CHRISTINE GIRARD
(JLL
6^/7
LATÉRALISATION DES PROCESSUS TEMPORELS :
SÉQUENCES RYTHMIQUES VISUELLES
Mémoire Présenté
à la Faculté des études supérieures de !’Université Laval
pour l’obtention
du grade de maître en psychologie (M. Ps.)
École de psychologie
FACULTÉ DES SCIENCES SOCIALES UNIVERSITÉ LAVAL
AOÛT 2003
Une expérience portant sur latéralisation des processus temporels tente de vérifier s'il existe des asymétries entre les hémisphères cérébraux en utilisant des séquences rythmiques visuelles. L'hémisphère gauche (HG) utiliserait un mode analytique pour faire traiter !'information reçue tandis que l'hémisphère droit (HD) utiliserait un mode holistique. S’il est possible de faire correspondre un petit nombre d'intervalles de temps (2) à un traitement analytique, l'HG devrait avoir une performance supérieure. Si un grand nombre d'intervalles (4) peut correspondre à un traitement holistique, c'est l'HD qui devrait avoir une performance supérieure. L'examen de ces variables se fait à l'aide de séquences de 2 et de 4 intervalles de temps standards ayant des durées de 900 ms, 1125 ms et 1406 ms. Les résultats démontrent que l'HD est significativement meilleur dans les séquences de 4 intervalles que dans celles de 2 et que l’HD a une performance plus élevée que l'HG lorsque les séquences comportent 4 intervalles de temps.
---y---Simon Grondin Directeur de recherche Christine Girard
Tout d’abord, je tiens à remercier mon directeur de recherche, Simon Grondin, pour son
encadrement et sa supervision tout au long de ce projet. Je tiens également à remercier ma famille et tous les gens qui m’entourent pour leur soutien et leurs encouragements au cours de ces deux dernières années. Grâce à tous ces gens, j’ai pu trouver l’énergie et la motivation nécessaire à T accomplissement de cet important travail de recherche. Finalement, je souhaite remercier le CRSNG pour le support financier qui m’a été accordé tout au long de mes études de maîtrise.
pages
RÉSUMÉ...:...ü
AVANT-PROPOS... iii
TABLE DES MATIÈRES... iv
LISTE DES TABLEAUX...vi
LISTE DES FIGURES... vii
INTRODUCTION GÉNÉRALE...1 CONTEXTE THÉORIQUE...2 Latéralité... 2 Différences hémisphériques... 2 Différences anatomiques... 2 Différences fonctionnelles...2 Communication interhémisphérique... 3
Voies visuelles vs Voies auditives... 4
Distinction verbale / non-verbale...5
Distinction analytique / holistique...7
Perception temporelle...8
Psychophysique et séquences rythmiques temporelles... 8
Méthodologie en perception du temps... 9
Latéralité et processus temporels... 10
L’HG et le temps... 10 Perception musicale... 12 Perception rythmique...12 La présente étude... 15 Hypothèses... 15 MÉTHODE... 17 Participant...17 Matériel... 17 Procédure... 17 iv
Comparaison des moyennes...19 Hypothèse 1...19 Hypothèse 2... 20 Erreur constante... 21 DISCUSSION... ... 22 Seuil différentiel... 22
Distinction analytique/ho Estique...23
Modèle temporel... 24 Loi de Weber... 24 Erreur constante... 25 Recommandation... 26 CONCLUSION...28 RÉFÉRENCES... 29 ANNEXE A : Tableaux... 33 ANNEXE B : Figures... 38
ANNEXE C : Formulaire de consentement...45
ANNEXE D : Tableaux des analyses de la variance... 47
ANNEXE E : Questionnaire sur la préférence manuelle...53
R2 pour chacun des participants à chacune des douze conditions à l’étude
Moyenne (M) et erreur-type (ET) du point d’égalité subjective (PES), du seuil différentiel (SD) et de la fraction de Weber (FW) des douze participants aux douze conditions à l’étude
Moyenne (M) et erreur-type (ET) du point d’égalité subjective (PES), du seuil différentiel (SD) et de la fraction de Weber (FW) des douze participants en fonction du champ visuel et de la longueur des standards.
Moyenne (M) et erreur-type (ET) du point d’égalité subjective (PES), du seuil différentiel (SD) et de la fraction de Weber (FW) des douze participants en fonction du champ visuel et du nombre de standards
Moyenne (M) et erreur-type (ET) du point d’égalité subjective (PES), du seuil différentiel (SD) et de la fraction de Weber (FW) des douze participants pour le champ visuel
Moyenne (M) et erreur-type (ET) du point d’égalité subjective (PES), du seuil différentiel (SD) et de la fraction de Weber (FW) des douze participants pour le nombre de standards
Moyenne (M) et erreur-type (ET) du point d’égalité subjective (PES), du seuil différentiel (SD) et de la fraction de Weber (FW) des douze participants pour la longueur des standards
Tableau 1. Tableau 2. Tableau 3. Tableau 4. Tableau 5. Tableau 6. Tableau 7. VI
Seuil différentiel en fonction du champ visuel et du nombre d’intervalles standards.
Seuil différentiel en fonction de la longueur de l’intervalle standard.
Fraction de Weber en fonction du champ visuel et du nombre d’intervalles standards.
Fraction de Weber en fonction de la longueur de l’intervalle standard, du champ visuel et du nombre d’intervalles standards.
Fraction de Weber en fonction de la longueur de l’intervalle standard.
Fraction de Weber en fonction de la longueur de !’intervalle standard et du champ Figure 1. Figure 2. Figure 3. Figure 4. Figure 5. Figure 6. visuel. vu
LATÉRALISATION DES PROCESSUS TEMPORELS : SÉQUENCES RYTHMIQUES VISUELLES
INTRODUCTION GÉNÉRALE
Élément fondamental, le temps influence la façon dont les êtres humains perçoivent et s’adaptent aux choses qui les entourent. Depuis quelques décennies, l’information temporelle reçoit une attention particulière de plusieurs chercheurs afin de déterminer comment celle-ci est traitée par le cerveau et, plus précisément, par chacun de ses hémisphères. Même si la plupart des études scientifiques rapportent une analyse temporelle supérieure dans l’hémisphère gauche (HD), la latéralisation de ce processus suscite encore de la controverse (Nicholls, 1996). En effet,
l’analyse temporelle pourrait être possible dans les deux hémisphères cérébraux (Grondin, 1998).
L’analyse du temps à l’aide de séquences rythmiques serait favorisée dans l’hémisphère gauche. En effet, plusieurs auteurs rapportent que l’hémisphère gauche serait supérieur à l’hémisphère droit (HD) pour la discrimination de quelques formes temporelles de séquences rythmiques auditives (Springer & Deutsch, 1998 ; Nicholls, 1996, Carmon & Nachshon, 1971, Robinson & Solomon, 1974 ; Gordon, 1978, Bradshaw & Nettleton, 1981) et visuelles (Carmon & Nachshon, 1971 ; Carmon, 1979). Ceci étonne si on considère que le rythme est une composante musicale et que l’HD serait supérieur au gauche dans le traitement de !’information musicale (Kallman & Corballis, 1975 ; Kimura, 1961, Springer & Deutsch, 1998). Toutefois, selon Bradshaw et Nettleton (1981), l’HD n’est pas toujours ou dans tous ses aspects, dominant pour la musique.
Ce mémoire vise à déterminer s’il existe des asymétries pour le traitement de !’information temporelle dans la modalité visuelle et ce, en utilisant une tâche de discrimination d’intervalles temporels. Pour effectuer cette vérification, des séquences rythmiques visuelles composées de deux ou quatre intervalles de temps standards (900, 1125 ou 1406 ms) séparés respectivement par trois ou cinq stimuli lumineux seront utilisées. Ces séquences seront présentées dans le champ visuel droit (CVD) ou dans le champ visuel gauche (CVG) afin de vérifier s’il existe des différences significatives entre ces deux champs visuels.
CONTEXTE THÉORIQUE
Tout d’abord, le contexte théorique expose les principes majeurs de latéralité retrouvés dans la littérature. Ensuite, certains aspects de la perception du temps sont discutés. Finalement, le lien entre la latéralité et les processus temporels sont abordés certains aspects de la perception du temps sont discutés.
Latéralité
Différences hémisphériques
Différences anatomiques. L’ensemble de l’organisme humain est divisé en deux parties et il
en n’est pas autrement avec le cerveau. Les deux hémisphères qui le composent paraissent semblables, mais ceux-ci diffèrent au plan anatomique et ils le sont particulièrement au niveau fonctionnel. Braun (2000) dresse un portrait récapitulatif des études ayant démontré une
asymétrie hémisphérique humaine au niveau anatomique. Entre autre, les asymétries favorisant l’hémisphère gauche sont : plus de substance grise et lobe occipital plus large et celles favorisant l’hémisphère droit sont : plus lourd et lobe frontal plus gros. De plus, presque toutes les aires du cerveau ont tendance à avoir un volume différent des deux côtés.
Différences fonctionnelles. Étant donné que l’HG possède davantage les fonctions mentales
jugées importantes dans les sociétés industrialisées, il était autrefois, et même encore aujourd’hui, qualifié d’hémisphère dominant (Springer & Deutsch, 1998). L’HD, quant à lui, qualifié
d’hémisphère mineur ou silencieux, ne semblait pas posséder de fonctions spéciales et il était considéré comme étant subordonné au contrôle de l’hémisphère dominant (gauche) (Lezak,
1995). John Hughlings Jackson a été un des premiers à considérer que l’HD possédait certaines fonctions importantes (Springer & Deutsch, 1998). Aujourd’hui, plusieurs fonctions sont associées à l’HD et ce, grâce au plus récent intérêt que plusieurs chercheurs lui portent.
L’intérêt général pour les différences fonctionnelles entre les deux hémisphères origine des observations faites au XIXe siècle sur des patients ayant des atteintes cérébrales unilatérales (Ivry & Robertson, 1998). Depuis ce temps plusieurs recherches sur la latéralisation ont été menées. Entre autre, Keller et Sutton, (1991) associent l’aphasie, les difficultés reliées à la mémoire
verbale, les problèmes de fluence verbale, la pensée concrète et des déficits de la lecture et de l’écriture aux lésions de l’hémisphère gauche. Quant aux patients ayant des lésions à
l’hémisphère droit, ils sont fluents, illogiques, aptes aux généralisations exagérées et aux mauvais jugements, ont de la difficulté à planifier, à bien comprendre des situations complexes et à bien
traiter les composantes musicales.
Chez environ 90% des gens les habiletés du langage, les habiletés mathématiques et la logique se situent dans l’HG. Quant aux habiletés visuo-spatiales, l’intuition, l’émotion, l’appréciation de l’art et de la musique, ainsi que la reconnaissance des visages, elles seraient situées dans l’HD (Marieb, 1993). Selon Ivry et Robertson (1998), les régions de l’HG jouent clairement un rôle dominant dans le langage tandis que les régions de l’HD sont plus utiles pour se déplacer dans un monde tridimensionnel. Même s’ils considèrent qu’il existe un continuum des fonctions plutôt qu’une dichotomie rigide entre les hémisphères cérébraux, Bradshaw et Nettleton (1981)
associent l’HG à la discrimination impliquant la durée, l’ordre temporel, l’analyse des séquences et le rythme et ce, au niveau sensoriel (tactile, visuel et auditif) et plus spécialement au niveau moteur (les doigts, les membres et l’appareil langagier). De plus, cet hémisphère serait associé aux fonctions analytiques. Quant à l’HD, il serait caractérisé par les aspects spatiaux : la cartographie de l’espace autour du corps et sa position. En plus, il analyserait les stimuli de manière synthétique et holistique.
Communication interhémisphérique
Les deux hémisphères cérébraux peuvent communiquer entre eux par de nombreuses
commissures, principalement à l’aide du corps calleux. Lorsque l’information parvient dans l’hémisphère non spécialisé dans le traitement de ce type d’information, celle-ci est transférée dans l’hémisphère spécialisé. L’analyse du stimulus est plus longue lorsque !’information est transférée d’un hémisphère à l’autre en raison du voyage qu’elle a à parcourir. En effet, les situations qui requièrent un transfert d’information d’un hémisphère à un autre pour obtenir une réponse ont des temps de réaction plus longs comparativement aux situations où aucun transfert n’est nécessaire (Davis & Schmit, 1971).
Voies visuelles vs Voies auditives
La modalité par laquelle un stimulus est produit détermine la façon dont celui-ci se rendra au cerveau. En effet, si le stimulus est de type visuel son cheminement vers le cerveau sera différent que s’il est de type auditif. Autrefois, certains théoriciens croyaient que l’HG était lié à l’audition et que l’HD était en lien avec les stimuli visuels (Lezak, 1995). Aujourd’hui, nous savons que ces associations sont fausses puisque chaque hémisphère peut traiter autant l’information visuelle qu’auditive. Même qu’en perception auditive, il est possible de voir des principes de
spécialisation hémisphérique similaire à ceux existant en perception visuelle (Ivry & Robertson, 1998). Concrètement, un avantage de l’HG est observé si la performance est supérieure lors de la présentation d’un stimulus à l’oreille droite et dans le champ visuel droit (CVD). Inversement, un avantage de l’HD est démontré lorsque un stimulus est présenté à l’oreille gauche et dans le champ visuel gauche (CVG) (Springer & Deutsch, 1998). Pour pouvoir étudier les asymétries fonctionnelles cérébrales des participants normaux, il existe des techniques spécifiques pour chacune des modalités sensorielles.
Sur le plan visuel, la technique utilisée pour étudier la latéralité est celle des champs visuels divisés. Elle consiste à faire clignoter un stimulus lumineux très brièvement soit à la gauche ou à la droite d’un point fixé par le participant. De cette manière, les chercheurs peuvent s’assurer que le stimulus est présenté à un seul hémisphère (Springer & Deutsch, 1998). L’organe récepteur de l’information visuelle est la rétine. Celle-ci se divise en deux parties : la partie temporale et la partie nasale. Les informations provenant de la partie temporale de la rétine se rendent de façon ipsilatérale à l’hémisphère cérébral correspondant tandis que les informations de la rétine nasale s’y dirigent plutôt de manière controlatérale, croisant au niveau du chiasma optique.
L’information qui est au centre du champ visuel est projetée sur les parties temporales de la rétine de chaque œil pour se rendre de façon équivalente aux deux hémisphères cérébraux.
L’information qui se situe à gauche dans le champ visuel est dirigée vers l’HD. Pour ce qui est de l’information située dans la partie droite du champ visuel, celle-ci est transmise à l’HG. Selon White (1969), les différences latérales en perception visuelle dépendent de plusieurs facteurs : • Type de présentation du stimulus : unilatéral ou bilatéral
• Quantité, nature et espacement des éléments stimulus-information • Intensité à laquelle l’information est présentée
• Ordre dans lequel l’information est rapportée
• Conditions visuelles employées et dominance oculaire des participants • Manualité et latéralisation des participants
Toutefois, selon Nicholls (1994), les asymétries observées dans les recherches à l’aide de champs visuels divisés auraient peu de hens avec l’asymétrie cérébrale sous-jacente.
Le cheminement de !’information s’effectue différemment au niveau auditif. En effet, toute !’information qui parvient à une oreille est retransmise aux deux hémisphères cérébraux, mais pas de manière équivalente. La voie auditive croisée reliant l’oreille d’un côté avec l’hémisphère de l’autre côté est plus importante dans le transfert d’information que la voie auditive non croisée reliant l’oreille d’un côté avec l’hémisphère du même côté (Kimura, 1961). Lorsque
Γ information arrive également aux deux oreilles, les hémisphères cérébraux reçoivent les mêmes données. Ainsi, en situation normale où les deux oreilles ont accès à la même information, il est difficile d’identifier la supériorité d’une oreille pour un type précis d’information. Avec la technique d’écoute dichotique proposée par Kimura, il est possible de déterminer qu’un type d’information est favorisé dans un hémisphère en particulier. En effet, chaque oreille reçoit un ensemble de stimuli différents et la personne doit rapporter ce qu’elle a entendu.
Distinction verbale/non-verbale
Quand les chercheurs essaient de comprendre comment les deux hémisphères fonctionnent ensemble, ils doivent garder à l’esprit la nature des stimuli. Ils ne doivent pas seulement savoir quelle voie sensorielle ils utiliseront, mais aussi la nature verbale ou non-verbale de !’information qui sera analysée (Milner, 1962). En effet, la différence fonctionnelle la plus évidente entre les hémisphères cérébraux est que l’HG, pour la plupart des gens, est spécialisé pour le langage et que FHD est spécialisé pour les stimuli difficiles à verbaliser (Lezak, 1995).
Il a été démontré à maintes reprises dans la littérature que l’HG est associé au langage. C’est au milieu du XIXe siècle que Broca et Wernicke ont clamé le rôle verbal de l’HG (Bradshaw & Nettleton, 1981). Le lien entre le langage et l’HG s’explique en partie par le fait que les désordres du langage ou les aphasies sont un signe fondamental d’un dommage à l’HG (Ivry & Robinson, 1998). De plus, les déficits produits par une lobectomie temporale gauche sont verbaux (Milner,
1962). La présentation de mots ou simplement de lettres résulte en une supériorité du CVD. Les stimuli n’ont pas besoin d’être significatifs pour produire une supériorité de l’HG, mais ils doivent être reliés au langage ou être verbal d’une certaine manière (Springer & Deutsch, 1998). Selon Lezak (1995), l’HG serait spécialisé pour les fonctions verbales incluant la lecture,
l’écriture, la compréhension, la parole, l’idéation verbale, la mémoire verbale et même les symboles verbaux tracés sur la peau.
Dans une expérience menée par Kimura (1961), 120 sujets ayant des lésions épileptogènes au cerveau ont été utilisés : 107 avaient le langage situé dans l’HG et 13 dans l’HD. Ceux-ci recevaient, à l’aide d’écouteurs, six chiffres dans l’oreille droite et six autres chiffres dans l’oreille gauche par la méthode d’écoute dichotique. Ensuite, ils devaient rapporter ce qu’ils avaient entendu dans l’ordre qu’ils désiraient. Les résultats ont démontré que lorsque des stimuli verbaux étaient présentés aux deux oreilles, les stimuli qui arrivaient à l’oreille opposée à
l’hémisphère dominant étaient reconnus plus efficacement. Bref, l’oreille droite était plus efficace que la gauche quand le langage était représenté dans l’HG et l’oreille gauche était plus efficace lorsque le langage était situé dans l’HD.
Parmi les stimuli visuels non-verbaux qui amènent une supériorité de l’HD, il y a les visages et les arrangements spatiaux de points. Les études d’écoute dichotique qui résultent en une
supériorité de l’HD sont diverses et ont utilisé des stimuli tels que des extraits de mélodie et des sons environnementaux (Springer & Deutsch, 1998). De plus, la forme, la texture et les motifs sont des stimuli non-verbaux favorisant PHD (Lezak, 1995). Selon Milner (1962), les pertes suivant une lobectomie temporale droite chez l’homme suggèrent que le lobe droit joue un rôle majeur dans les fonctions visuelles non-verbales et dans les fonctions auditives, tandis que le lobe temporal gauche ne joue qu’un rôle auxiliaire dans ces fonctions. Il semblerait que l’HD jouerait quand même un rôle au niveau du langage. Il démontrerait une compréhension de certains mots, spécialement des noms d’objets et il pourrait apporter une contribution unique à nos habiletés de communication (Springer & Deutsch, 1998 ; Lezak, 1995).
Selon Nicholls (1996), malgré les preuves en faveur d’une dichotomie verbale/non-verbale, certaines recherches suggèrent que cette dichotomie amène seulement une explication limitée des
capacités hémisphériques de traitement de l’information. En fait, la spécialisation hémisphérique est plus flexible. Ainsi, sous certaines circonstances, l’HD est capable d’accomplir certaines fonctions verbales et l’HG est en mesure de traiter certains stimuli non-verbaux quand les conditions le permettent. Une expérience (Nicholls & Cooper, 1991) menée auprès de 22 sujets normaux avait pour objectif de déterminer s’il existait une différence entre les hémisphères cérébraux en ce qui concerne la vitesse de traitement de !’information et ce, à l’aide d’une tâche d’inspection temporelle qui utilisait des stimuli non-verbaux. Les résultats de cette recherche ont démontré que les participants obtenaient de meilleurs scores lorsque les stimuli étaient présentés dans le CVD plutôt que dans le CVG. Ainsi, l’avantage du CVD-HG ne peut pas s’expliquer par sa spécialisation pour l’analyse du langage puisque les stimuli étaient de nature non-verbale et que l’expérience ne nécessitait pas de réponse verbale. L’auteur explique ces résultats par les capacités différentes des hémisphères à traiter !’information à un niveau plus fondamental. Donc, il est possible d’observer certaines exceptions à la dichotomie verbale/non-verbale.
Distinction analytique/holistique
Puisque la dichotomie verbale/non-verbale ne semble pas en mesure d’expliquer entièrement les résultats des recherches dans le domaine de la latéralité, il s’est avéré important pour les
chercheurs de trouver une autre façon de catégoriser les fonctions dans chaque hémisphère. Dans la littérature, il a souvent été mentionné que l’HG est spécialisé pour les fonctions analytiques et détaillées tandis que F HD peut procurer une image globale du monde puisqu’il est spécialisé pour les fonctions holistiques (Efron, 1990). Selon Bryden (1982), la plupart des tâches qui produisent un effet de l’HD peuvent être considérées intégratives ou holistiques : tâches visuo- spatiales, musique, sons environnementaux, reconnaissance tactile de formes, émotions et reconnaissance des visages. Parallèlement, la plupart des tâches liées à l’HG sont largement analytiques. D’un côté, les lésions à l’HG semblent être associées à des problèmes d’organisation séquentielle où il faut être capable de placer des objets dans certaines catégories conceptuelles ou abstraites. De plus, ces lésions seraient liées à de très grandes simplifications ou au manque de détails dans le dessin tout en conservant la forme et le contour général de ces dessins. De l’autre côté, les lésions à l’HD amènent la personne à une trop grande attention aux détails, à un manque de conscience de !’organisation totale d’une situation et à de la difficulté à se servir d’une carte routière (Bradshaw & Nettleton, 1981).
Selon Iaccino (1993), une des premières fonctions du mode analytique-gauche est de placer les stimuli importants de façon sérielle, bref de les disposer dans le bon ordre temporel. En effet, Γagencement de ces stimuli selon une séquence particulière est très important. Ce qui arrive premier, second ou troisième fait une différence dans la manière de traiter !’information et de la conserver en mémoire. En ce qui concerne le mode holistique-droit, il est très habile à saisir les patrons de relations qui existent entre les composantes d’un ensemble. Il intègre plusieurs données simultanément de façon parallèle pour finalement obtenir une configuration complète comme le veut le principe de la Gestalt. Il n’ordonne pas les éléments de façon temporelle.
Les travaux de Semmes (1968) ont apporté un grand soutien à la dichotomie
analytique/holistique. Elle a observé que les dommages à l’HG amenaient des déficits précis à la main droite tandis que les dommages à l’HD amenaient, souvent aux deux mains, des déficits plus généraux et moins sévères. Ainsi, elle a associé une représentation corticale plus focalisée à l’HG et une représentation corticale plus diffuse à l’HD. En effet, les habiletés spatiales et topographiques sont représentées de manière plus diffuse dans l’HD, permettant de former un tout à partir de plusieurs informations sensorielles. Inversement, les habiletés motrices et
langagières sont représentées de façon plus focalisée dans l’HG, accordant à l’humain la capacité de se servir habilement de ces fonctions.
Perception temporelle
Psvchophvsique et séquences rythmiques temporelles
La psychophysique est un domaine d’étude qui combine des phénomènes sensoriels cognitifs à la réalité physique. De cette façon, elle permet l’analyse de certains aspects du fonctionnement cognitif comme !’évaluation et l’estimation par rapport à un continuum physique, le temps. Pour étudier le temps psychologique, il existe plusieurs méthodes expérimentales. La loi de Weber constitue un outil très utile quant à l’analyse des résultats en psychophysique (Grondin, 2001b). Dans sa forme rigoureuse, la loi de Weber établit que le seuil différentiel, qui est la différence d’amplitude minimale entre deux stimuli pour les distinguer, augmente
proportionnellement selon l’ampleur du stimulus standard. Bref, le ratio entre le seuil différentiel et le stimulus standard est constant, constituant la fraction de Weber. Ce principe devrait
stimulus, la fraction de Weber est plus élevée. Ce phénomène semble se répéter pour certaines valeurs plus élevées.
La perception temporelle est étudiée dans une recherche de Grondin (2001a) qui emploie une technique de discrimination de séquences rythmiques visuelles continue et discontinue. Avec la méthode continue, une série de 2-4 intervalles de temps séparés par 3-5 stimuli visuels
respectivement sont projetés au centre du champ visuel du participant et celui-ci doit indiquer si le dernier intervalle entre les deux derniers stimuli visuels était plus court ou plus long que les intervalles précédents. Tandis qu’avec la méthode discontinue, la série de 1-3 intervalles de temps séparés par 2-4 stimuli visuels respectivement est coupée en deux parties. Le participant doit donc indiquer si les intervalles entre les stimuli de la deuxième partie de la série sont plus courts ou plus longs que ceux de la première série. Deux standards seulement étaient à l’étude dans chacune des expériences 2 et 3 de Grondin (2001a) soit 600 et 900 ms et 900 et 1200 ms respectivement. Les résultats de cette recherche concernant la fraction de Weber montrent que celle-ci demeure constante pour 600 et 900 ms mais qu’elle augmente entre 900 et 1200 ms. Puisque la constante de Weber est plus haute à 1200 ms qu’à 900 ms, il est possible d’affirmer que la loi de Weber subit une violation pour cette valeur. Les résultats obtenus indiquent aussi que !’augmentation de la présentation du standard améliore la performance des participants dans la méthode discontinue mais pas dans la méthode continue. En d’autres mots, la discrimination est meilleure dans la condition discontinue lorsque davantage d’intervalles sont utilisés. Dans la condition continue, l’effet du nombre de standards dépend de la durée du standard.
Méthodologie en perception du temps
Il existe plusieurs méthodes pour étudier la latéralité au niveau de la perception temporelle. Parmi celles-ci se retrouve la tâche de discrimination d’intervalles temporels qui consiste pour le
participant à déterminer si un intervalle est plus court ou plus long qu’un intervalle présenté précédemment (voir Grondin, 2001a).
Il est possible de présenter un intervalle sous deux formes : plein ou vide. Un intervalle plein est un signal continu qui constitue la durée à estimer. Un intervalle vide est la durée estimée entre la fin d’un premier signal et le début d’un deuxième signal. La présentation des stimuli peut aussi se
faire de deux façons. Premièrement, de manière unilatérale soit d’un seul côté du champ visuel ou seulement dans une oreille et deuxièmement, elle peut se faire de manière bilatérale soit dans les deux côtés du champ visuel ou dans les deux oreilles en même temps. Les stimuli peuvent être présentés sous plusieurs modalités. Les modalités qui sont le plus souvent utilisées par les chercheurs sont visuelles et auditives. Selon Ben-Dov et Camion (1984), certaines raisons
privilégient !’utilisation de rythmes en mode visuel comparativement à celle de rythmes en mode auditif. En effet, il est possible que les résultats des présentations auditives soient confondus en raison de leur association avec les séquences de code morse ou avec les mélodies. Le mode de présentation visuel éviterait ces ambiguïtés, reflétant seulement la spécialisation relative de l’hémisphère traitant le rythme.
Latéralité et processus temporels
L’HG et le temps
Plusieurs recherches démontrent que le traitement temporel est associé à l’HG, Entre autre, Mills et Roliman (1980) ont développé un modèle de l’asymétrie cérébrale qui propose que le jugement de l’ordre temporel se fait principalement dans l’HG. Davis et Wada (1977) affirment que l’HG analyse !’information ordonnée de façon temporelle. Selon Polzella, DaPolito et Hinsman (1977), il est évident que les deux hémisphères répondent différemment aux manipulations temporelles. En utilisant des ensembles de stimuli lumineux, ils ont découvert que la moyenne de la durée perçue pour les stimuli présentés dans le CVG-HD était significativement inférieure à la moyenne de la durée perçue des stimuli présentés dans le CVD-HG. Puisque la durée était sous-estimée dans les deux hémisphères, il est possible de dire que c’est l’HG qui avait la meilleure
approximation de la durée. De plus, selon Nicholls et Cooper (1991) l’HG analyserait !’information plus rapidement que l’HD.
L’HD est spécialisé au niveau de la perception spatiale tandis que la spécialisation de l’HG se situe au niveau de la perception temporelle. Ainsi, la division du travail entre les deux
hémisphères se fait selon les différentes dimensions perceptives. Le fait que l’HG soit dépendant de la dimension temporelle implique qu’une série de stimuli comme les mots dans une phrase, les objets dans un ensemble, les éléments d’une photo, les segments d’un film soient perçus,
perception temporelle serait interreliée avec la perception langagière, toutes deux traitées par l’HG. Toutefois, la perception temporelle ne dépend pas du langage, ce serait plutôt la perception langagière qui dépendrait d’une analyse séquentielle de l’ordre temporel (Carmon & Nachshon, 1971). Selon Davis et Wada (1977), il existerait une complémentarité au niveau du traitement temporel de l’hémisphère dominant et du traitement spatial de l’hémisphère non-dominant pour l’information afférente. Toutefois selon Vroon, Timmers et Tempelaars (1977), l’HD serait plus incertain par rapport au temps écoulé ce qui rendrait ses estimations peu fiables.
Thomas et Weaver (1975) ont proposé un modèle mathématique du temps et de la perception qui proposerait que la durée perçue d’un bref stimulus visuel serait fonction de deux processeurs, une horloge et un processeur d’information visuelle. L’attention serait divisée entre ces processeurs de sorte que si !’information visuelle augmente, plus d’attention sera accordée au processeur d’information visuelle. Par contre, si !’information visuelle diminue, plus d’attention sera donnée à l’horloge. La durée perçue serait donc une moyenne pondérée des données de ces deux
processeurs. Polzella et al. (1977) ont appliqué ce modèle de la perception temporelle à leurs données et cela leur a permis de suggérer que l’HG s’appuie sur une horloge pour estimer la durée tandis que l’HD s’appuie sur un processeur d’information visuelle pour estimer la durée. D’une part, les données présentent une meilleure précision dans le CVD-HG avec un petit nombre de données visuelles parce que !’attention relative à l’horloge est supérieure lorsque !’information visuelle est réduite. D’autre part, la précision est supérieure dans le CVG-HD lorsque le nombre de données visuelles augmente puisque !’attention relative au processeur d’information visuelle est meilleure lorsque !’information visuelle augmente.
Grondin (1998) rapporte des résultats qui ne supportent pas l’hypothèse de l’analyse temporelle par l’HG. L’expérience 1 de cette étude comprenant des intervalles de temps de 250 ms et des signaux projetés à 18° dans le champ visuel, démontre que la discrimination temporelle est légèrement meilleure dans le CVG que dans le CVD. D’autres résultats des expériences 1 et 2 n’appuient pas la spécialisation de l’HG pour le traitement temporel. En effet, lorsque des stimuli sont présentés dans les deux champs visuels successivement, soit de façon gauche-droite ou de façon droite-gauche, le mécanisme estimant le temps devrait être mis en marche plus tôt, c’est-à- dire lorsque le premier stimulus est présenté dans le CVD puisqu’il atteint directement l’HG. De
la même manière, l’activité du mécanisme temporel devrait se terminer plus tard lorsque le deuxième stimulus est présenté dans le CVG puisque la transmission de l’information du CVG vers l’HG prend un certain temps (Efron, 1990). Ainsi, il serait possible de prédire une durée perçue plus longue pour des séquences droite-gauche plutôt que pour des séquences gauche- droite. Toutefois, les résultats ne confirment pas cette hypothèse.
Perception musicale
La performance musicale, une habileté qui nécessite une précision temporelle, a souvent été associée à l’HD (Bradshaw & Nettleton, 1981). Par exemple, les sons musicaux présentés à l’oreille gauche sont traités plus rapidement et plus précisément que ceux présentés à l’oreille droite (Kallman & Corballis, 1975). Une étude (Wilson, Pressing & Wales, 2002) tente de modéliser les composantes des fonctions rythmiques en se basant sur un sujet droitier musicien amateur ayant acquis un trouble rythmique suite à un infarctus temporopariétal droit. Les résultats indiquent que le fonctionnement du cortex auditif temporal droit est fondamental pour être capable de conserver le rythme au niveau de la musique. La nature de !’information auditive musicale pose un problème en ce qui concerne la médiation exclusive de la musique par l’HD. En effet, les fonctions musicales ne sont pas strictement et entièrement latéralisées dans l’HD
(Bradshaw & Nettleton, 1981).
Des études récentes en neuropsychologie, par Doppler transcrânien sonographique, par
tomographie par émission de positron et par résonance magnétique fonctionnelle nucléaire ont indiquées que la perception musicale n’est pas dépendante de l’hémisphère droit mais d’un circuit neuronal correspondant aux composantes fondamentales de la musique dans les deux
hémisphères. Dans le cerveau, il n’y aurait pas de centre pour la musique (Baeck, 2002). Selon Vroon et al. (1977), la perception musicale serait une exception au processus de latéralisation gauche-droite. Elle devrait être mieux analysée par l’HG puisqu’elle consiste en une série ordonnée de tons comme le langage.
Perception rythmique
L’apparition régulière d’une série d’éléments en ce qui concerne le temps a été décrite comme un phénomène rythmique. Il existe des résultats contradictoires concernant l’asymétrie
hémisphérique au niveau de la perception du rythme (Ben-Dov & Carmon, 1984). Ces
contradictions proviennent du fait que PHG est spécialisé dans le traitement du langage tandis que l’HD est spécialisé dans le traitement des mélodies, deux traitements dans lesquels le rythme est une composante majeure. Certains modèles théoriques prédisent que P hémisphère qui est spécialisé dans le traitement du langage (HG) est le même que celui qui traite le rythme et ce, en dépit du fait que plusieurs recherches ont établi que le traitement de stimuli non-verbaux se faisait dans PHD. Il s’avérait que le langage et le rythme requerraient une organisation hiérarchique et que PHG serait spécialisé dans ce type de traitement (Robinson & Solomon, 1974).
Une étude (Alcock, Wade, Anslow et Passingham, 2000) faite sur des participants dysphasiques a démontré que les participants dont PHG était touché avaient beaucoup de difficulté à produire et à percevoir auditivement le rythme. Dans une autre recherche (Gates & Bradshaw, 1977), des sujets entendent une mélodie à l’aide d’écouteurs et ils doivent y détecter des changements soit au niveau de la tonalité, du rythme ou de l’harmonie. Les résultats de l’expérience 3 soutiennent que pour détecter un changement de rythme auditif (séquence de cinq notes), l’oreille gauche est plus rapide (temps de réaction plus court) et que l’oreille droite est plus précise (nombre moyen de changements de rythme manqués moins élevé). À l’expérience 5 où des changements au niveau de la tonalité et du rythme sont apportés, les auteurs concluent que PHD est plus rapide pour les détections plus faciles qui peuvent être faites rapidement tandis que PHG est plus précis pour les changements plus difficiles qui sont moins facilement reconnaissables. Appliquées à la musique, ces données permettent de conclure d’une part que PHD analyse le son de manière globale ainsi il détecte plus facilement les changements qui sont plus évidents (faciles) et d’autre part que la supériorité de PHG pour les détections plus difficiles vient du fait qu’il interprète les
changements musicaux par analyse (de manière analytique). Malgré le grand nombre d’études reliant PHG au traitement rythmique, il existe certaines recherches qui associent ce traitement à PHD (Ben-Dov & Carmon, 1984). Par exemple, Milner (1962) présente des séquences
rythmiques auditives comportant cinq à sept éléments à des patients avant et après une
lobectomie temporale. Elle démontre qu’il y a une baisse de performance après une chirurgie à PHD.
Par ailleurs, Gordon (1978) démontre une supériorité de l’oreille droite pour des participants qui avaient à distinguer des mélodies selon le rythme par la méthode d’écoute dichotique. Selon l’auteur de cette recherche, le mode de réponse comme la nature d’un passage musical, d’une mélodie ou d’une séquence tonale sont tous des éléments importants pour établir la préférence d’un hémisphère pour ce type de processus. Ainsi, les processus dépendants du temps (séquentiel ou temporel), dont le rythme est un exemple, seraient mieux accomplis par les fonctions de l’HG tandis que les processus indépendants du temps, dont les accords musicaux sont un exemple, seraient mieux accomplis par les fonctions de l’HD. Les résultats supportent une dichotomie gauche-droite basée sur la notion du temps et non une dichotomie fondée sur la notion
verbale/non-verbale. En effet, parmi tous les tests portant sur les composantes musicales, le test de signal rythmique utilisé dans cette recherche s’insère davantage dans une liste de tests de l’HG désignés analytiques, étape par étape, séquentiels ou temporels (= dépendant du temps) plutôt que ceux associés à l’HD désignés synthétiques, entiers, en unité, complétant l’ensemble ou Gestalt (- indépendant du temps). Bref, selon Gordon (1978), la recatégorisation de ces termes dans une dichotomie basée sur le temps apporte une base solide sur laquelle les propriétés des hémisphères peuvent être définies.
Les différences fonctionnelles des hémisphères concernant la dichotomie analytique/holistique peuvent s’appliquer dans le traitement temporel de séquences rythmiques visuelles. Ben-Dov et
Carmon (1984) demandaient à 24 participants droitiers de déterminer si deux séquences rythmiques visuelles étaient identiques ou différentes. Les intervalles de temps utilisés étaient courts (200 ms) ou longs (400 ms) tandis que la longueur des séquences rythmiques pouvait inclure 1, 2, 3 ou 4 intervalles de temps entrecoupés de flashes lumineux de 5ms. La première séquence rythmique était envoyée dans les deux champs visuels tandis que la deuxième séquence était dirigée dans un seul champ visuel (droit ou gauche). Ainsi, lorsque tous les intervalles de temps de la deuxième séquence rythmique étaient identiques à ceux de la première séquence le participant répondait «identique» et lorsqu’un seul intervalle de temps était différent, celui-ci devait répondre «différent». Les résultats ont démontré que les temps de réaction moyens augmentaient significativement de 1 intervalle de temps à 4 intervalles de temps. De plus, il s’effectue un changement graduel entre la supériorité des hémisphères passant de l’HG pour 1 intervalle de temps à l’HD pour 4 intervalles de temps. Ainsi, en augmentant le nombre
d’éléments la dominance cérébrale passe de gauche à droite. En effet, le nombre d’éléments contenus dans la séquence influencerait le mode de traitement utilisé par le cerveau. Lorsque le nombre d’éléments est petit, les rythmes sont comparés séquentiellement, un par un, bref, selon le mode de traitement caractérisant l’HG est utilisé. Lorsque le nombre d’éléments augmente, ce mode de traitement devient inefficace alors PHD prend la place en analysant holistiquement la séquence de rythme comme étant un patron temporel global. Ainsi, le rythme peut être représenté comme une séquence d’éléments ou comme un patron temporel (Gestalt) (Ben-Dov & Carmon, 1984).
La présente étude
L’objectif de la présente étude est d’examiner s’il existe des différences de performance entre l’HG et PHD en ce qui concerne la discrimination temporelle d’intervalles placés dans une séquence rythmique de stimuli visuels.
Il y a trois variables indépendantes (VI) qui sont à l’étude dans cette recherche : 1- La durée des intervalles de temps standards (900, 1125 et 1406 ms)
2- U endroit de présentation du stimulus dans le champ visuel (droite ou gauche) 3- Le nombre d’intervalles de temps standards présenté (deux ou quatre).
Cette étude comporte trois variables dépendantes (VD) :
1- Le seuil différentiel démontrant la sensibilité et la capacité du participant à discriminer les intervalles temporels.
2- Le point d’égalité subjective ou sa valeur dérivée, Y erreur constante (point d’égalité
subjective - standard), déterminant la durée perçue du sujet et sa proportion de bonnes réponses. 3- La fraction de Weber (seuil différentiel / point d’égalité subjective) qui est aussi une mesure de la sensibilité de discrimination du participant.
Hypothèses
Le temps et le rythme sont des composantes majeures de plusieurs processus fondamentaux tels que le langage (lié à l’HG) et la musique (liée à l’HD). Ainsi, le traitement de ces composantes pourraient avoir lieu autant dans un hémisphère que dans l’autre. Toutefois, la majorité des
études obtiennent des résultats qui établissent des liens entre PHG et les processus temporels et rythmiques.
Hypothèse 1
La performance relative des deux hémisphères cérébraux, telle qu’évaluée avec le seuil différentiel, dépendra du nombre d’intervalles standards présentés.
Hypothèse 2
Il y aura une augmentation de la fraction de Weber avec les intervalles de temps les plus longs (i.e., avec l’intervalle standard = 1406 ms).
MÉTHODE
Participants
Trois étudiants et neuf étudiantes droitiers âgés de 20 à 32 ans (moyenne = 24.5 ans) de l’Université Laval participent à l’expérience. Celle-ci consiste en douze sessions d’une durée approximative allant de 23 à 43 minutes chacune. Les participants reçoivent 5,00$ par session pour un grand total de 60,00$. La dominance manuelle est déterminée par le test de manualité de Bryden (1977) comportant cinq items (Annexe E).
Matériel
Chaque participant est assis sur une chaise dans une chambre expérimentale dont la lumière est tamisée. Un ordinateur Zénith contrôle les intervalles de temps à estimer. Chaque intervalle est séparé successivement par un signal visuel de 10 ms. Une diode circulaire rouge (LEDs :Radio- Shack # 276-088) constitue ce signal visuel. Cette diode est placée à 25° à gauche ou à droite d’un point de fixation. De cette manière, le stimulus n’est pas projeté sur la tache aveugle de la rétine de l’œil. Le point de fixation est positionné à un mètre en face du participant. Le
participant appuiera sur l’un des deux boutons du boîtier de réponse pour donner sa réponse. Le bouton de gauche sera utilisé par le participant pour répondre court et le bouton de droite pour répondre long.
Procédure
Chaque essai consiste en une présentation dans le CVG ou dans le CVD de deux ou quatre
intervalles de temps séparés respectivement par trois ou cinq signaux visuels. Les participants ont à indiquer, en appuyant sur le bouton approprié, si le dernier intervalle (comparaison) de la séquence est plus court ou plus long que les intervalles précédants qui sont les standards. Il y a trois intervalles de temps standards à l’étude : 900, 1125 et 1406 ms. Bref, les participants complètent 12 conditions expérimentales chacun : 2 endroits de présentation X 3 durées de base X 2 nombre de standards.
Pour chaque intervalle standard, il y a 6 intervalles de comparaisons. Ceux-ci suivent une règle consistant à multiplier par 1,25 une durée à l’autre. À 900 ms, les intervalles de comparaison vont
de 650 à 1150 ms par tranche de 100 ms. Avec 1125 ms, la valeur des intervalles de comparaison vont par tranche de 125 ms, de 813 à 1438 ms ; à 1406 ms ils vont par tranche de 156 ms, de
1016 à 1797 ms. La durée entre chaque essai est fixée à 2 secondes. Comme pour ten Hoopen, Hartsuiker, Sasaki, Nakajima, Tanaka et Tsumura (1995), il n’y a pas de feedback. Chaque session est divisée en quatre blocs de 60 essais. Dans chaque bloc, il y a 10 répétitions de chacun des 6 intervalles de comparaison. Entre chaque bloc, il y a une pause de 20 secondes.
Un contre-balancement des participants dans les différentes conditions a été effectué. Trois groupes de quatre participants sont formés selon l’ordre de présentation des intervalles de temps standards (900-1125-1406 ms ; 1406-900-1125 ms ; 1125-1406-900 ms) pour chacune des sessions. À l’intérieur de ces trois groupes, chaque participant est assigné à chacune des quatre combinaisons résultant de la combinaison des deux conditions du champ visuel et des deux conditions du nombre de standards.
RÉSULTATS
Données préliminaires
Une fonction psychométrique a été tracée pour chacun des douze participants et pour chacune des douze conditions à Vétude. Sur l’axe des x, on retrouve la valeur des stimuli de comparaison et sur Taxe des y, on retrouve les probabilités de répondre que le stimulus de comparaison est plus grand que le stimulus standard. La fonction psychométrique obtenue est ajustée à une distribution normale cumulative. La valeur de cet ajustement est définie à l’aide de l’indice R2. Les valeurs de R2 se situent, à l’exception d’un cas (.8819), entre .9356 et .9999 (Tableau 1). Ainsi, les valeurs élevées des R2 dans la présente étude permettent d’avoir un niveau de confiance plus élevée envers les données recueillies.
Insérer Tableau 1
Comparaison des moyennes
La moyenne et T erreur-type des VD (le point d’égalité subjective, le seuil différentiel et la fraction de Weber définie comme le seuil différentiel divisé par le point d’égalité subjective) en fonction des VI à l’étude (le champ visuel, le nombre d’intervalles de temps standards et la longueur du standard) sont présentées dans les tableaux 2 à 7.
Insérer Tableaux 2 à 7
Trois ANOVAs selon un plan factoriel en blocs aléatoires 2x2x3 ont été effectués afin de déterminer s’il existait des différences significatives entre les trois moyennes et ce, pour les trois VD à l’étude. Afin de permettre une comparaison des trois niveaux utilisés pour la variable standard, Terreur constante (point d’égalité subjective - standard) est utilisée plutôt que le point d’égalité subjective.
Hypothèse 1
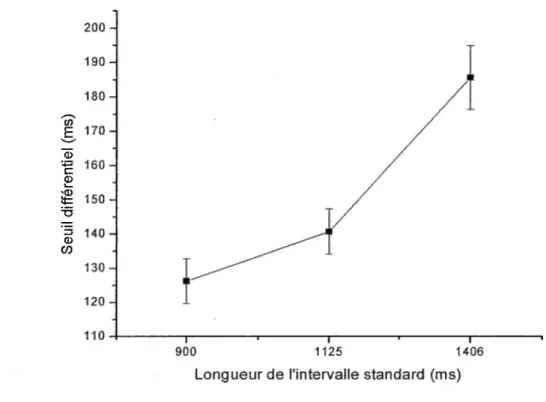
L’ANOVA pour le seuil différentiel (Tableau 8) montre un effet principal significatif de longueur des intervalles de temps. Il consiste en !’augmentation du seuil différentiel (de 1
sensibilité) à mesure que la longueur des intervalles de temps standards augmente [F(2,l 1) = 28.84, p < .001]. L’effet principal du champ visuel ne s’est pas révélé significatif [F( 1,11) = .32,
p = .575] ainsi que celui du nombre de standards utilisé [F(l,l 1) = 3J9,p = .054]. L’analyse
statistique montre un effet d’interaction significatif du champ visuel utilisé par le nombre
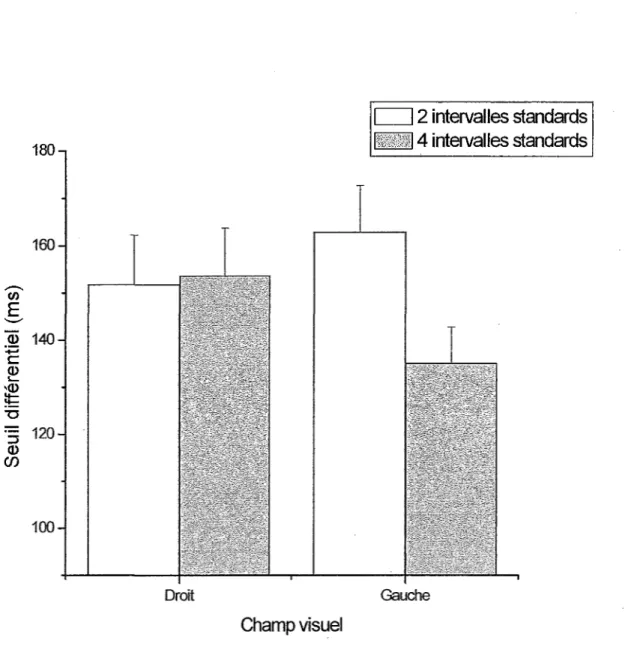
d’intervalles de temps standards [F(l,l 1) = 4.96,p < .05]. Un test d’effets simples principaux est effectué pour décortiquer !’interaction entre le champ visuel et le nombre d’intervalles standards. Celui-ci indique qu’il n’existe pas de différences significatives entre les hémisphères cérébraux lors de !’utilisation de deux standards [F(l,l 1) = 1.33, p = .254] (Tableau 9). Ainsi, la sensibilité des participants n’est pas différente entre les deux hémisphères quand deux intervalles de temps sont utilisés. Par contre, il existe des différences significatives sur le plan du champ visuel pour 4 intervalles de temps standards [F(l,l 1) = 4.03, p < .05] (Tableau 10). Ainsi, lorsque 4 intervalles de temps sont présentés, l’HD a une plus grande sensibilité pour discriminer ces intervalles de temps que l’HG.
Insérer Tableaux 8, 9 et 10
Les tests montrent également qu’il existe des différences significatives entre le nombre de standards utilisé (2 ou 4) dans le CVG [F(l,l 1) = 12.07,p < .01] mais pas dans le CVD [F(l,l 1) = .04,p = .851]. Ainsi, lorsque 4 standards plutôt que 2 sont présentés dans le CVG, la
discrimination temporelle est supérieure.
Insérer Tableaux 11 et 12
Hypothèse 2
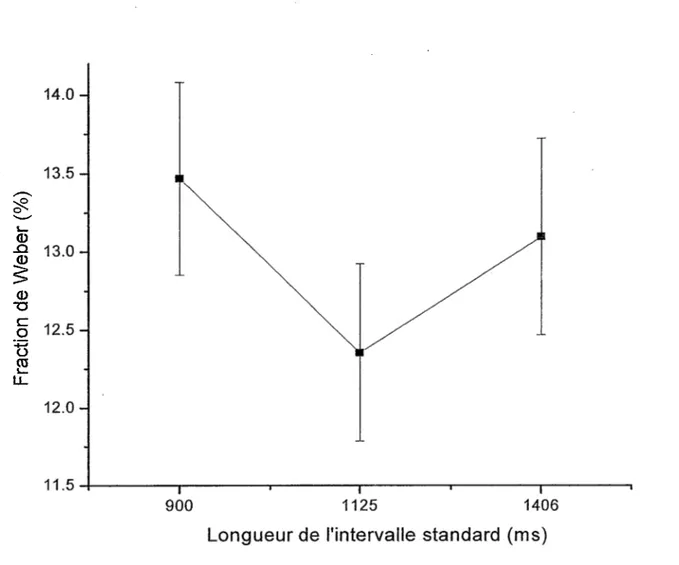
En ce qui concerne la fraction de Weber, aucun effet ne s’est révélé significatif concernant la longueur des intervalles de temps [F(2,ll) = 1.43, p = .243] (Tableau 13). Toutefois, les résultats montrent une certaine tendance : de 900 ms à 1125 ms la fraction de Weber diminue (ce qui est attendu) et de 1125 ms à 1406 ms celle-ci augmente.
Erreur constante
Même si aucune hypothèse spécifique n’avait été formulée concernant l’erreur constante, l’analyse des résultats pour cette variable a été faite et VANOVA n’a révélé aucune différence significative (Tableau 14).
DISCUSSION
Le but principal de cette recherche est de déterminer s’il existe des différences de performance au niveau des hémisphères gauche et droit selon que les intervalles de temps standards sont
présentés en série de 2 ou de 4 et qu’ils sont de 900 ms, 1125 ms ou 1406 ms. Un autre objectif de cette étude est de vérifier si la loi de Weber démontre des irrégularités pour les valeurs de 900 ms, 1125 ms et 1406 ms et ce, pour les hémisphères gauche et droit. Principalement, les résultats de cette expérience ont permis de découvrir un effet de latéralité au niveau du seuil différentiel. De plus, les données démontrent que la fraction de Weber tend à ne pas demeurer constante pour la valeur supérieure à 1125 ms.
Seuil différentiel
En ce qui concerne le seuil différentiel, des différences significatives ont été trouvées au niveau de la durée de l’intervalle de temps standard. Conformément à ce qui est établi dans le domaine de la psychophysique et plus précisément par rapport à la fraction de Weber, le seuil différentiel augmente à mesure que la durée de l’intervalle de temps standard augmente. C’est ce que les résultats ont montré lors de l’analyse.
Des différences significatives au niveau du seuil différentiel sont présentes pour le nombre d’intervalles de temps standards par rapport au champ visuel. Une différence significative de moyennes a été démontrée entre la condition de 2 standards et la condition de 4 standards. Cette différence se retrouve exclusivement au niveau de l’HD. En effet, l’HD est significativement meilleur (seuil différentiel plus bas) dans les conditions comportant 4 intervalles de temps standards que dans les conditions n’en comportant que 2. Pour l’HG, il en est tout autrement puisqu’aucune différence significative n’a pu être décelée. De plus, en utilisant 4 standards l’HD obtient une performance significativement meilleure que l’HG.
Distinction analvtique/holistique
Il est possible d’expliquer certaines données de la présente recherche, en ce qui concerne le changement de supériorité hémisphérique selon le nombre de standards utilisé, par la dichotomie analytique/holistique. Les résultats au niveau du seuil différentiel montrent qu’à 2 standards la performance de l’HG est supérieure, mais de manière non significative, à celle de PHD tandis qu’à 4 standards, PHD permet d’obtenir une performance significativement meilleure que l’HG. Ainsi, l’HG utiliserait un mode analytique (un par un) pour traiter !’information et serait plus rapide que PHD lorsque le nombre d’éléments dans la séquence serait petit (2). Cependant, lorsque le nombre d’éléments augmente (4), c'est la capacité d’analyser !’information globalement de PHD qui prendrait le relais (Ben Dov & Carmon, 1984).
Les résultats de la présente étude sont en grande partie conformes à ceux obtenus par Ben-Dov et Carmon (1984) illustrant aussi la dichotomie analytique/holistique et par conséquent, ils
soutiennent l’une de nos hypothèses de base concernant les différences hémisphériques. Cette hypothèse supporte l’idée que la performance est supérieure au niveau de l’HG lorsque le nombre d’intervalles de temps standards présenté est petit (2), et lorsque le nombre d’intervalles de temps standards présenté est grand (4), c’est au niveau de PHD que la performance est supérieure. Les données de Ben-Dov et Carmon (1984) montraient qu’il y avait un passage de la supériorité de PHG à PHD lorsque le nombre d’intervalles de temps augmentait. Toutefois, l’expérience de Ben-Dov et Carmon (1984) n’était pas en tout point identique à la présente expérience. En effet, les sujets devaient déterminer si deux séquences rythmiques visuelles étaient identiques ou différentes, les intervalles de temps utilisés étaient de 200 ms ou 400 ms et la longueur des séquences rythmiques étaient de 1, 2, 3 ou 4 intervalles de temps. De plus, la première séquence rythmique était envoyée dans les deux champs visuels tandis que la deuxième séquence était dirigée dans un seul champ visuel (droit ou gauche).
Une expérience de Milner (1962) dans laquelle des séquences rythmique auditives comportant cinq à sept éléments étaient présentées à des patients avant et après une lobectomie temporale démontrait qu’il y a une certaine baisse de performance (le nombre d’erreurs augmente) après une chirurgie à PHD. Ainsi, lorsque le nombre d’éléments dans une série est élevé il semble que c’est la performance de PHD qui est affectée.
Le fait que FHD soit significativement meilleur dans les conditions comprenant 4 intervalles de temps que dans celles en comprenant 2 va dans le même sens que les résultats de l’expérience A de Grondin (2001a). Ces résultats démontrent une tendance à l’amélioration de la discrimination avec !’augmentation de la présentation du standard à 600 et 900 ms pour la condition continue (condition comparable à celle utilisée dans ce mémoire). Dans la présente expérience, il est aussi possible d’observer une baisse du seuil différentiel avec !’augmentation du nombre de standards présentés : à 900, 1125 et 1406 ms pour FHD et d’une manière non significative à 900 ms pour FHG.
Modèle temporel
Il est aussi possible d’expliquer les données de la présente recherche pour les différences
interhémisphériques du seuil différentiel en rapport avec le nombre de standards en appliquant le modèle de Thomas et Weaver (1975), tout en conservant !’interprétation de Polzella et al. (1977) en ce qui concerne la différence fonctionnelle des hémisphères pour ce modèle. Cette
interprétation établissait que FHD avait un processeur pour !’information visuelle tandis que FHG fonctionnait à l’aide d’une horloge. Sans être significatifs, certains résultats de ce mémoire démontrent que la sensibilité est supérieure à FHG comparativement à FHD lorsque le nombre de stimuli est plus réduit (2 standards). Toutefois, cette situation s’inverse lorsque le nombre de stimuli augmente (4 standards). L’HD a donc une meilleure sensibilité pour un plus grand nombre de stimuli. Ceci est en accord avec les résultats rapportés par Polzella et al. (1977) qui montraient que FHG avait une performance supérieure lorsque le nombre de stimuli était plus petit puisque !’attention était dirigée vers l’horloge avec laquelle il fonctionnait. Quant à FHD, il montrait une meilleure performance lorsque l’information visuelle augmentait (avec plus de stimuli visuels) car cet hémisphère fonctionnerait avec un processeur d’information visuelle.
Loi de Weber
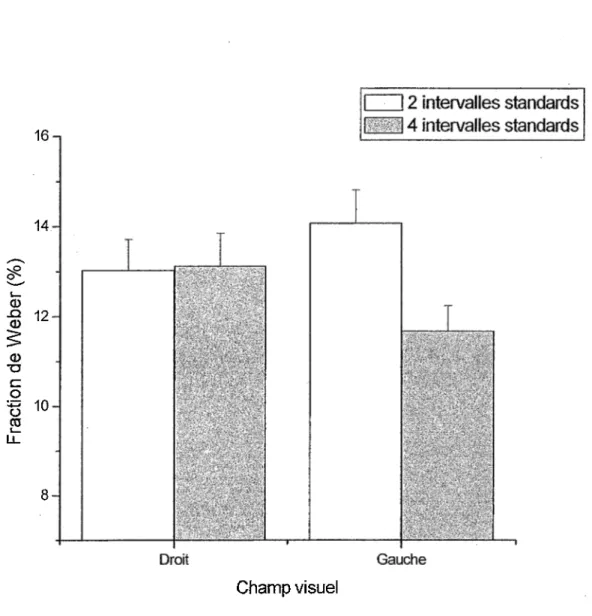
Pour la fraction de Weber, les résultats ont démontré une interaction significative entre le nombre d’intervalles de temps standards et l’endroit où était projeté les stimuli dans le champ visuel (CVD ou CVG). Ces résultats sont similaires à ceux obtenus au niveau du seuil différentiel. Pour ce qui est de l’effet du nombre de standards, il s’avère qu’à 2 standards la fraction de Weber est significativement plus élevée qu’à 4 standards. Ainsi, le nombre d’éléments contenus dans la
séquence semble influencer l’allure de la courbe de la loi de Weber. Selon Grondin (2001b), la fraction de Weber demeure constante pour tous les intervalles de temps à l’exception de ceux très courts où la fraction de Weber augmente. Ce principe n’est toutefois pas démontré dans cette étude.
Insérer Figure 3
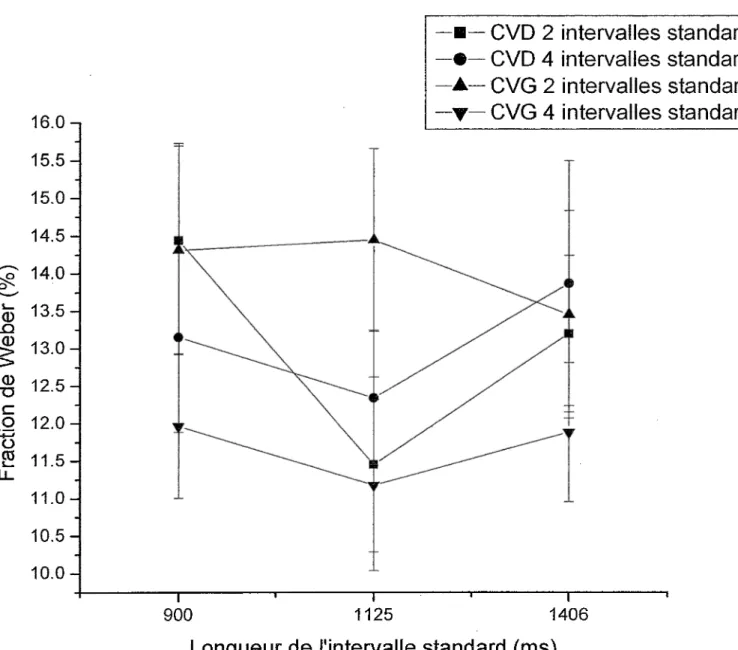
Malgré l’absence de différences significatives au niveau de !’interaction entre le champ visuel et la longueur de l’intervalle standard (sa durée), les résultats démontrent la tendance de la fraction de Weber à diminuer de 900 ms à 1125 ms pour ensuite remonter à 1406 ms (Figure 5). Plus précisément, la valeur moyenne de la fraction de Weber pour le CVD baisse de 900 ms à 1125 ms pour ensuite remonter à 1406 ms, ce qui n’est pas le cas pour le CVG. En considérant aussi le nombre de standards utilisé, il est possible d’observer que cette tendance est présente pour toutes les combinaisons entre le champ visuel et le nombre de standards excepté lorsque 2 standards sont projetés dans le CVG (Figure 1). En général, ces résultats vont dans le même sens que ceux obtenus par Grondin (2001a) soutenant que la fraction de Weber serait plus élevée pour 1200 ms plutôt que pour 900 ms. D’autres recherches utilisant les processus rythmiques démontrent des résultats semblables indiquant des valeurs où la fraction de Weber est plus basse. Par exemple, Drake et Botte (1993) ont trouvé que pour des valeurs allant de 100 à 1500 ms, c’est entre 300 et 800 que la fraction de Weber est la plus basse. Les résultats de Friberg et Sundberg (1995), quant à eux, montrent que c’est à 500 ms que la fraction de Weber subit une diminution.
Insérer Figures 4, 5 et 6
Erreur constante
Aucun effet ne s’est révélé significatif en ce qui concerne le champ visuel et la durée perçue (telle qu’exprimée par l’erreur constante). Les résultats ne sont pas conformes à ceux obtenus par Polzella et al. (1977) qui démontraient que la durée moyenne est estimée comme étant plus longue lorsque les stimuli sont présentés dans le CVD-HG comparativement à lorsqu’ils sont présentés dans le CVG-HD. Dans la présente expérience, les résultats ne démontrent pas que les intervalles de temps ont été perçus comme étant plus longs lorsqu’ils étaient présentés dans le
CVD-HG. De cette manière, la durée perçue des intervalles de temps ne serait pas
significativement différente d’un hémisphère à l’autre dans cette étude. Ceci pourrait s’expliquer par le fait que la méthode utilisée dans l’expérience de Polzella et al. (1977) était différente. En effet, la durée à estimer concernait des ensembles de points qui flashaient contrairement à la présente étude qui utilise l’intervalle de temps entre des séquences de flash lumineux pour estimer le temps. Alors, il pourrait avoir une différence pour l’estimation du temps d’apparition d’un stimulus visuel plutôt que pour l’apparition d’un intervalle de temps entre deux stimuli visuels. Il est aussi possible que les deux hémisphères cérébraux possèdent une capacité à estimer la durée qui ne diffère pas beaucoup. Les résultats de cette même recherche (Polzella et al., 1977) montraient que la durée était sous-estimée sans égard au champ visuel. Ces données ne sont pas tout à fait en accord avec ceux de la présente recherche étant donné qu’il y a plutôt une tendance non significative à surestimer la durée des intervalles de temps de comparaison par rapport aux intervalles de temps standard pour toutes les conditions qui sont à l’étude dans la présente expérience.
Recom mandations
Les données de cette expérience viennent valider les résultats de recherches antérieures et
apportent un éclairage nouveau sur le sujet de la latéralité. En effet, la structure de la séquence de stimuli utilisée amène des changements significatifs au niveau de la supériorité hémisphérique. De cette manière, il devient possible de tester les différences hémisphériques de multiples manières en changeant la forme et l’apparition de la séquence de stimuli. Par exemple, il serait possible d’utiliser des séquences visuelles plus longues soit des séquences comprenant 6 ou même 8 stimuli visuels afin de vérifier si la tendance à la supériorité de l’HD demeure. De plus, il serait intéressant d’utiliser d’autres gammes de durées, soit plus longues ou soit plus courtes, ce qui permettrait de voir si la loi de Weber subit des violations pour des valeurs différentes.
Finalement, la même expérience pourrait être effectuée avec des populations cliniques telles que des personnes ayant subi un traumatisme crânio-cérébral (TCC) afin de déterminer si les
différences hémisphériques au niveau de la perception temporelle se retrouvent aussi au niveau de ce type de population. Il serait intéressant de comparer celles qui ont un dommage cérébral à droite à celles qui ont un dommage à gauche et les comparer à une population normale pour voir
si leur performance est inférieure, étant donné les résultats de la présente recherche favorisant PHD dans le traitement de séquences rythmiques visuelles.
CONCLUSION
Cette recherche permet d’apporter un appui aux théories de la latéralisation. En effet, les résultats permettent de supposer qu’il existe des différences entre les hémisphères cérébraux en ce qui concerne les processus temporels rythmiques en modalité visuelle. Toutefois, celles-ci se situent uniquement au niveau de la sensibilité des participants (seuil différentiel et fraction de Weber). La présente recherche fait avancer les connaissances en neuropsychologie et permet de faire des liens entre le temps, les hémisphères cérébraux, le langage (hé à l'HG) et le rythme (lié à !,HD). Elle permet aussi !’application de ces connaissances sur le plan clinique. En effet, en s’appuyant sur les lésions cérébrales associées à certaines pathologies (aphasie, traumatisme crânio-cérébral, etc.) et sur la dichotomie analytique-holistique, il est possible de déterminer la manière optimale de transmettre !’information temporelle à chacune des populations cliniques. Par exemple, pour les personnes lésées à droite, la présentation de !’information devrait se faire avec un plus petit nombre d’éléments. Afin de confirmer ce principe, des recherches ayant un plus grand
échantillon de sujets qui proviendraient de populations cliniques devront être réalisées. À la lumière de ces nouvelles connaissances, il sera possible de développer des outils pour aider les gens souffrant de troubles neurologiques.
RÉFÉRENCES
Alcock, K. J., Wade, D., Anslow, P., & Passingham, R. E. (2000). Pitch and timing abilities in adult left-hemisphere-dysphasic and right-hemisphere-damaged subjects. Brain and
Language, 75, 47-65.
Baeck, E. (2002). The neural network of music. European Journal of Neurology, 9, 449-456.
Ben-DoV, G., & Carmon, A. (1984). Rhythm length and hemispheric asymmetry. Brain &
Cognition, 3, 35-41.
Bradshaw, J. L., & Nettleton, N. C. (1981). The nature of hemispheric specialisation in man.
Behavioral and Brain Sciences, 4, 51-91.
Braun, C. M. J. (2000). Neuropsychologie du développement. Paris: Flammarion Médecine- Sciences.
Bryden, M.P. (1977). Measuring handedness with questionnaires. Neuropsychologia, 15, 617-624.
Bryden, M. P. (1982). Laterality: functional asymmetry in the intact brain. New York: Academic Press.
Carmon, A. (1979). Spatial and temporal factors in visual perception of patients with unilateral cerebral lesions. In M. Kinsboume (Ed.), Asymmetrical function of the brain. Cambridge : Cambridge University Press.
Carmon, A., & Nachshon, I. (1971). Effect of unilateral brain damage on perception of temporal order. Cortex, 7, 410-418.
Davis, A. E., & Wada, J. A. (1977). Hemispheric asymmetries of visual and auditory information processing. Neuropsychologia, 15, 799-806.
Davis, R., & Schmit, V. (1971). Timing the transfert of information between hemispheres in man.
Acta Psychologica, 35, 335-346.
Drake, C., & Botte, M.-C. (1993). Tempo sensitivity in auditory sequences: Evidence for a multiple-look model. Percepetion & Psychophysics, 54,277-286.
Efron, R. (1990). The decline and fall of hemispheric specialization. Hillsdale, NJ: Lawrence Erlbaum Associates.
Friberg, A., & Sundberg, J. (1995). Time discrimination in a monotonie, isochronic sequence.
Journal of the Acoustical Society ofAmerica, 98, 2524-2531.
Gates, A. & Bradshaw, J. L. (1977). Music perception and cerebral asymmetries. Cortex, 13, 390-401.
Gordon, H. W. (1978). Left hemisphere dominance for rhythmic elements in dichotically- presented melodies. Cortex, 14, 58-70.
Grondin, S. (1998). Judgments of the duration of visually marked empty time intervals: Linking perceived duration and sensivity. Perception & Psychophysics, 60, 319-330.
Grondin, S. (2001a). Discriminating time intervals presented in sequences marked by visual signals. Perception & Psychophysics, 63, 1214-1228.
Grondin, S. (2001b). From physical time to the first and second moments of psychological time.
Iaccino, J. F. (1993). Left brain - right brain differences: Inquiries, evidence, and new
approaches. Hillsdale, NJ: Lawrence Erlbaum Associates.
Ivry, R. B., & Robertson, L. C. (1998). The two sides of perception. Massachaussets: The MIT Press.
Kallman, H. J., & Corballis, M. C. (1975). Ear asymmetry in reaction time to musical sounds.
Perception & Psychophysics, 17, 368-370.
Kimura, D. (1961). Cerebral dominance and the perception of verbal stimuli. Canadian Journal
of Psychology, 15, 166-171.
Lezak, M. (1995). Neuropsychological Assessment, 3rd ed. New York: Oxford University Press.
Marieb, E. N. (1993). Anatomie et physiologie humaines. St-Laurent : ERPI.
Mills, L., & Rollman, G. B. (1980). Hemispheric asymmetry for auditory perception of temporal order. Neuropsychologia, 18, 41-47.
Milner, B. (1962). Laterality effects in audition. In V. R Mountcastle (Ed.) Interhemispheric
relations and cerebral dominance, (pp. 177-195) Baltimore: John Hopkins University
Press.
Nicholls, M. E. R (1994). The non-contribution of attentional biases to visual field asymmetries for temporal discrimination. Neuropsychologia, 32, 209-220.
Nicholls, M. E. R. (1996). Temporal processing asymmetries between the cerebral hemispheres: Evidence and implications. Laterality, 1, 97-137.
Nicholls, M. E. R, & Cooper, C. J. (1991). Hemispheric differences in the rates of information processing for simple non-verbal stimuli. Neuropsychologia, 29, 677-684.
Polzella, D. 1, DaPolito, F., & Hinsman, M. C. (1977). Cerebral asymmetry in time perception.
Perception & Psychophysics, 21, 187-192.
Robinson, G. Μ. & Solomon, D. J. (1974). Rhythm is process by the speech hemisphere. Journal
of Experimental Psychology, 102, 508-511.
Semmes, J. (1968). Hemispheric specialisation: A possible clue to mechanism.
Neuropsychologia, 6, 11-26.
Springer, S. P. & Deutsch, G. (1998). Left brain, right brain: Perspectives from cognitive
neurosciences (5e ed.). New York: W. H. Freeman.
ten Hoopen, G., Hartsuiker, R., Sasaki, T., Nakajima, Y., Tanaka, M., & Tsumura, T. (1995). Auditory isochrony: Time shrinking and temporal patterns. Perception, 24, 577-593.
Thomas, E. A. C., & Weaver, W. B. (1975). Cognitive processing intime perception. Perception
& Psychophysics, 17, 363-367.
Vroon, P. A., Timmers, H., & Tempelaars, S. (1977). On the hemispheric representation of time. In S. Domic (Ed.), Attention and performance VI: Proceedings of the Sixth Internationa;
Symposium on Attention and Performance (pp. 231-245). Hillsdale, NJ: Erlbaum.
White, M. J. (1969). Laterality differences in perception: A review. Psychological Bulletin, 72,
387-405.
Wilson, S. J., Pressing, J. L., & Wales, R. J. (2002). Modelling rhythmic fonction in a musician post-stroke. Neuropsychologia, 40, 1494-1505.