Inhibition cellulaire de la proprotéine convertase 1 et activité des proprotéines convertases dans le réticulum endoplasmique
Texte intégral
Figure
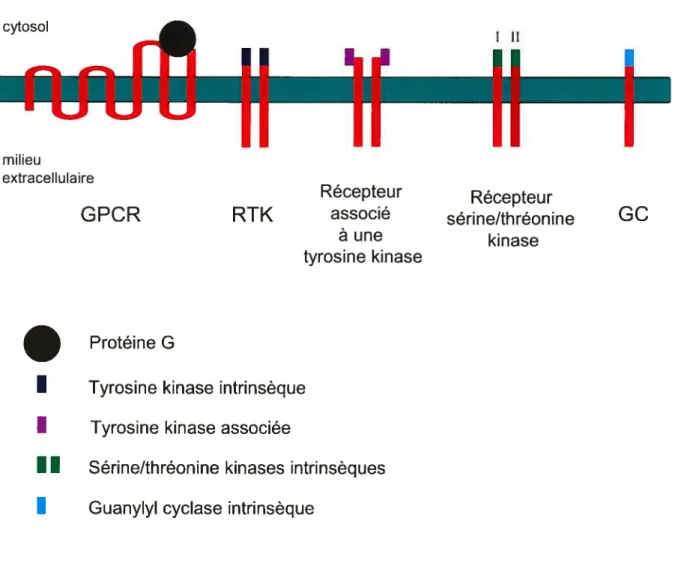

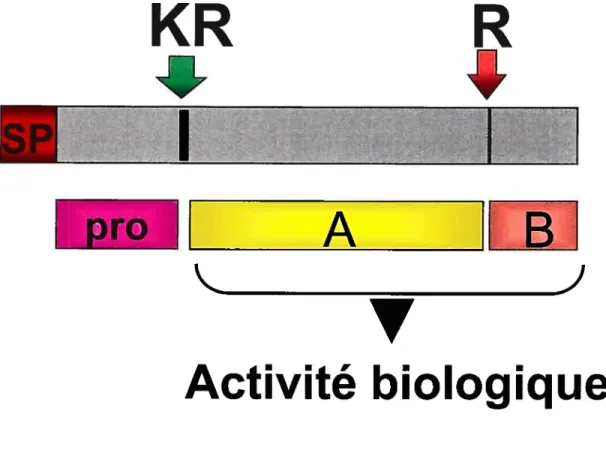

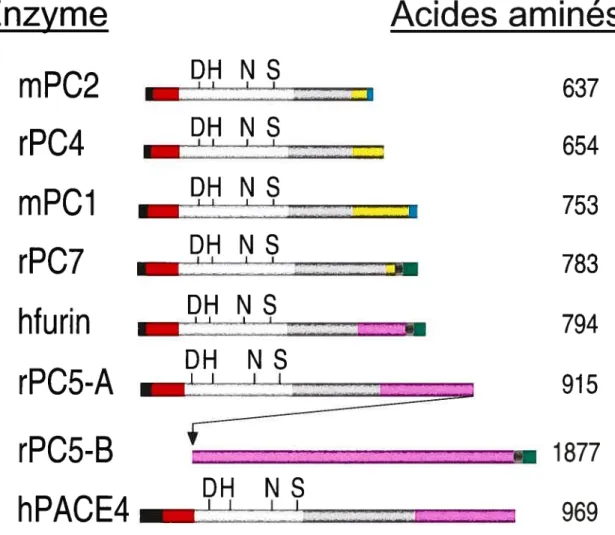
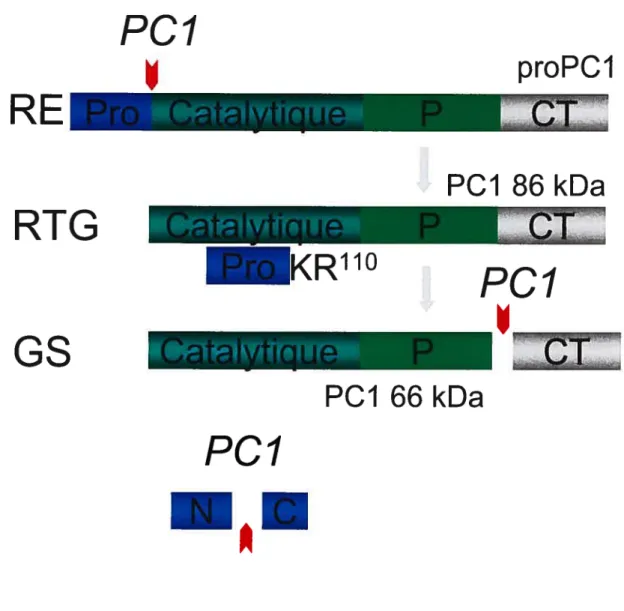

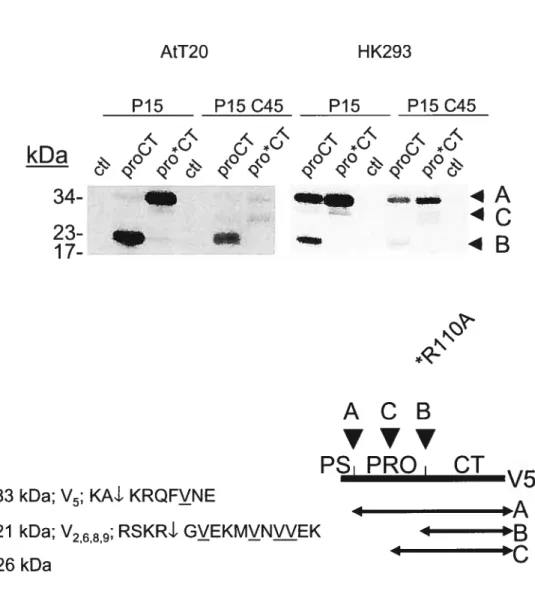
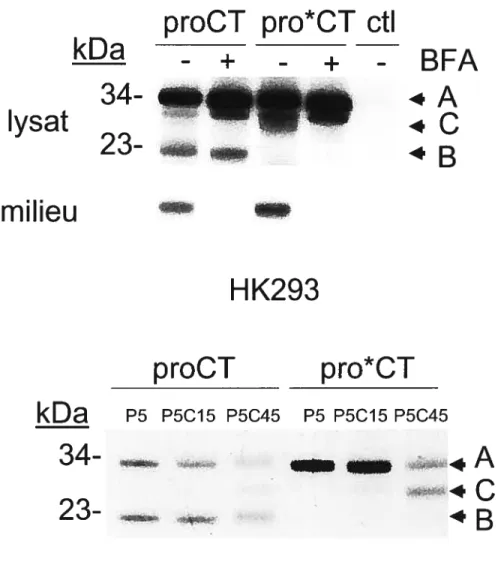
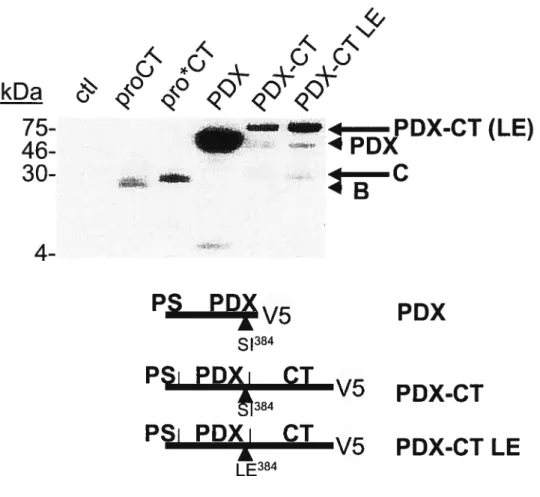
Documents relatifs
Cet article de synthèse présente les résultats des premières études ayant inves- tigué les effets de la pratique de la méditation chez le sujet âgé, dans différents
Sur un total de 1409 objectifs éducatifs du programme éducatif congolais du niveau primaire, seuls 78 (soit 5,53 %) concernent explicitement la créativité ou la synthèse
Ces résultats mettent en évidence (i) l’importance du stress du RE comme nouvelle voie de signalisation pro-apoptotique induite par les LDL oxydées dans les
Le magazine Do Rzeczy que nous avons choisi comme exemple polonais regroupe des commentaires se référant aux articles dans un numéro du magazine paru plus tôt ou à
Si l'hypothèse que FMRP joue un rôle commun aux autres RBPs impliqués dans la différenciation cellulaire et dans la synthèse protéique locale par son
Ju ji n g La P e ti te G re n o u il le Manon Emmanuelle B.. La phase inflammatoire 22 ° Le rôle du surfactant dans la défense de l’hôte 22 ° La production de
Our observations that acetylation and phosphor- ylation of eIF2α increase in parallel in response to ER stress when SIRT1 is inhibited or knocked out and that acetylation (K143R)
1. Etude de l’élevage dans le développement des zones cotonnières : le Mali. Mémoire Ing., Cnearc, Montpellier, France, p. L’élevage en voie d’intensification :
