélèvements les plus près et les plus éloignés de la berge sur l’isobathe -2m en application de l’IBL (Borderelle, 2006; Verneaux et al., 2004).
Figure 3. Schéma de la stratégie d’échantillonnage pour l’analyse de la variabilité du peuplement
benthique des échantillons en fonction de la distance à la berge
Dans la mesure du possible, les points de prélèvements sont répartis le long d’un transect de façon à établir une certaine liaison entre eux comme indiqué sur les figures 2 et 3. Par exemple, pour l’effet distance à la berge, ils sont de préférence positionnés sur des axes de plus faible pente afin d’éviter de fortes variations de profondeur entre les échantillons (Mazzella, De Bortoli, & Argillier, 2009a; Mazzella et al., 2009c).
Chaque transect correspond à un jeu de 3 points regroupant les différentes distances à la berge ou profondeurs étudiées : 7 transects correspondant au test « effet de la distances à la berge » sont obtenus : P231, P233, P234, P235, P236, P237 et P238 (chaque transect regroupe 1 point à 10m + 1 point à 20m + 1 point à 40m) ; 5 transects correspondant au test « effet de la profondeur » sont obtenus : Zm1, Zm2, Zm3, Zm4 et Zm5 (chaque transect regroupe 1 point à 0.5Zmax + 1 point à 0.75Zmax + 1 point à 0.9Zmax).
10M 20M 40M
Ligne à nM distance à la berge 7 Points de prélèvement par ligne traduisant une distance à la berge Isobathe 0.9Zmax, 0.75Zmax et Zmax/2
5 Points de prélèvement par isobathes 0.9Zmax,0.75Zmax et Zmax/2 Zmax/2 0.75Zmax 0.9Zmax
2.2.2.3. Nature du substrat
Pour chaque point de prélèvement, la nature du sustrat (composition et granulométrie) a été décrite en pourcentage de façon grossière par observation suite au fractionnement des particules organiques et minérales dans la colonne de tamis. Les différents paramètres estimés sont :
o % Matière organique fine (<1mm)
o % Matière organique grossière (>1mm)
o % Matière minérale
o % Algues
o % Autres végétaux vivants
2.2.3. Traitement des données
Le niveau de détermination des organismes aurait du être celui recommandé par l’IBL simplifié (Mazzella et al., 2009b). Cependant, celui-ci n’a pas été atteint pour tous les taxons du fait de la complexité dans la détermination (travail de spécialistes). Les Oligochètes et les Chironomidés ont été déterminés au niveau le plus fin possible (Tableau 2)
Tableau 2. Niveau de détermination des macroinvertébrés benthiques
Niveau de détermination atteint dans
cette étude
Nématodes
Embranchement
Oligochètes
Naididae
Tubificinae
Tubificinae avec soies capillaires
Tubificinae sans soie capillaire
Autres
Genre (ou espèce)
Autres
Famille
Diptères
Chironomidae
Chironominae Tribu (+ Chironomus groupe plumosus)
Autres
Sous-famille
Autres
Famille
Autres
Genre (ou espèce)
Une Analyse Factorielle des Correspondances (AFC) (Hirschfeld, 1935) a été réalisée sur le tableau faunistique afin d’observer la structure des peuplements de macroinvertébrés benthiques sur le lac. Les différents points de prélèvements et/ou les taxons sont affichés sur un plan factoriel, sur lequel un regroupement de ces points (distance/profondeur, transect) ou des taxons (Oligochètes, Chironomidae, ETO,…) a été effectué.
Pour identifier les points de prélèvement les plus homogènes d’un point de vue assemblage taxonomique, une classification hiérarchique ascendante utilisant le critère d’agrégation de type Ward et la distance de Bray et Curtis a été appliquée. Des groupes homogènes du point de vue de la composition en macroinvertébrés benthiques ont été identifiés selon le critère de la « Mediane Split Silhouette » (Pollard & van der Laan, 2002); la procédure de permutation multi-réponse (MRPP P<0,001,(Mielke, Berry, & Johnson, 1976) a été utilisée pour tester la significativité de ces groupes.
Une Analyse Canonique des Correspondances (ACC) (Ter Braak, 1986) a enfin été réalisée dans le but de déterminer le rôle des différentes variables environnementales sur la mise en place des communautés de macroinvertébrés benthiques.
2.3. Résultats
La liste faunistique est dominée par les oligochètes (55 % de l’abondance totale), les nématodes (34%) et les chironomes (8%). Les oligochètes sont les plus diversifiés avec 16 taxons dont 14 appartiennent à la famille des Naididae.
Des différences importantes sont observées en ce qui concerne la densité totale, la richesse taxonomique et la densité en Oligochètes sensibles entre les différentes zones prospectées (AD10,
AD20, AD40, 0.75Zm et 0.9Zm).
En effet, la densité en macroinvertébrés augmente avec la distance à la berge et la profondeur. Elle est six fois plus importante à la profondeur maximale étudiée (0.9Zm) qu’à 10 mètres de la berge (AD10).
La richesse taxonomique est maximale à 20 mètres de la berge (48 taxons en AD20) et décroît au fur et à mesure que la profondeur augmente ; à 0,9Zm, nous n’observons plus que seize taxons.
Enfin, les quatre taxons de la famille des Naididae sensibles à la pollution dans les sédiments lacustres disparaissent à partir d’une profondeur égale à 75 % de la profondeur maximale. La forte diminution de cette densité en fonction de la distance à la berge (entre 10 et 40 mètres) est essentiellement due à la variation d’abondance de Stylaria lacustris (367 individus/m² à 10 mètres de la berge, 82 à 20 mètres et 7 à 40 mètres).
La figure suivante (Figure 4)présente les résultats de l’AFC réalisée sur la liste faunistique de chaque point de prélèvement. Le diagramme des valeurs propres montre que les axes F1 et F2 expliquent respectivement 27,4 % et 14,7 % de l’inertie totale du tableau. Les deux premiers axes factoriels sont donc conservés et expliquent près de 42 % de la variance observée.
Les différents groupes constitués sur la base de la distance à la berge et de la profondeur peuvent être distingués du point de vue des assemblages d’invertébrés benthiques. En effet, une forte séparation existe entre les points de prélèvements situés à 10 et 20 mètres de la berge et ceux à 40 mètres. De plus, ces derniers se rapprochent des points de profondeurs plus élevées.
Enfin, le groupe qui semble le plus homogène est celui constitué des points échantillonnés à une profondeur égale à 90% de la profondeur maximale.
d = 0.5 0.75Zm 0.9Zm AD10 AD20 AD40 27.44 % 14.77 % F2 F1 d = 0.5 0.75Zm 0.9Zm AD10 AD20 AD40 27.44 % 14.77 % d = 0.5 d = 0.5 0.75Zm 0.9Zm AD10 AD20 AD40 27.44 % 14.77 % F2 F1 F2 F1
Figure 4. Plan F1 et F2 de l’analyse factorielle des correspondances (AFC) réalisée sur le tableau
faunistique des macroinvertébrés de l’étang des Aulnes. Les points de prélèvements sont regroupés en fonction de la distance à la berge et de la profondeur (AD10, AD20 et AD40 : 10, 20 et 40 mètres respectivement de la berge ; 0.75Zm et 0.9Zm : 75 et 90% respectivement de la profondeur maximale). L’histogramme en bas à droite est celui des valeurs propres de l’AFC.
Les groupes de points de prélèvements se distinguent également fortement en fonction de leur appartenance à un transect (Figure 5). En effet, les différents transects peuvent être séparés en trois groupes :
- les transects P233, P234 et P235 ;
- les transect P231 et P236 ;
- et le transect P238.
Le transect P237 se trouve entre les deux premiers groupes cités.
Les points prélevés à 0.75 et 0.9 de la profondeur maximale semblent homogènes entre les différents transects.
d = 0.5 P231 P233 P234 P235 P236 P237 P238 Zm1 Zm3 Zm4 Zm5 27.44 % 14.77 % Eigenvalues F2 F1 d = 0.5 P231 P233 P234 P235 P236 P237 P238 Zm1 Zm3 Zm4 Zm5 27.44 % 14.77 % Eigenvalues F2 F1 F2 F1
Figure 5. Plans F1 et F2 de l’analyse factorielle des correspondances (AFC) réalisée sur le tableau faunistique des macroinvertébrés de l’étang des Aulnes. Les points de prélèvements sont regroupés en fonction des différents transects. L’histogramme en bas à droite est celui des valeurs propres de l’AFC.
Les résultats de la classification hiérarchique de tous les points de prélèvements sont présentés en Figure 6. Trois groupes homogènes et significativement différents dans leur composition en macroinvertébrés benthiques ont été retenus.
Le groupe 1 est constitué de tous les points de prélèvement situés à 10 et 20 mètres de la berge. Les points localisés à 90% de la profondeur maximale constituent le groupe 3 avec AD20P235, AD40P235, AD40P234, ainsi que AP0.75 Zm1, Zm4 et Zm5. Enfin, le groupe 2 comprend les autres points de prélèvements. Il est à noter que le groupe le plus homogène est le groupe 3 tandis que le groupe 1 est le plus hétérogène.
Figure 6. Dendrogramme issu de la classification ascendante hiérarchique (distance de Bray et
Curtis, méthode d’agrégation de type « ward »)
La profondeur ainsi que la distance à la berge vont en augmentant du groupe 1 au groupe 3 (Figure 7). Le groupe 1 est caractérisé par des profondeurs et des distances à la berge faibles et une part importante du substrat constitué de matière organique grossière. Au contraire, le groupe 3 est défini par des points de prélèvements situés à des profondeurs plus grandes et dont la part de sédiments fins est plus importante que dans les autres groupes.
1 2 3 2 3 4 5 groupes p ro fo n d e u r (m ) 1 2 3 0 1 0 0 2 0 0 3 0 0 groupes d is ta n c e à l a b e rg e ( m ) 1 2 3 0 2 0 4 0 6 0 8 0 1 0 0 groupes m a ti è re o rg a n iq u e f in e ( 0 ,2 5 -1 m m ) (% ) 1 2 3 0 2 0 4 0 6 0 8 0 groupes m a ti è re o rg a n iq u e g ro s s iè re ( > 1 m m ) (% )
Figure 7. Boîtes à moustache représentant la variation de quatre conditions environnementales
(profondeur, distance à la berge, pourcentage de substrat fin et de substrat grossier) entre les trois groupes issus de la classification hiérarchique ascendante
Les axes F1 et F2 du premier plan factoriel de l’ACC (Figure 8) expliquent respectivement environ 46 % et 22 % de la variance observée. L’axe F1 est très corrélé positivement avec la profondeur et la distance à la berge alors que l’axe F2 expliquerait plutôt la nature du substrat. De plus, d’après les poids des différentes variables environnementales, la taille de la matière organique semble être prépondérante dans la mise en place des communautés de macroinvertébrés, ainsi que la profondeur. Cette dernière jouerait un rôle beaucoup plus important que la distance à la berge.
Les points de prélèvements appartenant au groupe 3 créé précédemment par une classification hiérarchique ascendante (Figure 6) se retrouvent corrélés avec l’axe F1; les communautés semblent donc être sous l’influence de la profondeur (et accessoirement de la distance à la berge). En revanche, tous les autres points échantillonnés se répartissent dans le plan factoriel plutôt en fonction du second axe ; la nature du substrat serait donc un facteur important dans la mise en place des communautés de macroinvertébrés de ces points.
-3
-2
-1
0
1
2
3
-3
-2
-1
0
1
2
F
1F
2 AD10P231 AD20P231 AD40P231 AD10P233 AD20P233 AD40P233 AD10P234 AD20P234 AD40P234 AD10P235 AD20P235 AD40P235 AD10P236 AD20P236 AD40P236 AD10P237 AD20P237 AD40P237 AD10P238 AD20P238 AD40P238 AP0.75Zm1 AP0.9Zm1 AP0.75Zm2 AP0.9ZM2 AP0.75Zm3 AP0.9Zm3 AP0.75Zm4 AP0.9Zm4 AP0.75Zm5 AP0.9Zm5Profondeur
Distance
MO<1mm
MO>1mm
algue
Végétaux vivants
0
1
48.44 %
21.98 %
-1
Eigenvalues d = 0.1 Loadings d = 0.1 Profondeur Distance MO<1mm MO>1mm Algues Végétaux vivants Poids-3
-2
-1
0
1
2
3
-3
-2
-1
0
1
2
F
1F
2 AD10P231 AD20P231 AD40P231 AD10P233 AD20P233 AD40P233 AD10P234 AD20P234 AD40P234 AD10P235 AD20P235 AD40P235 AD10P236 AD20P236 AD40P236 AD10P237 AD20P237 AD40P237 AD10P238 AD20P238 AD40P238 AP0.75Zm1 AP0.9Zm1 AP0.75Zm2 AP0.9ZM2 AP0.75Zm3 AP0.9Zm3 AP0.75Zm4 AP0.9Zm4 AP0.75Zm5 AP0.9Zm5Profondeur
Distance
MO<1mm
MO>1mm
algue
Végétaux vivants
0
1
48.44 %
21.98 %
-1
Eigenvalues d = 0.1 Loadings d = 0.1 Profondeur Distance MO<1mm MO>1mm Algues Végétaux vivants Poids-3
-2
-1
0
1
2
3
-3
-2
-1
0
1
2
F
1F
2 AD10P231 AD20P231 AD40P231 AD10P233 AD20P233 AD40P233 AD10P234 AD20P234 AD40P234 AD10P235 AD20P235 AD40P235 AD10P236 AD20P236 AD40P236 AD10P237 AD20P237 AD40P237 AD10P238 AD20P238 AD40P238 AP0.75Zm1 AP0.9Zm1 AP0.75Zm2 AP0.9ZM2 AP0.75Zm3 AP0.9Zm3 AP0.75Zm4 AP0.9Zm4 AP0.75Zm5 AP0.9Zm5Profondeur
Distance
MO<1mm
MO>1mm
algue
Végétaux vivants
0
1
48.44 %
21.98 %
-1
Eigenvalues-3
-2
-1
0
1
2
3
-3
-2
-1
0
1
2
F
1F
2 AD10P231 AD20P231 AD40P231 AD10P233 AD20P233 AD40P233 AD10P234 AD20P234 AD40P234 AD10P235 AD20P235 AD40P235 AD10P236 AD20P236 AD40P236 AD10P237 AD20P237 AD40P237 AD10P238 AD20P238 AD40P238 AP0.75Zm1 AP0.9Zm1 AP0.75Zm2 AP0.9ZM2 AP0.75Zm3 AP0.9Zm3 AP0.75Zm4 AP0.9Zm4 AP0.75Zm5 AP0.9Zm5Profondeur
Distance
MO<1mm
MO>1mm
algue
Végétaux vivants
0
1
48.44 %
21.98 %
-3
-2
-1
0
1
2
3
-3
-2
-1
0
1
2
F
1F
2 AD10P231 AD20P231 AD40P231 AD10P233 AD20P233 AD40P233 AD10P234 AD20P234 AD40P234 AD10P235 AD20P235 AD40P235 AD10P236 AD20P236 AD40P236 AD10P237 AD20P237 AD40P237 AD10P238 AD20P238 AD40P238 AP0.75Zm1 AP0.9Zm1 AP0.75Zm2 AP0.9ZM2 AP0.75Zm3 AP0.9Zm3 AP0.75Zm4 AP0.9Zm4 AP0.75Zm5 AP0.9Zm5Profondeur
Distance
MO<1mm
MO>1mm
algue
Végétaux vivants
0
1
48.44 %
21.98 %
-3
-2
-1
0
1
2
3
-3
-2
-1
0
1
2
F
1F
2-3
-2
-1
0
1
2
3
-3
-2
-1
0
1
2
F
1F
2 AD10P231 AD20P231 AD40P231 AD10P233 AD20P233 AD40P233 AD10P234 AD20P234 AD40P234 AD10P235 AD20P235 AD40P235 AD10P236 AD20P236 AD40P236 AD10P237 AD20P237 AD40P237 AD10P238 AD20P238 AD40P238 AP0.75Zm1 AP0.9Zm1 AP0.75Zm2 AP0.9ZM2 AP0.75Zm3 AP0.9Zm3 AP0.75Zm4 AP0.9Zm4 AP0.75Zm5 AP0.9Zm5Profondeur
Distance
MO<1mm
MO>1mm
algue
AD10P231 AD20P231 AD40P231 AD10P233 AD20P233 AD40P233 AD10P234 AD20P234 AD40P234 AD10P235 AD20P235 AD40P235 AD10P236 AD20P236 AD40P236 AD10P237 AD20P237 AD40P237 AD10P238 AD20P238 AD40P238 AP0.75Zm1 AP0.9Zm1 AP0.75Zm2 AP0.9ZM2 AP0.75Zm3 AP0.9Zm3 AP0.75Zm4 AP0.9Zm4 AP0.75Zm5 AP0.9Zm5Profondeur
Distance
MO<1mm
MO>1mm
algue
Végétaux vivants
0
1
48.44 %
21.98 %
-1
Eigenvalues-1
Eigenvalues d = 0.1 Loadings d = 0.1 Profondeur Distance MO<1mm MO>1mm Algues Végétaux vivants Poids d = 0.1 Loadings d = 0.1 Profondeur Distance MO<1mm MO>1mm Algues Végétaux vivants Poids d = 0.1 Loadings d = 0.1 Profondeur Distance MO<1mm MO>1mm Algues Végétaux vivants PoidsFigure 8. Plans factoriel F1et F2 de l’Analyse canonique des correspondances (ACC) réalisée sur
l’ensemble du jeu de données, abondances des taxons et variables environnementales. En bas à gauche : poids des différentes variables environnementales ; MO = Matière organique ; en bas à droite : histogramme des valeurs propres de l’ACC.
Au vu de ces résultats, le groupe 3 paraît le groupe le plus homogène ainsi que le groupe le moins influencé par la nature du substrat. De plus, tous les points de prélèvements situés à une profondeur égale à 90% de la profondeur maximale se retrouvent dans ce groupe.
2.4. Conclusions
Les résultats obtenus sur l’Etang des Aulnes peuvent donner quelques indications quant à la diversité du peuplement benthique à différentes profondeurs et distances à la berge et donc les orientations à prendre dans la mise en place d’un protocole d’échantillonnage. Cependant, il serait nécessaire au préalable d’appliquer ce même exercice à un plus grand nombre de lacs peu profonds pour pouvoir confirmer ces résultats, comme par exemple sur le deuxième lac méditerranéen situé dans les Bouches du Rhône (Entressen), ainsi que sur d’autres qui se trouvent majoritairement en région Aquitaine.
Les résultats de l’échantillonnage seront d’autant plus stables que la variabilité liée à l’environnement sera faible. Cependant, il est aussi nécessaire de conserver un maximum d’information pour caractériser la qualité du milieu.
Au vue de l’ensemble des résultats, l’échantillonnage de l’isobathe 0.9Zm de la zone profonde semble être le plus approprié pour éviter toutes variations naturelles dans la composition de la macrofaune benthique. Néanmoins, la perte d’information due à un échantillonnage uniquement en zone profonde par rapport à un échantillonnage plus représentatif de l’intégralité du lac est élevée. En effet, même si la densité en macroinvertébrés benthiques est plus importante dans les points de prélèvements de l’isobathe 0.9Zm, les deux tiers des taxons retrouvés dans l’étang des Aulnes sont absents de cette zone. Les taxons des groupes d’éphéméroptères, de trichoptères, d’odonates et de mollusques observés en zone littorale disparaissent complètement en zone profonde, ainsi que la majorité des genres d’oligochètes. De plus, aucune espèce de Naididae sensible à la pollution dans les sédiments lacustres cités dans l’IOBL n’est retrouvée à la plus grande profondeur, alors que leur densité est importante à 10 mètres de la berge. Ces différentes observations vont donc plutôt en faveur d’un échantillonnage de la zone littorale, comme cela est déjà le cas dans l’Indice Biologique Lacustre (IBL) (Verneaux et al., 2004) ainsi que dans l’Indice Malacologique de qualité des systèmes Lacustres (IMOL) (Mouthon, 1993). Cependant, le substrat dans cette zone est très hétérogène et les communautés de macroinvertébrés subissent des variations importantes. Il pourrait donc être nécessaire de décrire la nature du substrat afin de pouvoir comparer différents lacs peu profonds entre eux. C’est aussi pour pallier à cette forte diversité en zone littorale qu’un même substrat (e.g. sédiments fins) et qu’un nombre important d’échantillons sont préconisés par le protocole d’échantillonnage de l’IBL ou dans sa version simplifié (Mazzella et al., 2009b).
A ce niveau de l’étude, l’ensemble des résultats montre qu’un échantillonnage en zone littorale et profonde « serait nécessaire » pour une bonne représentation qualitative et quantitative de la communauté macrobenthique indicatrice de l’état du milieu.
Néanmoins, ce travail ne constitue qu’une première étape de la démarche. L’identification de l’isobathe en zone littorale par rapport à une distance à la berge n’a pas été définie précisément. De plus, le rôle de la distance à la berge n’a pas pu être dissocié de celui de la profondeur (dont le poids semble plus important sur la mise en place des communautés de macroinvertébrés benthiques), ces deux variables environnementales étant très corrélées entre elles. A notre connaissance, aucune étude n’a cependant été réalisée pour dissocier ces effets, certainement à cause de la difficulté à faire varier la distance à la berge sans que la profondeur ne soit affectée.
Cette étape franchie, l’effort d’échantillonnage à appliquer sur chaque strate restera à définir, en particulier sur la zone littorale où il influencera fortement la diversité spécifique. Cette définition passe nécessairement par l’analyse de l’effet de l’effort d’échantillonnage sur la stabilité des métriques dans différentes conditions environnementales, ce qui représente un travail considérable en terme de prélèvements et de déterminations taxinomiques.
Il est donc proposé à ce stade d’analyser les données collectées en application du protocole IOBL. Ces données ont été collectées a priori en zone profonde sur lesquelles l’homogénéité des échantillons est démontrée ici. En fonction des résultats et si elles sont suffisamment nombreuses, des explorations complémentaires seront envisagées afin de préciser la méthode à mettre en œuvre pour compléter cet échantillonnage par des prélèvements en zone littorale.
3. Développement d’outils de bioindication basés sur les
communautés d’invertébrés benthiques lacustres
3.1.
Métriques adaptées aux plans d’eau artificiels
3.1.1. Les données disponibles
3.1.1.1. Description des plans d’eau
Les données concernent principalement des retenues mais des plans d’eau naturels qui ont subit le même protocole de prélèvement de macroinvertébrés sont également considérés.
Des données sont disponibles pour plus de 188 plans d’eau avec une certaine hétérogénéité dans la manière dont les variables sont renseignées pour tous ces plans d’eau. Après sélection des plans d’eau pour lesquels un maximum de variables est disponible, un jeu de 115 plans d’eau a été constitué dont 78 Retenues et 37 lacs naturels.
3.1.1.2. Données faunistiques
Le protocole IOBL
Le protocole utilisé pour recueillir les données faunistiques sur les retenues est celui de la norme (AFNOR, 2005) relative à l’IOBL (Indice Oligochètes de Bioindication lacustre). Les sédiments sont prélevés à l’aide d’une benne de prélèvement (e.g. Ekman ou Friedinger). Un échantillon est constitué d’au moins 3 prélèvements soit environ 0.1m² de surface de sédiments. En général, un seul échantillon est réalisé à la plus grande profondeur pour les plans d’eau inférieurs à 5 hectares. Pour ceux dont la superficie est égale ou supérieure à 5 hectares, le nombre d’échantillons est variable. Des échantillons, souvent deux, réalisés à Zmax/2 peuvent compléter le jeu de données pour l’ensemble du système lacustre. Pour un échantillon, les prélèvements sont effectués au hasard à la profondeur considérée, séparés d’une dizaine de mètres respectivement. Le contenu des bennes est ensuite tamisé à 0,315mm de vide de maille. La période d’échantillonnage se fait généralement en été jusqu’au début de l’automne.
Concernant la détermination au laboratoire, le niveau d’identification requis est l’espèce pour la majorité des taxons ou un ensemble taxonomique plus général (genre, famille, sous-famille).
Les données faunistiques
Les données faunistiques recueillies proviennent de la base plans d’eau (Pôle ONEMA-CEMAGREF), elles sont issues du protocole IOBL réalisé sur l’ensemble du territoire français dans le cadre des suivis réseau (e.g. Référence, RCS, RCO).
La norme Afnor NF T90-391 ayant été éditée en 2005, les jeux de données utilisés ne contiennent que les plans d’eau dans lesquels l’IOBL a été effectué à partir de cette date afin de retirer le biais de protocole non standardisé (e.g. nb échantillons, surface de prélèvement et niveau de détermination). Dans le but d’obtenir des listes faunistiques les plus comparables possibles, seules les données correspondant aux oligochètes de la zone centrale ont été prises en compte. En effet, sur quelques plans d’eau, des zones dites intermédiaires ont également fait l’objet de prélèvements mais ce groupe de plans d’eau a été jugé encore trop peu important pour le développement de l’indice.
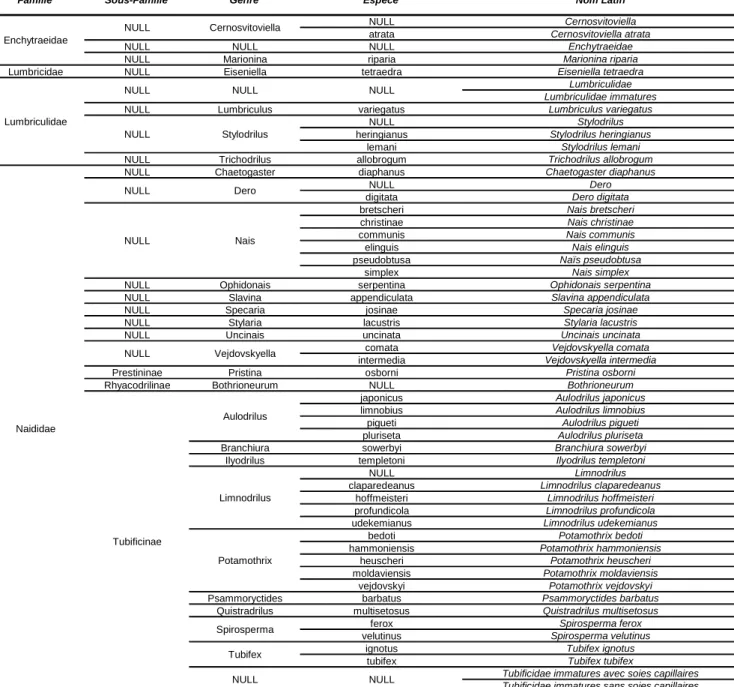
D’autre part, seuls les plans d’eau ayant une richesse spécifique supérieure à 1 ont été conservés. Le détail de la liste faunistique se trouve dans le Tableau 3.
Tableau 3. Détails de la liste faunistique
Famille Sous-Famille Genre Espece Nom Latin
NULL Cernosvitoviella
atrata Cernosvitoviella atrata
NULL NULL NULL Enchytraeidae
NULL Marionina riparia Marionina riparia
Lumbricidae NULL Eiseniella tetraedra Eiseniella tetraedra Lumbriculidae Lumbriculidae immatures
NULL Lumbriculus variegatus Lumbriculus variegatus
NULL Stylodrilus
heringianus Stylodrilus heringianus
lemani Stylodrilus lemani
NULL Trichodrilus allobrogum Trichodrilus allobrogum
NULL Chaetogaster diaphanus Chaetogaster diaphanus
NULL Dero
digitata Dero digitata
bretscheri Nais bretscheri
christinae Nais christinae
communis Nais communis
elinguis Nais elinguis
pseudobtusa Naïs pseudobtusa
simplex Nais simplex
NULL Ophidonais serpentina Ophidonais serpentina
NULL Slavina appendiculata Slavina appendiculata
NULL Specaria josinae Specaria josinae
NULL Stylaria lacustris Stylaria lacustris
NULL Uncinais uncinata Uncinais uncinata
comata Vejdovskyella comata
intermedia Vejdovskyella intermedia
Prestininae Pristina osborni Pristina osborni
Rhyacodrilinae Bothrioneurum NULL Bothrioneurum
japonicus Aulodrilus japonicus
limnobius Aulodrilus limnobius
pigueti Aulodrilus pigueti
pluriseta Aulodrilus pluriseta
Branchiura sowerbyi Branchiura sowerbyi
Ilyodrilus templetoni Ilyodrilus templetoni
NULL Limnodrilus
claparedeanus Limnodrilus claparedeanus
hoffmeisteri Limnodrilus hoffmeisteri
profundicola Limnodrilus profundicola
udekemianus Limnodrilus udekemianus
bedoti Potamothrix bedoti
hammoniensis Potamothrix hammoniensis
heuscheri Potamothrix heuscheri
moldaviensis Potamothrix moldaviensis
vejdovskyi Potamothrix vejdovskyi
Psammoryctides barbatus Psammoryctides barbatus
Quistradrilus multisetosus Quistradrilus multisetosus
ferox Spirosperma ferox
velutinus Spirosperma velutinus
ignotus Tubifex ignotus
tubifex Tubifex tubifex
Tubificidae immatures avec soies capillaires Tubificidae immatures sans soies capillaires
NULL Tubifex Spirosperma NULL Aulodrilus Tubificinae Limnodrilus Potamothrix NULL NULL Stylodrilus NULL Dero NULL Enchytraeidae Lumbriculidae Naididae Cernosvitoviella NULL NULL NULL Nais NULL Vejdovskyella
3.1.1.3. Paramètres environnementaux naturels
Vingt cinq paramètres environnementaux de différentes origines (mésologique, climatique…) sont potentiellement disponibles pour les plans d’eau ayant fait l’objet d’un échantillonnage de type IOBL.
Une première sélection de paramètres a été réalisée sur la base d’une analyse en composantes principales (ACP) croisée aux valeurs de corrélations de Spearman entre ces paramètres afin d’éviter d’utiliser dans les modèles des paramètres fortement corrélés.
De ces deux résultats, les paramètres retenus sont :
- la superficie du plan d’eau en km² (SuperfPla),
- la profondeur maximale du plan d’eau en m (ProfMaxPla),
- la surface du bassin versant en km² (Surf_BV),
- le temps de séjour en années (TpsSim),
- L’écart entre la température moyenne de juillet et celle de janvier en °C (Jui_moins_Jan)
- Les précipitations moyennes annuelles en mm (PrecipMoyAnnuelle)
3.1.1.4. Pressions anthropiques
Trois types de pressions anthropiques sont associés à ces données : pressions d’occupation du sol, de densité de population et de chimie.
Les cinq variables d’occupation du sol sont exprimées en pourcentage surfacique (urbanisme, agriculture, agriculture intensive, 1-occupation du sol naturelle et semi-naturelle, 1-occupation du sol naturelle et semi-naturelle, agriculture extensive inclue) et les deux variables de densité de populations sur le bassin versant sont la moyenne et l’écart-type en nombre d’habitants au km². Des pressions chimiques mesurées dans les sédiments en milligrammes par kilogramme de matière sèche (azote kjeldahl, phosphore et carbone) sont également disponibles. Ces variables chimiques seront dans ce travail intermédiaire considérées comme des pressions même si elles ne sont pas totalement d’origine anthropique et que la suite de notre travail devra prendre en compte cet élément.
3.1.1.5. Métriques
Les métriques ont été classifiées selon (Hering, Feld, Moog, & Ofenbock, 2006) en différents types :
1. les métriques d’abondance et de composition 2. les métriques de richesse et de diversité
3. les métriques employant les traits fonctionnels des taxons
4. les métriques employant le degré de sensibilité/tolérance des taxons aux pollutions.
Le nombre de métriques qu’il est possible de calculer et de tester dans notre cas est limité par le jeu de données qui concerne uniquement les oligochètes.
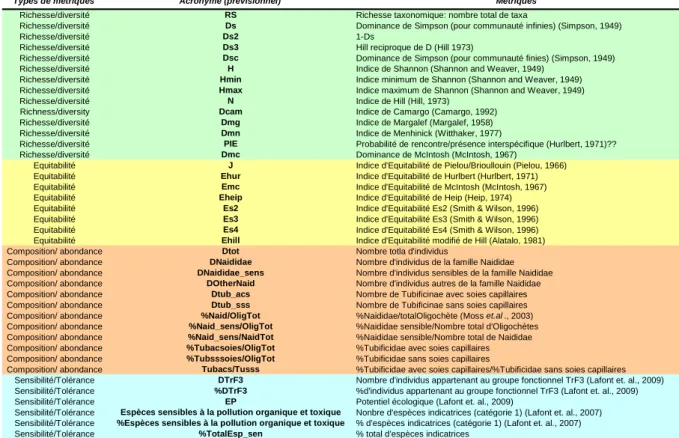
Le Tableau 4 rassemble l’ensemble des métriques calculées avant les phases de sélection. Ces métriques sont soit des indices classiques de diversité et équitabilité, soit des métriques de composition ou d’abondance de taxons oligochètes, soit des métriques représentant la sensibilité ou tolérance d’espèces oligochètes aux pollutions selon (Lafont et al., 2007)
Tableau 4. Description de l’ensemble des métriques calculées avant sélection
Types de métriques Acronyme (previsionnel) Métriques
Richesse/diversité RS Richesse taxonomique: nombre total de taxa
Richesse/diversité Ds Dominance de Simpson (pour communauté infinies) (Simpson, 1949)
Richesse/diversité Ds2 1-Ds
Richesse/diversité Ds3 Hill reciproque de D (Hill 1973)
Richesse/diversité Dsc Dominance de Simpson (pour communauté finies) (Simpson, 1949)
Richesse/diversité H Indice de Shannon (Shannon and Weaver, 1949)
Richesse/diversité Hmin Indice minimum de Shannon (Shannon and Weaver, 1949)
Richesse/diversité Hmax Indice maximum de Shannon (Shannon and Weaver, 1949)
Richesse/diversité N Indice de Hill (Hill, 1973)
Richness/diversity Dcam Indice de Camargo (Camargo, 1992)
Richesse/diversité Dmg Indice de Margalef (Margalef, 1958)
Richesse/diversité Dmn Indice de Menhinick (Witthaker, 1977)
Richesse/diversité PIE Probabilité de rencontre/présence interspécifique (Hurlbert, 1971)??
Richesse/diversité Dmc Dominance de McIntosh (McIntosh, 1967)
Equitabilité J Indice d'Equitabilité de Pielou/Brioullouin (Pielou, 1966)
Equitabilité Ehur Indice d'Equitabilité de Hurlbert (Hurlbert, 1971)
Equitabilité Emc Indice d'Equitabilité de McIntosh (McIntosh, 1967)
Equitabilité Eheip Indice d'Equitabilité de Heip (Heip, 1974)
Equitabilité Es2 Indice d'Equitabilité Es2 (Smith & Wilson, 1996)
Equitabilité Es3 Indice d'Equitabilité Es3 (Smith & Wilson, 1996)
Equitabilité Es4 Indice d'Equitabilité Es4 (Smith & Wilson, 1996)
Equitabilité Ehill Indice d'Equitabilité modifié de Hill (Alatalo, 1981)
Composition/ abondance Dtot Nombre totla d'individus
Composition/ abondance DNaididae Nombre d'individus de la famille Naididae
Composition/ abondance DNaididae_sens Nombre d'individus sensibles de la famille Naididae
Composition/ abondance DOtherNaid Nombre d'individus autres de la famille Naididae
Composition/ abondance Dtub_acs Nombre de Tubificinae avec soies capillaires
Composition/ abondance Dtub_sss Nombre de Tubificinae sans soies capillaires
Composition/ abondance %Naid/OligTot %Naididae/totalOligochète (Moss et.al ., 2003)
Composition/ abondance %Naid_sens/OligTot %Naididae sensible/Nombre total d'Oligochètes
Composition/ abondance %Naid_sens/NaidTot %Naididae sensible/Nombre total de Naididae
Composition/ abondance %Tubacsoies/OligTot %Tubificidae avec soies capillaires
Composition/ abondance %Tubsssoies/OligTot %Tubificidae sans soies capillaires
Composition/ abondance Tubacs/Tusss %Tubificidae avec soies capillaires/%Tubificidae sans soies capillaires
Sensibilité/Tolérance DTrF3 Nombre d'individus appartenant au groupe fonctionnel TrF3 (Lafont et. al., 2009)
Sensibilité/Tolérance %DTrF3 %d'individus appartenant au groupe fonctionnel TrF3 (Lafont et. al., 2009)
Sensibilité/Tolérance EP Potentiel écologique (Lafont et. al., 2009)
Sensibilité/Tolérance Espèces sensibles à la pollution organique et toxique Nonbre d'espèces indicatrices (catégorie 1) (Lafont et. al., 2007)
Sensibilité/Tolérance %Espèces sensibles à la pollution organique et toxique % d'espèces indicatrices (catégorie 1) (Lafont et. al., 2007)
Sensibilité/Tolérance %TotalEsp_sen % total d'espèces indicatrices
3.1.2. L’approche méthodologique
L’objectif de cette étude est de définir quelles peuvent être les métriques pouvant être utilisées comme outils de bioindication pour définir l’état écologique des plans d’eau artificiels, métriques qui seraient sensibles aux perturbations anthropiques et qui répondraient donc le mieux aux pressions exercées sur les plans d’eau.
Dans les approches les plus classiques, afin d’étudier les réponses de métriques aux pressions anthropiques, la part due à l’environnement est dans un premier temps éliminée en utilisant un jeu de données composé uniquement de sites de références et comme métriques les résidus d’un modèle composé uniquement de paramètres environnementaux (Pont et al., 2006). Ces résidus sont ensuite normalisés, transformés en « EQR » (Ecological Quality ratio) et confrontés à des paramètres de pressions.
Le nombre de plans d’eau de référence dans notre jeu de données est non seulement assez faible (35 plans d’eau), mais aussi peu fiable car la notion de référence au niveau des retenues n’a pas encore été validée par les experts. Nous avons donc préféré une approche de type « hindcasting » à l’approche classique (basée sur des plans d’eau de référence). Cette approche est justement adaptée lorsque le nombre de site de référence ne permet pas l’approche classique (Baker et al., 2005).
L’hindcasting, inspiré par (Baker et al., 2005; Kilgour & Barton, 1999; Kilgour & Stanfield, 2006) et également utilisé dans le rapport de développement de bioindicateurs basés sur l’ichtyofaune (Launois, Veslot, Irz, & Argillier, Under review; Pédron, De Bortoli, & Argillier, 2010), consiste à développer dans un premier temps un modèle de prédiction des métriques par les paramètres environnementaux et les paramètres de pression puis d’utiliser dans un second temps ce même modèle en injectant un nouveau jeu de données dans lequel les valeurs correspondants aux
paramètres de pression sont toutes fixées à 0, afin d’obtenir des valeurs de la métrique en absence de pression.
L’écart entre la valeur observée et prédite de la métrique peut ensuite être calculé pour obtenir des résidus. Ces résidus sont normalisés sous forme d’EQR (Ecological Quality Ratio) et alors mis en relation avec les pressions retenues dans les modèles.
Les corrélations entre métriques sont aussi calculées afin de procéder à une sélection définitive de métriques, c'est-à-dire celles qui répondent le plus à la pression sans être redondantes.
L’ensemble des paramètres est au préalable transformé afin de répondre aux exigences des modèles linéaires utilisés. Une transformation en log est appliquée aux paramètres environnementaux, aux pressions sauf celles d’occupation du sol et aux métriques qui ne sont pas en pourcentage. Une transformation en arcsinus de la racine-carré de la variable est appliquée aux pourcentages d’occupation du sol et aux métriques en pourcentage.
Les métriques subissent une première phase de sélection en fonction de leur corrélations (>0.2 et significatives) avec les facteurs de pression.
Les modèles utilisés sont des régressions linéaires multiples. L’ensemble des paramètres environnementaux est proposé comme variables explicatives du modèle ainsi que le carré de ses variables et tous les paramètres de pressions. Une procédure « pas à pas » mixte (stepwise) est ensuite appliquée permettant de ne conserver que les variables pertinentes à la construction du modèle avec l’aide du critère de l’information d’Aikaike (AIC) (Akaike, 1974). Seuls les modèles présentant un R² ajusté supérieur à 0.3 sont retenus pour l’hindcasting, ayant pour conséquence encore une fois une nouvelle sélection de métriques.
3.1.3. Résultats quant à la proposition de métriques candidates pour les plans
d’eau artificiels
3.1.3.1. Prétraitement des données
Dans notre jeu de données, nous disposons de données provenant des retenues et de lacs naturels. Une première étape d’exploration des données a été nécessaire pour définir si les retenues et les lacs naturels devaient être pris en compte indépendamment ou non. Une classification hiérarchique (en Figure 9a) sur les données faunistiques et une analyse en composante principale sur les paramètres environnementaux (en Figure 9b) ont été menées. Nous pouvons voir des différences au niveau de la composition faunistique puisque un groupe ne comprenant pratiquement que des retenues se détache des deux autres. Cependant les deux autres groupes sont composés aussi bien de retenues que de lacs naturels. Les communautés d’invertébrés benthiques ne sont donc pas si clairement différenciées entre les deux types de plans d’eau. Les paramètres environnementaux séparent également les lacs naturels des retenues tout en conservant des chevauchements entre les deux types de milieux.
Par ailleurs, l’étude séparée des retenues n’a pas donné de résultats différents, si ce n’est de moins bons résultats du fait du nombre plus faible d’échantillons ce qui nous a poussé à conserver notre jeu complet contenant aussi bien les retenues que les plans d’eau naturels, considérant que le faible nombre de lacs naturels n’influencerait pas les résultats observables sur les retenues.
0 2 4 6 8 hclust T U I4 5 _ 2 0 0 6 L A V 2 3 _ 2 0 0 7 R L B 1 1 _ 2 0 0 9 S C R 0 4 _ 2 0 0 7 R S V 3 4 _ 2 0 0 7 E S T 1 1 _ 2 0 0 7 T Y X 6 3 _ 2 0 0 6 T R E 3 5 _ 2 0 0 6 R IN 5 3 _ 2 0 0 6 S F P 5 3 _ 2 0 0 6 R G L 6 9 _ 2 0 0 9 A P R 8 5 _ 2 0 0 7 V IO 4 4 _ 2 0 0 6 A R Z 3 5 _ 2 0 0 7 P R O 4 4 _ 2 0 0 6 C A E 3 5 _ 2 0 0 6 N O Y 5 6 _ 2 0 0 7 E P T 3 5 _ 2 0 0 6 C A Z 4 0 _ 2 0 0 5 B L I3 6 _ 2 0 0 6 P D F 4 4 _ 2 0 0 6 C A R 3 5 _ 2 0 0 6 O U E 3 5 _ 2 0 0 6 B A Y 5 8 _ 2 0 0 6 B O U 3 5 _ 2 0 0 6 J O U 1 1 _ 2 0 0 8 R E A 1 3 _ 2 0 0 9 B O S 2 2 _ 2 0 0 7 R O P 2 2 _ 2 0 0 7 B O R 3 3 _ 2 0 0 7 C A Z 4 0 _ 2 0 0 7 C O D 2 B _ 2 0 0 9 E T I3 9 _ 2 0 0 7 M O N 7 4 _ 2 0 0 5 B U L 8 5 _ 2 0 0 7 E C H 3 3 _ 2 0 0 7 B O U 6 6 _ 2 0 0 6 B O U 6 6 _ 2 0 0 7 R M N 2 9 _ 2 0 0 8 N D C 3 8 _ 2 0 0 9 S P O 0 4 _ 2 0 0 7 L A N 6 6 _ 2 0 0 7 L N C 0 4 _ 2 0 0 7 S O U 0 9 _ 2 0 0 7 G O U 2 2 _ 2 0 0 8 F O R 3 5 _ 2 0 0 8 D U C 5 6 _ 2 0 0 8 M O N 7 4 _ 2 0 0 6 H A R 2 2 _ 2 0 0 8 M U S 3 5 _ 2 0 0 8 D R E 2 9 _ 2 0 0 8 G U E 2 2 _ 2 0 0 7 T O U 7 9 _ 2 0 0 7 V A L 3 8 _ 2 0 0 7 L L I6 6 _ 2 0 0 6 B O U 4 3 _ 2 0 0 7 S A U 3 8 _ 2 0 0 9 L L I6 6 _ 2 0 0 7 L A F 3 8 _ 2 0 0 9 B O U 4 3 _ 2 0 0 6 R D V 3 5 _ 2 0 0 8 R O S 7 3 _ 2 0 0 7 S E R 6 3 _ 2 0 0 7 A L L 0 4 _ 2 0 0 5 P E T 3 8 _ 2 0 0 9 B IR 0 1 _ 2 0 0 8 L A C 2 1 _ 2 0 0 6 A U L 1 3 _ 2 0 0 8 E N T 2 5 _ 2 0 0 8 P A R 1 2 _ 2 0 0 7 P U I1 8 _ 2 0 0 7 C H O 0 4 _ 2 0 0 9 L D C 2 5 _ 2 0 0 7 C H A 2 3 _ 2 0 0 6 O S P 2 A _ 2 0 0 8 R R T 2 3 _ 2 0 0 6 C A R 6 6 _ 2 0 0 9 S P O 2 5 _ 2 0 0 9 R G M 3 8 _ 2 0 0 8 V IN 6 6 _ 2 0 0 9 C H A 4 8 _ 2 0 0 7 A V E 3 4 _ 2 0 0 9 E S P 0 4 _ 2 0 0 7 L R O 3 9 _ 2 0 0 8 A L E 2 B _ 2 0 0 7 M O N 6 3 _ 2 0 0 6 C A S 0 4 _ 2 0 0 8 T O L 2 A _ 2 0 0 7 C H A 3 9 _ 2 0 0 7 E G U 3 6 _ 2 0 0 7 M O N 3 8 _ 2 0 0 9 M O N 7 4 _ 2 0 0 7 C H E 7 3 _ 2 0 0 9 V O U 3 9 _ 2 0 0 8 P A L 3 8 _ 2 0 0 8 S C A 8 3 _ 2 0 0 7 B L E 6 5 _ 2 0 0 7 E T I3 9 _ 2 0 0 5 G R A 4 2 _ 2 0 0 7 M O N 6 3 _ 2 0 0 7 C H A 3 9 _ 2 0 0 6 R S T 3 5 _ 2 0 0 8 L G M 3 9 _ 2 0 0 5 IL A 3 9 _ 2 0 0 9 L A U 3 8 _ 2 0 0 7 S E R 6 3 _ 2 0 0 6 V IL 3 5 _ 2 0 0 7 A N T 7 4 _ 2 0 0 7 E T I3 9 _ 2 0 0 6 G L C 3 9 _ 2 0 0 5 G L C 3 9 _ 2 0 0 9 M A T 6 6 _ 2 0 0 9 R B J 3 5 _ 2 0 0 8 B IS 7 3 _ 2 0 0 9 R O C 0 3 _ 2 0 0 6 26R/1LN 23R/15LN 29R/21LN D is ta n c e d e B ra y e t C u rt is 0 2 4 6 8 hclust T U I4 5 _ 2 0 0 6 L A V 2 3 _ 2 0 0 7 R L B 1 1 _ 2 0 0 9 S C R 0 4 _ 2 0 0 7 R S V 3 4 _ 2 0 0 7 E S T 1 1 _ 2 0 0 7 T Y X 6 3 _ 2 0 0 6 T R E 3 5 _ 2 0 0 6 R IN 5 3 _ 2 0 0 6 S F P 5 3 _ 2 0 0 6 R G L 6 9 _ 2 0 0 9 A P R 8 5 _ 2 0 0 7 V IO 4 4 _ 2 0 0 6 A R Z 3 5 _ 2 0 0 7 P R O 4 4 _ 2 0 0 6 C A E 3 5 _ 2 0 0 6 N O Y 5 6 _ 2 0 0 7 E P T 3 5 _ 2 0 0 6 C A Z 4 0 _ 2 0 0 5 B L I3 6 _ 2 0 0 6 P D F 4 4 _ 2 0 0 6 C A R 3 5 _ 2 0 0 6 O U E 3 5 _ 2 0 0 6 B A Y 5 8 _ 2 0 0 6 B O U 3 5 _ 2 0 0 6 J O U 1 1 _ 2 0 0 8 R E A 1 3 _ 2 0 0 9 B O S 2 2 _ 2 0 0 7 R O P 2 2 _ 2 0 0 7 B O R 3 3 _ 2 0 0 7 C A Z 4 0 _ 2 0 0 7 C O D 2 B _ 2 0 0 9 E T I3 9 _ 2 0 0 7 M O N 7 4 _ 2 0 0 5 B U L 8 5 _ 2 0 0 7 E C H 3 3 _ 2 0 0 7 B O U 6 6 _ 2 0 0 6 B O U 6 6 _ 2 0 0 7 R M N 2 9 _ 2 0 0 8 N D C 3 8 _ 2 0 0 9 S P O 0 4 _ 2 0 0 7 L A N 6 6 _ 2 0 0 7 L N C 0 4 _ 2 0 0 7 S O U 0 9 _ 2 0 0 7 G O U 2 2 _ 2 0 0 8 F O R 3 5 _ 2 0 0 8 D U C 5 6 _ 2 0 0 8 M O N 7 4 _ 2 0 0 6 H A R 2 2 _ 2 0 0 8 M U S 3 5 _ 2 0 0 8 D R E 2 9 _ 2 0 0 8 G U E 2 2 _ 2 0 0 7 T O U 7 9 _ 2 0 0 7 V A L 3 8 _ 2 0 0 7 L L I6 6 _ 2 0 0 6 B O U 4 3 _ 2 0 0 7 S A U 3 8 _ 2 0 0 9 L L I6 6 _ 2 0 0 7 L A F 3 8 _ 2 0 0 9 B O U 4 3 _ 2 0 0 6 R D V 3 5 _ 2 0 0 8 R O S 7 3 _ 2 0 0 7 S E R 6 3 _ 2 0 0 7 A L L 0 4 _ 2 0 0 5 P E T 3 8 _ 2 0 0 9 B IR 0 1 _ 2 0 0 8 L A C 2 1 _ 2 0 0 6 A U L 1 3 _ 2 0 0 8 E N T 2 5 _ 2 0 0 8 P A R 1 2 _ 2 0 0 7 P U I1 8 _ 2 0 0 7 C H O 0 4 _ 2 0 0 9 L D C 2 5 _ 2 0 0 7 C H A 2 3 _ 2 0 0 6 O S P 2 A _ 2 0 0 8 R R T 2 3 _ 2 0 0 6 C A R 6 6 _ 2 0 0 9 S P O 2 5 _ 2 0 0 9 R G M 3 8 _ 2 0 0 8 V IN 6 6 _ 2 0 0 9 C H A 4 8 _ 2 0 0 7 A V E 3 4 _ 2 0 0 9 E S P 0 4 _ 2 0 0 7 L R O 3 9 _ 2 0 0 8 A L E 2 B _ 2 0 0 7 M O N 6 3 _ 2 0 0 6 C A S 0 4 _ 2 0 0 8 T O L 2 A _ 2 0 0 7 C H A 3 9 _ 2 0 0 7 E G U 3 6 _ 2 0 0 7 M O N 3 8 _ 2 0 0 9 M O N 7 4 _ 2 0 0 7 C H E 7 3 _ 2 0 0 9 V O U 3 9 _ 2 0 0 8 P A L 3 8 _ 2 0 0 8 S C A 8 3 _ 2 0 0 7 B L E 6 5 _ 2 0 0 7 E T I3 9 _ 2 0 0 5 G R A 4 2 _ 2 0 0 7 M O N 6 3 _ 2 0 0 7 C H A 3 9 _ 2 0 0 6 R S T 3 5 _ 2 0 0 8 L G M 3 9 _ 2 0 0 5 IL A 3 9 _ 2 0 0 9 L A U 3 8 _ 2 0 0 7 S E R 6 3 _ 2 0 0 6 V IL 3 5 _ 2 0 0 7 A N T 7 4 _ 2 0 0 7 E T I3 9 _ 2 0 0 6 G L C 3 9 _ 2 0 0 5 G L C 3 9 _ 2 0 0 9 M A T 6 6 _ 2 0 0 9 R B J 3 5 _ 2 0 0 8 B IS 7 3 _ 2 0 0 9 R O C 0 3 _ 2 0 0 6 26R/1LN 23R/15LN 29R/21LN D is ta n c e d e B ra y e t C u rt is Eigenvalues 19.54 % 46.94 % d = 5 LN Retenue F1 F2
a)
b)
Figure 9. Dendrogramme de la classification hiérarchique réalisé sur le tableau faunistique des
macroinvertébrés (a) et plans F1 et F2 de l’analyse en composantes principales (ACP) réalisé sur les paramètres environnementaux (b) du jeu de données de 115 plans d’eau. Les points de prélèvements sont regroupés en fonction de leur type : R : retenues et LN : Lacs Naturels. L’histogramme en bas à droite est celui des valeurs propres de l’ACP.
3.1.3.2. Première sélection de métriques
Plusieurs auteurs décrivent différentes approches pour sélectionner des métriques (Hering, Bohmer, Haase, & Schaumburg, 2004; Pont et al., 2006). Nous proposons dans notre étude de faire une première sélection en se basant sur les corrélations de nos métriques avec les pressions disponibles. Nous n’avons alors retenu que celles qui avaient une corrélation (Spearman) significative et > 0.2 avec les pressions.
Cette approche est cependant un peu risquée dans notre cas car l’effet de la pression peut encore être masqué à cette étape par l’environnement. Néanmoins, dans le but d’avoir dans ce rapport d’avancement une première idée des métriques les plus pertinentes, nous avons maintenus cette première étape de sélection qui permet d’accélérer la procédure. L’ensemble des métriques sera considéré par la suite.
Le Tableau5 résume l’ensemble des coefficients de corrélation obtenus pour les 16 métriques sélectionnées à l’issue de cette première étape.
Tableau 5 : Valeurs des coefficients de corrélation de Spearman entre métriques sélectionnées et les
paramètres de pression. La significativité du coefficient est représenté par * (0.01<p.value<0.05) et par ** (p.value<0.01). Occupation du sol naturelle et semi-naturelle Occupation du sol naturelle et semi-naturelle, agriculture extensive
Urbanisme Agriculture Agriculture intensive Population (moyenne) (écart-type)Population Phosphore Carbone Azote
RS -0.07 -0.16 0.13 0.03 0.14 0.1 0.19 * -0.11 -0.21 * -0.19 * H -0.15 -0.23 * 0.11 0.1 0.21 * 0.11 0.13 -0.04 -0.09 -0.07 Hmin 0.02 -0.03 0.08 -0.04 0.02 0.09 0.2 * -0.17 -0.24 * -0.2 * Hmax -0.07 -0.16 0.13 0.03 0.14 0.1 0.19 * -0.11 -0.21 * -0.19 * N -0.15 -0.23 * 0.11 0.1 0.21 * 0.11 0.13 -0.04 -0.09 -0.07 DNaididae_sens 0.32 ** 0.28 ** -0.13 -0.36 ** -0.29 ** -0.35 ** -0.32 ** -0.05 -0.06 -0.13 DOtherNaid 0.24 ** 0.21 * -0.11 -0.23 * -0.19 * -0.17 -0.1 -0.12 -0.21 * -0.21 * DTub_sss -0.24 ** -0.23 * 0.07 0.25 ** 0.26 ** 0.2 * 0.2 * 0 -0.04 -0.06 Tubacs/Tusss 0.26 ** 0.23 * -0.02 -0.26 ** -0.27 ** -0.1 -0.04 -0.03 0.01 0.11 EP 0.08 0.01 0.1 -0.14 -0.07 0.05 0.05 -0.25 ** -0.18 -0.16
Espèces sensibles à la pollution
organique et toxique 0.37 ** 0.32 ** -0.15 -0.41 ** -0.33 ** -0.36 ** -0.27 ** -0.12 -0.15 -0.18 *
%Naid/OligTot -0.25 ** -0.21 * 0.12 0.24 * 0.2 * 0.18 0.12 0.12 0.21 * 0.22 *
%Naid_sens/OligTot 0.32 ** 0.28 ** -0.13 -0.37 ** -0.3 ** -0.36 ** -0.32 ** -0.05 -0.06 -0.14
%Naid_sens/NaidTot 0.32 ** 0.28 ** -0.13 -0.37 ** -0.3 ** -0.36 ** -0.32 ** -0.05 -0.06 -0.15
%Tubsssoies/OligTot -0.36 ** -0.32 ** 0.06 0.36 ** 0.35 ** 0.22 * 0.17 0.03 0.02 -0.04
%Espèces sensibles à la pollution
organique et toxique 0.38 ** 0.33 ** -0.15 -0.43 ** -0.34 ** -0.38 ** -0.3 ** -0.1 -0.13 -0.18
3.1.3.3. Résultats des modèles par métriques
Des modèles de prédiction par régression linéaire multiple avec une approche pas à pas (stepwise) utilisant le critère d’AIC pour sélectionner les variables environnementales naturelles et anthropiques explicatives du modèle ont été construits pour chacune des métriques sélectionnées. Les résultats des ces modèles (R² ajustés) sont regroupés dans le Tableau 6. La métrique Hmin ne figure plus dans le tableau car aucune pression après stepwise n’a été conservée.
Seuls les modèles qui ont obtenu un R² ajusté supérieur à 0.3 ont été utilisés pour l’approche hindcasting considérant que cette valeur était le minimum acceptable pour avoir des modèles capables de prédire au mieux la métrique.
Ce critère a pour conséquence une nouvelle sélection des métriques qui ne sont plus que 4 à cette étape : DTub_sss, Tubacs/Tusss, %Tubsssoies/OligTot, %Espèces sensibles à la pollution organique et toxique.