Pépite | Apport de la spectroscopie Raman à l’étude des mécanismes de stabilisation des protéines par les disaccharides et cyclodextrines
149
0
0
Texte intégral
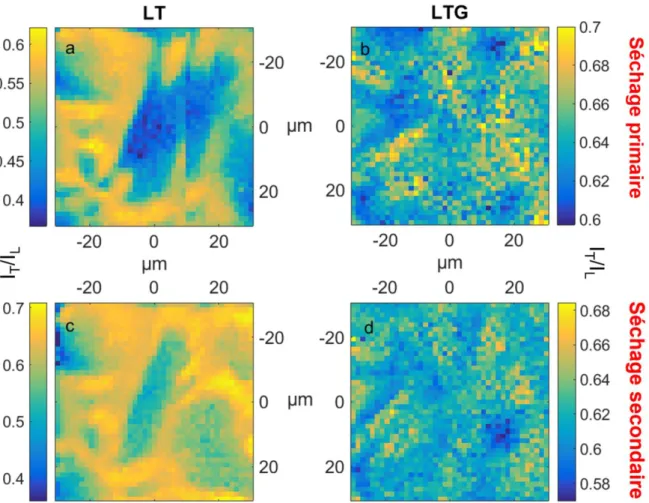
Figure
Documents relatifs