The roles of the NOOT-BOP-COCH-LIKE genes in the symbiotic organ identity and in plant development
314
0
0
Texte intégral
Figure
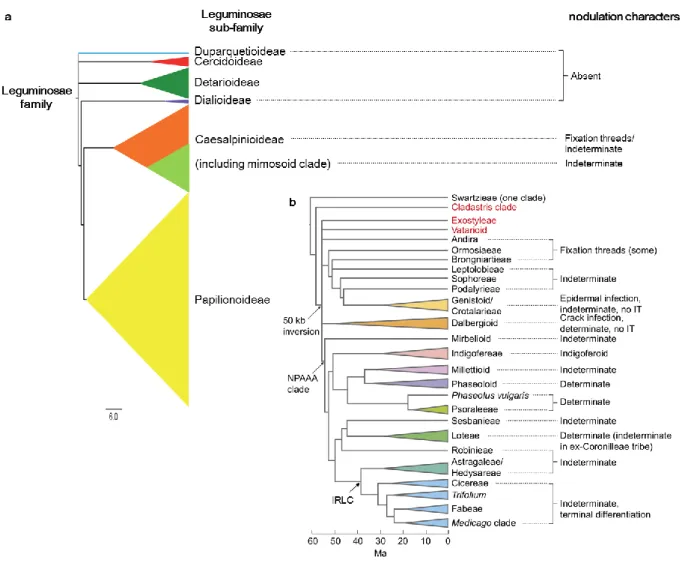
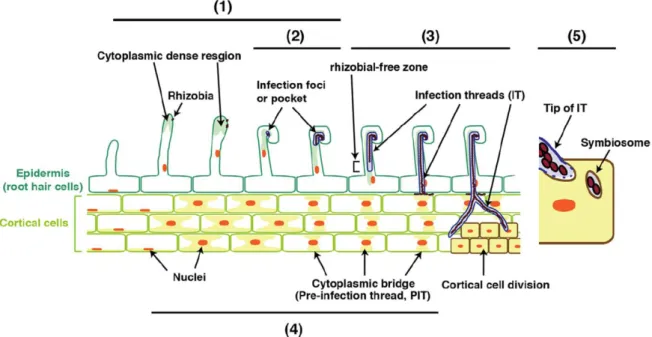
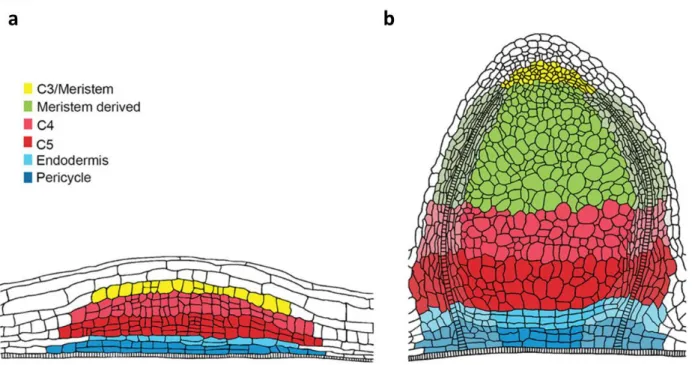

+7
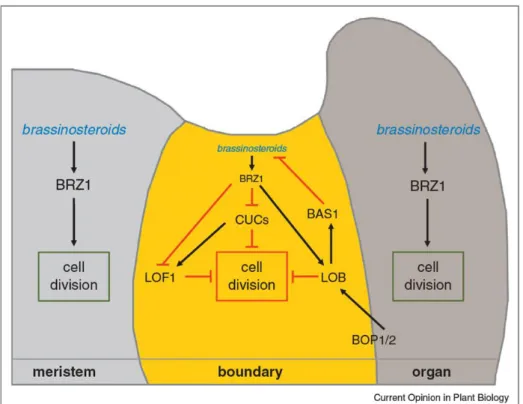
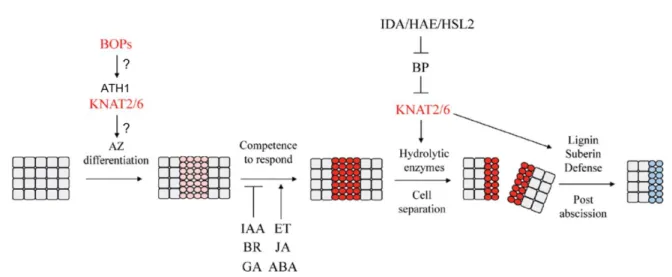

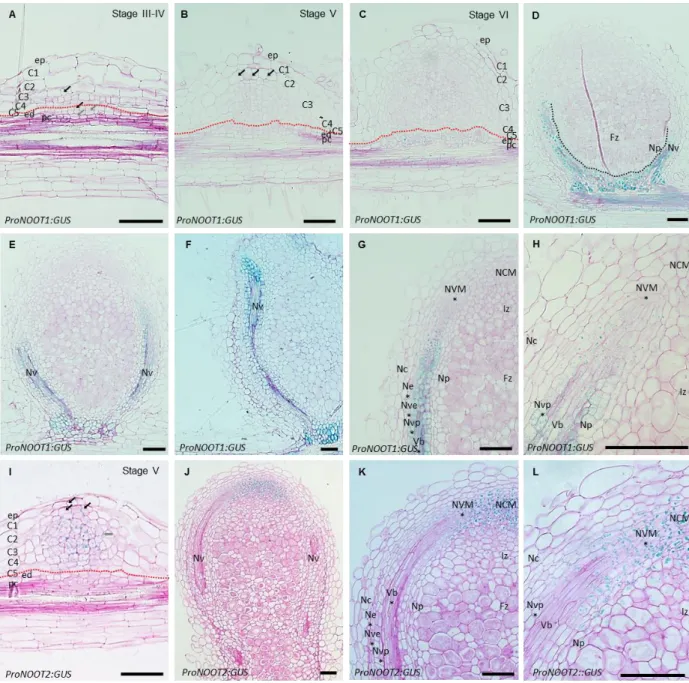
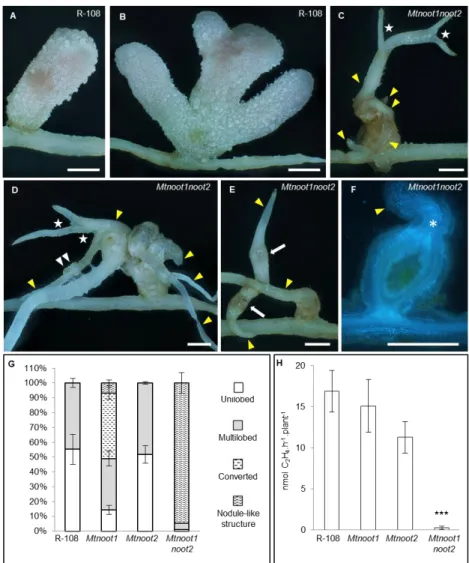
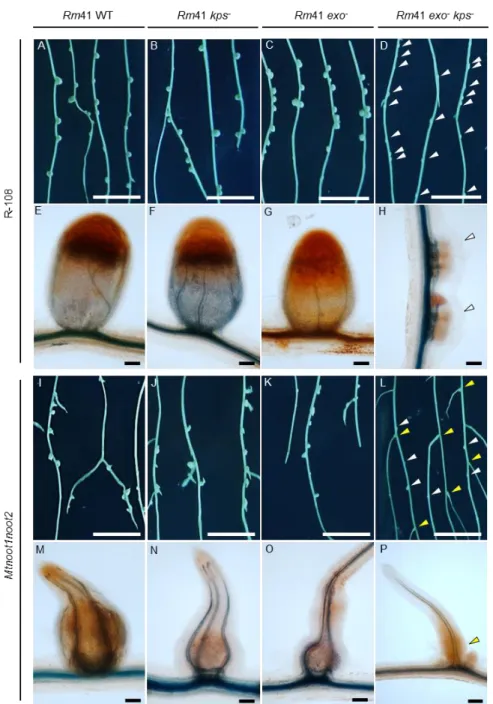
Documents relatifs