Répartition spatiale et sélection d’habitat de nidification du
faucon pèlerin au Nunavut, Canada
Mémoire présenté
dans le cadre du programme de maîtrise en gestion de la faune et de ses habitats en vue de l’obtention du grade de maître ès sciences
PAR
© KRISTEN PECK
Composition du jury :
Martin-Hugues St-Laurent, président du jury, Université du Québec à Rimouski Joël Bêty, directeur de recherche, Université du Québec à Rimouski
Alastair Franke, codirecteur de recherche, Canadian Circumpolar Institute Chris Johnson, examinateur externe, University of Northern British Columbia
UNIVERSITÉ DU QUÉBEC À RIMOUSKI Service de la bibliothèque
Avertissement
La diffusion de ce mémoire ou de cette thèse se fait dans le respect des droits de son auteur, qui a signé le formulaire « Autorisation de reproduire et de diffuser un rapport, un mémoire ou une thèse ». En signant ce formulaire, l’auteur concède à l’Université du Québec à Rimouski une licence non exclusive d’utilisation et de publication de la totalité ou d’une partie importante de son travail de recherche pour des fins pédagogiques et non commerciales. Plus précisément, l’auteur autorise l’Université du Québec à Rimouski à reproduire, diffuser, prêter, distribuer ou vendre des copies de son travail de recherche à des fins non commerciales sur quelque support que ce soit, y compris l’Internet. Cette licence et cette autorisation n’entraînent pas une renonciation de la part de l’auteur à ses droits moraux ni à ses droits de propriété intellectuelle. Sauf entente contraire, l’auteur conserve la liberté de diffuser et de commercialiser ou non ce travail dont il possède un exemplaire.
À ma famille québécoise – vous savez qui vous êtes.
REMERCIEMENTS
Merci premièrement aux mes directeurs, Joël Bêty et Alastair Franke, et à Nicolas Lecomte de m’avoir guidé dans ce projet du début à la fin. Il n’aurait pas été possible sans votre patience et générosité. Merci aux gens de Rimouski et les laboratoires de Joël Bêty, de Dominique Berteaux et de Martin-Hugues St-Laurent d’avoir été aussi généreux avec l’amitié pour une fille de l’ouest qui ne parlait pas ben bon en arrivant. C’est à cause de vous que je n’ai pas quitté un mois après mon arrivée, et c’est à cause de vous que j’ai eu le grand plaisir d’apprendre le français et de vivre la belle vie québécoise. Je pars une autre personne que j’étais en arrivant ; je ne vous oublierai jamais. Merci au Conseil de recherches en sciences naturelles et en génie du Canada, à la Fondation W. Garfield Weston, à Mitacs Canada en collaboration avec Agnico-Eagle Mines, et à l’Université du Québec à Rimouski d’avoir fourni mon financement personnel pour ce projet.
Qujannamiik to the people of Igloolik for the warm welcome into your community, back in the days when this project was still unmolded clay. I know that we qallunaat can be annoying when we show up at the Northern with our cameras and silly grins, but I gained so much love and respect for the Inuit way of life and the beauty of being out on the land. Thanks especially to Michael Qrunnut - I would have surely have fallen into a crack in the sea ice without you (although we did still manage to sink the boat). Thanks for your patience and willingness to teach me a couple of things along the way, even if I slowed things down a bit. And thanks to Matt Fredlund, for the conversations through the bookshelf, the random fieldtrips to escape from the office, and the dog.
Finally, thanks to the people of Rankin Inlet and the Fredlunds, who welcomed us into their town and their home and made us feel like part of the family. Though our stay was brief, I will never forget your stories Steve!
RÉSUMÉ
Dans l’Arctique, une région immense et reculée, la répartition spatiale est bien connue seulement pour quelques espèces charismatiques ou d’importance économique. Les changements climatiques ont déjà commencé à influencer les écosystèmes arctiques; la température de l'air dans l’Arctique en Amérique du Nord a augmenté de 2 à 3°C depuis 1950. Dans ces régions éloignées, les pressions associées à l’exploitation des ressources naturelles peuvent aussi interagir avec le réchauffement climatique. Le coût prohibitif de la logistique impliquée dans les études menées loin des aires habitées dans l’Arctique fait en sorte qu'il est difficile d’améliorer notre connaissance des répartitions géographiques des espèces. Les modèles de répartition des espèces, dans lesquels l’information environnementale mesurée par télédétection est combinée à des occurrences pour prédire les répartitions d’espèces, peuvent servir à combler ce manque de savoir. Cette approche de modélisation pourrait aussi permettre d'étudier l’influence relative des facteurs environnementaux qui sont stables versus ceux qui sont variables (par exemple, la topographie versus le climat) dans la sélection d’habitat. L’objectif de notre étude était d’examiner la répartition spatiale des sites de nidification et la sélection d’habitat du faucon pèlerin (Falco peregrinus tundrius) au Nunavut, un vaste territoire nordique canadien. Nous avons comparé des données à long terme de répartition de nids aux données environnementales obtenues par télédétection, afin d'estimer la répartition des faucons pèlerins dans le reste du territoire. Nous avons ainsi prédit une probabilité relative d’occurrence de nids élevée dans les aires qui, à notre connaissance, n’ont jamais été échantillonnées. Nous avons aussi pu évaluer quels attributs environnementaux étaient les plus importants dans la sélection d’habitat de cette espèce à la limite nordique de sa répartition, soit le climat, la topographie, ou la productivité primaire. Dû à la grande échelle de cette sélection d’habitat, ces attributs environnementaux sont probablement ceux limitant la répartition nordique de cette espèce. Cette étude illustre l’utilité des modèles
développés à l’échelle régionale comme une première étape permettant d’investiguer la répartition des espèces dans les régions éloignées comme l’Arctique canadien. Nos résultats contribueront à une meilleure conservation et gestion d’une espèce arctique largement répandue.
Mots clés : Modèles de répartition des espèces, sélection d’habitat, faucon pèlerin de la toundra, base de données collaborative, changement climatique, l’échelle régionale, Nunavut, toundra, Arctique
ABSTRACT
Throughout the vast and remote area of the Arctic, the spatial distributions of only a few charismatic or economically important species are well known. Climate change has already impacted arctic ecosystems; air temperatures in the North American Arctic have increased by 2-3oC since the 1950s. The increasing pressures of resource development may also interact with a warming climate to place new pressures on these remote areas in the future. The prohibitive cost of logistics for studies outside of populated areas in the Arctic makes it difficult for researchers to improve the current knowledge of species ranges. Species distribution models, in which a sample of occurrences and remotely-sensed environmental information are combined to predict species distributions over areas with unknown species presence, may be one method of addressing this knowledge gap. This modelling approach can also explicitly describe the relative influence of stable versus changing environmental features (e.g. topography versus climate) in habitat selection. The objective of our study was to determine the nesting distribution and the broad scale habitat selection of the tundra peregrine falcon (Falco peregrinus tundrius) throughout most of Nunavut, a massive northern Canadian territory. We compared a long-term nest dataset with remotely-sensed environmental variables to estimate the habitat selection and distribution of nesting peregrine falcons throughout this northern territory. We not only predicted a high relative probability of nesting occurrences in areas that, to our knowledge, have never been surveyed, we were also able to rank the most important environmental features at the northern limit of this species’ distribution, such as climate, topography and primary productivity. Due to the broad scale at which these habitat features were selected, they likely have a limiting effect on the northern distribution of this species. This study demonstrates the utility of using regional-scale distribution modelling as a first step for investigating species ranges in remote regions like the Canadian Arctic. These results will
xvi
contribute to a better conservation and management of a widespread arctic species throughout its northern range.
Keywords: Species distribution models, habitat selection, tundra peregrine falcon, collaborative data base, climate change, regional scale, Nunavut, tundra, Arctic
TABLE DES MATIÈRES
REMERCIEMENTS ... ix
RÉSUMÉ ... xii
ABSTRACT ... xv
TABLE DES MATIÈRES ... xviii
LISTE DES TABLEAUX ... xx
LISTE DES FIGURES ... xxii
LISTE DES ANNEXES ... xxv
INTRODUCTION GÉNÉRALE ... 1
CHAPITRE 1 Répartition et sélection d’habitat de nidification du faucon pèlerin au Nunavut, Canada ... 12
1.1 RÉSUMÉ EN FRANÇAIS DU PREMIER ARTICLE ... 12
1.2 NESTING HABITAT SELECTION AND DISTRIBUTION OF THE PEREGRINE FALCON (FALCO PEREGRINUS TUNDRIUS) IN NUNAVUT,CANADA ... 14
1.3 ABSTRACT ... 14
1.4 INTRODUCTION ... 15
1.5 METHODS ... 19
1.5.1 Study area ... 19
1.5.2 Nest sites and occurrence cells ... 20
1.5.3 Comparison points ... 21
1.5.4 Habitat variables ... 22
1.5.5 Data analysis ... 23
1.6.1 Peregrine falcon distribution ... 26
1.7 DISCUSSION ... 26
1.8 CONCLUSION ... 30
1.9 REFERENCES ... 31
TABLES AND FIGURES ... 39
ANNEXE ... 46
CONCLUSION GÉNÉRALE ... 50
LISTE DES TABLEAUX
Table 1 : Source data (grey rows) and derived habitat variables (white rows) included in a peregrine falcon (Falco peregrinus) habitat selection analysis at the regional scale in Nunavut, Canada. All variables were calculated within a 10x10 km cell using the source data resolution. When appropriate, sample mean ( ) and standard deviation (s) of variables in “occurrence” and an equal number of “pseudo-absence” cells were reported. For “Coastal,” a binary variable, the percentage of cells classed as coastal was reported instead ... 39 Table 2 : Competing resource selection functions predicting the relative probability of occurrence of nesting peregrine falcons (Falco peregrinus) in Nunavut, Canada. Models were calculated using all known nests and a random subset of pseudo-absences of equal number. We ranked models by the lowest Akaike’s Information Criterion (AIC), ∆AIC, and AIC weight. Models represent variables in competing hypotheses: “Clim”= climate variables, “Topo”= topographic variables, “Bio”= proxy for prey availability. All variables except Coastal (which was binary) were centred on the mean and standardized by the standard deviation. For variable definitions, see Table 1 ... Erreur ! Signet non défini.1
LISTE DES FIGURES
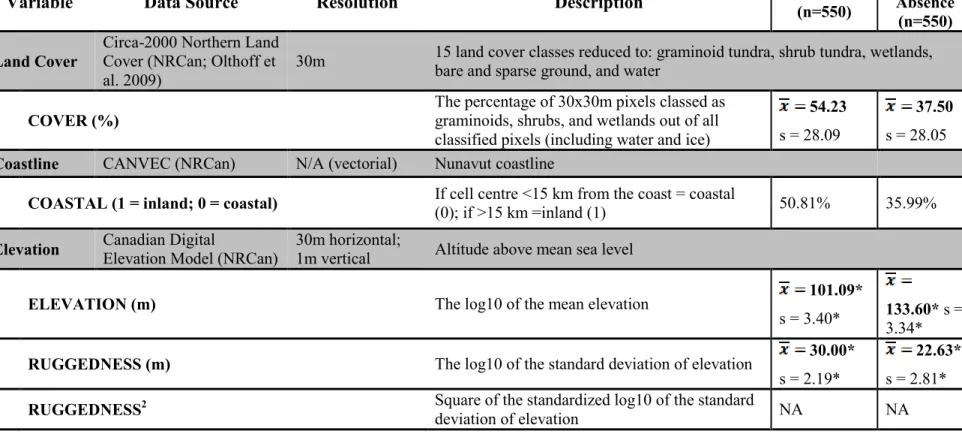
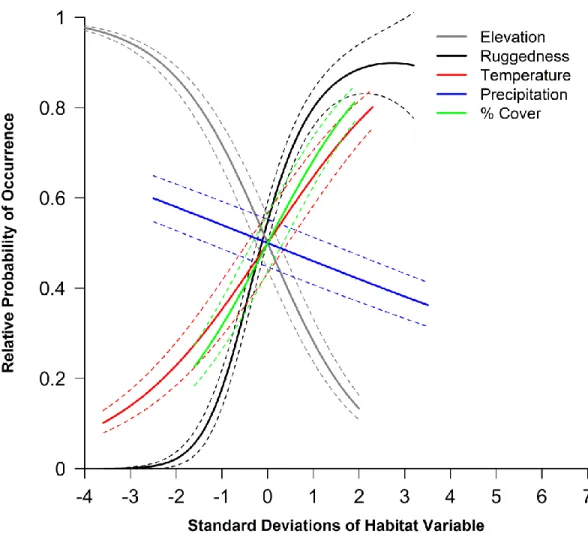
Figure 1 : The 1 110 known peregrine falcon (Falco peregrinus) nest sites, occupied at least once from 1951-2013, throughout the majority of Nunavut, Canada. Nest locations are from the Nunavut and Northwest Territories Raptor Database. The study area (light green) used in the analysis and subsequent predictions is bounded to the south by the tree line (broken green line) or by territorial borders (straight black lines), water bodies (in blue), and to the north by the Parry Channel (ca. 74°N). Areas lacking environmental data or with year-round ice were excluded from the analysis ... 38 Figure 2 : The individual effects of environmental variables on the relative probability of peregrine occurrence at the 100 km2 grain size in Nunavut, Canada, when all other variables are held constant. Environmental variables include average elevation (Elevation), terrain ruggedness (Ruggedness), mean summer temperature (Temperature), mean summer precipitation (Precipitation), and the percentage of productive cover (% Cover). All variables have been centred on their mean and standardized by their standard deviation. Dotted lines represent the 95% confidence intervals. For means and standard deviations of variables, see Table 1 ... 42 Figure 3 : Validation plot of the best model (in italics; lowest AIC among candidate models) describing the relative probability of nesting occurrence of peregrine falcons (Falco peregrinus) in Nunavut, Canada. The true positive rate (the rate at which the model correctly classifies presence cells) is plotted versus the false positive rate (the rate at which the model falsely classifies presence cells) and the area under the curve (AUC) reported in bold. The function was generated with 70% of the data and validated with 30%, bootstrapped 1000 times. The red dotted line is when the AUC is 0.50, or the classification is random ... 43 Figure 4 : Predicted relative probability nesting occurrence of the peregrine falcon (Falco peregrinus) in Nunavut, Canada. Nesting probabilities were derived from the best model explaining peregrine nest habitat selection based on the lowest AIC: P(Presence)~Temperature-Elevation+Ruggedness+Ruggedness2-Precipitation+ %Cover. Areas of year-round ice with an area greater than 1 km2 (stippled white), areas lacking environmental data (grey) and large water bodies (blue) were excluded
from the prediction area. The three regions of Nunavut are delimited in an inset in the bottom right ... 44
LISTE DES ANNEXES
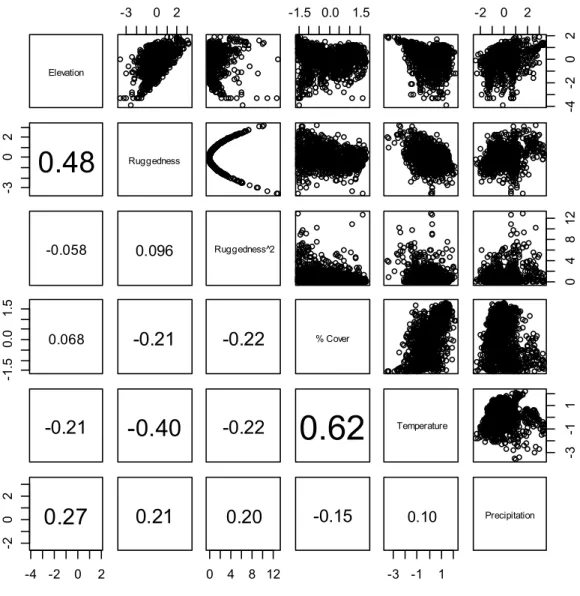
Figure A1 : Example illustration of the conversion of known peregrine falcon (Falco peregrinus) nest sites (red) to occurrence grid cells (orange) and the selection of pseudo-random grid cells (blue) for comparison around the community of Rankin Inlet, Nunavut, Canada. Throughout the study area covering the majority of Nunavut, 1 110 nest sites were reduced to 550 occurrence cells. For comparison, 550 pseudo-absence cells were randomly selected from background cells and re-sampled 10 times (see Methods for details) ... 45 Figure A2 : Distribution of nesting habitat variables at occurrence (blue, n=550) and one representative set of pseudo-absence (red with stripes, n=550) cells of peregrine falcons (Falco peregrinus) in Nunavut, Canada. All variables are centred on their mean and standardized by their standard deviation except for the variable “Coastal,” which is binary. For variable definitions, see Table 1 ... 46 Figure A3 : Pair-wise Pearson’s correlations and simple scatter plots between variables included in the analysis of habitat selection of the peregrine falcon (Falco peregrinus) in Nunavut, Canada. The font size of the correlation coefficient increases with the strength of the correlation. Data includes equal numbers presence and one representative set of pseudo-absence points (n=550 each). For variable definitions, see Table 1 ... 47 Table A1 : Example of variable values of one high and one low relative probability cell. Cells were randomly chosen from among the highest and lowest 0.05 relative probability; the “high” cell is from Bathurst Inlet south of Kugluktuk, and the “low” cell is from northwest Southampton Island. For variable definitions, see Table 1. Variables are presented in their original units. ... 48
INTRODUCTION GÉNÉRALE Les modèles de répartition d’espèces
Les modèles de répartition d’espèces sont des outils fréquemment utilisés en biogéographie de la conservation (Guisan et Thuiller 2005, Franklin 2013). Bien que le type d'analyses et les méthodes employées varient grandement, l'objectif est de créer un modèle empirique qui réfère à l'occurrence et l'abondance des espèces ou groupes d'espèces en fonction de variables environnementales (Guisan et Zimmermann 2000). Afin d'estimer la répartition d’espèces ou d’ensembles d'espèces, les modèles peuvent se baser sur une variété de sources de données, incluant des données issues des musées (Graham et al. 2004), des données paléontologiques (Pearman et al. 2008) ou portant sur la réponse physiologique aux facteurs environnementaux des organismes (Buckley et al. 2010). Étant donné que les modèles de répartition d’espèces s'emploient souvent à l’échelle de la région ou du paysage, ils sont efficaces pour produire des cartes de référence pour la répartition d’espèces se basant sur des données incomplètes, ce qui est souvent le cas pour les espèces rares, à faible densité ou se trouvant en régions isolées. Par exemple, ces cartes de répartition ont été utilisées pour améliorer la conservation du Courvite de Jerdon (Rhinoptilus bitorquatus), un limicole en voie de disparition pour lequel il existe peu d'observations (Jeganathan et al. 2004). En utilisant les traces laissées par ces oiseaux et les connaissances acquises sur leur habitat, les auteurs ont pu estimer leur répartition à partir des données de végétation obtenues par télédétection et mesurées sur le terrain. Szor et al. (2008) ont étudié la répartition et la productivité des tanières de renards arctiques (Vulpes lagopus) à un site d'étude dans le Haut-Arctique en lien avec la topographie, le microclimat et les ressources alimentaires. Cette étude a identifié les variables environnementales affectant à la fois la localisation des tanières et le succès reproducteur des individus utilisant les tanières (Szor et al. 2008). Dans une étude multi-espèces, Qi et al. (2009) se
2
sont intéressés à la relation entre l’environnement et deux espèces de pandas en danger provenant de populations sympatriques pour comprendre comment ils partagent et se séparent les ressources, et comment ces interactions pouvaient être modifiées par les changements environnementaux. Étudier la répartition des espèces n'implique pas simplement la production de cartes d'occurrence, mais le processus en soi permet de mettre de l'avant la manière dont les espèces interagissent avec leur environnement (Rushton et al. 2004). Par conséquent, les modèles de répartition d’espèces sont particulièrement pertinents pour la production d'information de base sur la relation entre les espèces et leur environnement en régions éloignées.
Sélection d'habitat
L'habitat se définit comme l'ensemble des ressources et des conditions où vivent un ou plusieurs organismes vivants (Hall et al. 1997, Morrison et al. 2001). Il est généralement déterminé par des contraintes individuelles ou spécifiques (Hall et al. 1997), de telle sorte que si nous savons où se trouve un individu, nous pouvons déterminer son utilisation de l'habitat (Morrison et al. 2001). Quand cette utilisation est comparée à l'habitat disponible (c.-à-d. ressources ou composantes environnementales auxquelles l'individu a accès; Hall et al. 1997), nous pouvons mesurer la sélection de l'habitat d'un individu. Un individu est sélectif s'il utilise une ressource ou une composante de l'habitat de manière disproportionnée en comparaison à sa disponibilité (Johnson 1980). La sélection de l'habitat est hiérarchique, et Johnson (1980) définit quatre ordres hiérarchiques de sélection allant du plus grossier au plus fin : le territoire géographique d'une espèce (premier ordre), la sélection saisonnière des aires utilisées à l'intérieur de ce territoire (second ordre; souvent considéré comme le domaine vital), la sélection des parcelles utilisées à l'intérieur de ce domaine (troisième ordre), et finalement, la micro-sélection à l'intérieur de ces parcelles (quatrième ordre). Bien que l'habitat soit fondamentalement sélectionné par l’individu, l'utilisation de l'habitat peut être mesurée à plusieurs niveaux organisationnels tels que la population ou l'espèce. Thomas et Taylor (1990) décrivent trois approches générales pour les recherches en sélection d'habitat : 1- l'utilisation et la disponibilité de l'habitat sont
communes pour tous les individus d'une aire d'étude; 2- l'utilisation de l'habitat est mesurée pour chaque individu, mais la disponibilité est commune pour tous les individus; et 3- l'utilisation et la disponibilité sont estimées pour chaque individu dans l'aire d'étude. Cette dernière approche permet d’identifier la variabilité de séléction entre les individus d’une population, tandis que la première approche identifie seulement la séléction moyenne de la population entière.
Bien que la terminologie entre les champs de fonctions de sélection de ressources et les modèles de répartition d’espèces se recoupent rarement (Mcdonald et al. 2013), les modèles qui estiment la répartition d’espèces en fonction de l'utilisation de l'habitat appartiennent généralement à la première approche selon Thomas et Taylor (1990). En d'autres termes, les modèles de répartition d’espèces ne mesurent habituellement pas l'utilisation de l'habitat par chaque individu, mais comparent davantage l'abondance ou l'occurrence des individus au sein d'une aire (souvent sous la forme d'une cellule dans une grille; Thuiller 2003) à l'habitat présent ou disponible (Guisan et Zimmermann 2000). L'habitat préférentiel est alors cartographié et l'occurrence de l'espèce est prédite là où l'habitat est présent. C'est ce qu'on appelle la règle de la correspondance avec l'habitat (habitat-matching rule; Cassini 2011), une des principales prémisses des modèles de répartition d’espèces.
L'influence de l'échelle
En plus de la complexité liée à l'ordre hiérarchique de la sélection d'habitat (Johnson 1980) et la variété d’approches de recherche en sélection d'habitat (Thomas et Taylor 1990), le processus de sélection de l'habitat diffère également en fonction de l'échelle considérée (Levin 1992). Les limites géographiques de répartition des espèces sont grandement déterminées par les patrons climatiques (Pearson et Dawson 2003), alors qu'à échelle spatiale fine, comme le domaine vital ou le site de reproduction, les individus sont plus susceptibles de faire leur sélection en fonction de la disponibilité des ressources alimentaires (Hutto 1985) ou de l'évitement des prédateurs (Chalfoun et Schmidt 2012). En général, la sélection à grande échelle spatiale devrait limiter la sélection à l’échelle fine
4
(Wiens 1989). Par exemple, si l'habitat à l’échelle fine pour le site de reproduction est disponible dans un secteur (quatrième ordre de sélection; Johnson 1989), mais que les limites de température d'une espèce sont surpassées (premier ordre de sélection), les individus ne pourront pas sélectionner parmi les sites de reproduction disponibles, car ils évitent déjà la région dans son ensemble à cause de la température. Rettie et Messier (2000) suggèrent que les facteurs ayant la plus grande influence sur la valeur adaptative devraient aussi être sélectionnés à grande échelle. Par exemple, une étude sur l'orignal (Alces americanus) au Québec a démontré que les individus sélectionnent les secteurs les moins protégés contre les prédateurs à l'échelle du paysage afin de combler en premier lieu la forte demande énergétique pour leur consommation, mais qu'à l'échelle du domaine vital, les femelles avec veau sélectionnent fortement les secteurs à faibles risques de prédation (Dussault et al. 2005). Dans ce cas, les ressources alimentaires et la prédation étaient tous les deux importants pour la sélection d'habitat chez l'orignal, mais le facteur limitant le plus leur répartition étaient les ressources alimentaires bien que les endroits à haut risque de prédation étaient évités à plus petite échelle. Lorsque les facteurs les plus importants pour la sélection de l'habitat d'une espèce sont altérés, cela peut entraîner des effets sur le succès des individus ou des populations. Par exemple, dans un secteur du Québec, les caribous (Rangifer tarandus caribou) évitaient les zones avec une présence humaine, mais depuis que ces zones sont grandissantes, les caribous ne peuvent plus complètement les éviter, ce qui constitue un facteur susceptible de contribuer au déclin de leur population (Leclerc et al. 2012). Par conséquent, les études de sélection de l'habitat à grande échelle devraient être en mesure d'identifier les attributs environnementaux qui ont le plus grand impact sur la valeur adaptative, ou les facteurs qui sont les plus prépondérants pour limiter la répartition d'une espèce. Identifier ces facteurs est utile pour la conservation ou la gestion des espèces, en particulier si les composantes de l'habitat sont susceptibles d'être affectées par les changements environnementaux futurs.
Climat et répartition des espèces
Le climat est l'une des forces les plus influentes agissant sur la répartition des espèces à l'échelle continentale (Pearson et Dawson 2003). De 1951 à 2012, la température mondiale à la surface a augmenté au taux de 0,12oC (variant de 0,08 à 0,14oC) par décennie (Stocker et al. 2013). La température mondiale moyenne à la surface du globe devrait augmenter de 1 à 2oC (variant de 0,4 à 2,6oC) à partir de la moitié du 20e siècle, selon les scénarios de concentration des gaz à effet de serre (Stocker et al. 2013). Bien que les précipitations soient plus difficiles à prédire, les modèles suggèrent une augmentation aux latitudes nordiques d'ici la fin du 21e siècle (Stocker et al. 2013). Étant donné qu'il s'agit d'une moyenne globale, certaines zones seront confrontées à des hausses de température supérieures comparativement à d'autres, comme les zones terrestres et celles des latitudes nordiques (Stocker et al. 2013). Ces changements de température ont déjà eu des impacts sur la répartition des espèces (Parmesan et al. 1999, Chen et al. 2011), et devraient continuer à influencer les répartitions futures (Guisan et al. 2013). La répartition des espèces répond différemment au climat, soit en se déplaçant, en se compressant ou en prenant de l'expansion (Guisan et al. 2013). Finalement, les espèces peuvent s'adapter aux changements dans l'aire de répartition actuelle si les déplacements sont entravés (Aitken et al. 2008). Par exemple, un modèle mécanistique avec lequel on a modélisé les contraintes physiologiques connues du Hêtre commun (Fagus sylvatica) en fonction d’une combinaison entre les changements de température de l’air et différents scénarios d’utilisation du paysage a démontré que cette espèce va probablement subir une contraction de son aire de répartition d’ici 2100 étant donné ses traits d’histoire de vie (i.e., un temps de génération élevé; Saltré et al. 2015). Pour certaines espèces, de nouveaux défis risquent d’apparaître par des modifications des interactions interspécifiques, telles que l’invasion par des compétiteurs ou l’éloignement des proies pour les prédateurs. Par exemple, le renard arctique est un prédateur endémique de l’Arctique qui se retrouve directement et indirectement surpassé par son congénère du Sud, le renard roux (Vulpes vulpes; Hersteinsson et MacDonald 1992). Hof et al. (2012) prédisent que la répartition du renard arctique en Fennoscandinavie sera réduite de 43% en raison de contraintes climatiques,
6
mais en considérant la diminution de l’amplitude des variations d’abondance de lemmings et l’expansion du renard roux, cette réduction s’élève à 56%. Il devient donc important de considérer que les effets des changements climatiques sur la répartition géographique des espèces peuvent affecter les espèces de maintes façons, soit en affectant directement les limites physiologiques d’une espèce ou indirectement par la répartition et l’abondance des proies, des compétiteurs et des prédateurs.
Répartitions actuelles des espèces
L’un des défis avec la prédiction des changements futurs de la répartition géographique des espèces est le manque d’information à propos de leur répartition actuelle. Dans plusieurs cas, comme avec les espèces rares ou présentes en régions éloignées, seulement une fraction des lieux utilisés par l’espèce sont connus, ce qui peut entraîner des défis pour la gestion des espèces (Whittaker et al. 2005). Afin de prédire les effets d’un changement du climat sur la répartition des espèces, nous devons en premier lieu établir où se trouve actuellement l’espèce. Par exemple, la répartition actuelle d’un perroquet cryptique vivant au sol dans l’ouest de l’Australie et pour lequel les exigences environnementales sont peu connues a été estimée en utilisant des données d’occurrence très limitées et des informations environnementales (Gibson et al. 2007). Cette étude a fourni une première étape vers la conservation de cette espèce cryptique et a mis de l’avant l’importance de divers facteurs environnementaux sur son occurrence. L’étude de la répartition actuelle des espèces peut également aider à orienter les efforts d’échantillonnage dans les endroits où l’information est manquante (Guisan and Thuiller 2005), peut aider les décideurs dans la désignation d’aires protégées (Margules and Pressey 2000), permet d’évaluer le risque des changements des caractéristiques clés d'un habitat par le développement (Guisan et al. 2013a), et fournit une comparaison afin de prédire les modifications de la répartition d’espèces en réponse aux changements climatiques (Thuiller 2004). Le manque d’information concernant la répartition actuelle des espèces peut être important dans les régions difficiles d’accès ou coûteuses à atteindre (Kamino et al. 2012), et ce phénomène est particulièrement marqué dans l’Arctique (Ims et al. 2013).
Répartition d’espèces de l’Arctique
L’Arctique fait partie des lieux qui ont été et vont être les plus touchés par les changements climatiques (Serreze et Francis 2006, Stocker et al. 2013). Non seulement les températures moyennes et les événements de précipitation ont déjà changé depuis les dernières décennies, mais le climat de l’Arctique est amené à se modifier de façon dramatique même d’après les scénarios de changements climatiques les plus modérés (Larsen et al. 2014). En plus des impacts des changements climatiques dans l’Arctique nord-américain, le développement industriel a également pris de l’expansion dans les dernières décennies. Seulement au Nunavut, un territoire nordique recouvrant 22% de la surface terrestre canadienne, le produit intérieur brut des mines et de l’extraction du minerai, du pétrole et du gaz a augmenté de 65% entre 2010 et 2013 (Government of Nunavut, Nunavut Statistics Dept. 2014). En se basant sur les politiques actuelles, le secteur des mines va probablement s’accroitre dans le futur (Bigio et al. 2015). De plus, le développement et l’exploration industriel dans le Nord risquent d’augmenter dans le futur au fur et à mesure que les changements environnementaux donneront un plus grand accès aux voies maritimes navigables et une fenêtre d’exploitation saisonnière plus étendue (Pearce et al. 2011). La détection et la gestion des impacts de ces changements globaux sur les écosystèmes terrestres nordiques pourront, par contre, être limitées par le manque d’information de base sur les espèces. Quelques études à long terme dans l’Arctique canadien ont détecté des changements temporelles à l’échelle de la population et des communautés (p. ex. Franke et al. 2010, Trefry et al. 2010, Gauthier et al. 2013), mais les coûts et les difficultés logistiques pour mener des études à grande échelle limitent la portée des conclusions de ces recherche (Foy et al. 2014). En effet, les études à grande échelle portant sur la répartition d’une espèce particulière ou la diversité des espèces sont plutôt rares dans l’Arctique nord-américain, sauf pour quelques espèces charismatiques ou à intérêt économique élevé (p. ex. caribou: Yannic et al. 2014; bœuf musqué: Campbell et al. 2012; ours polaire: Wilson et al. 2014). Même les grandes bases de données issues de la science citoyenne tel que eBird manquent d’information concernant les régions nordiques puisque les efforts d’observation sont biaisés positivement vers les secteurs à plus forte
8
densité de personnes (Kelling et al. 2012). Étant donné l’étendue des changements qui affecteront les écosystèmes nordiques, des outils pour les gestionnaires de la faune (tel que des modèles de répartition d’espèces) doivent être développés pour détecter et limiter les impacts des changements globaux futurs sur les espèces à risque.
Le faucon pèlerin
Le faucon pèlerin (Falco peregrinus) est un prédateur aviaire au sommet des réseaux trophiques et dont les aires de reproduction sont largement répondu à l’échelle mondiale, s’étendant jusqu’à l’océan Arctique (Ratcliffe 1980, White et al. 2002). La sous-espèce F. p. tundrius se reproduit au nord de la limite des arbres en Amérique du Nord (White et al. 2013), où elle se nourrit en milieu arctique terrestre et marin (Court et al. 1988, L’Hérault et al. 2013). Cette sous-espèce migre vers l’Amérique centrale et l’Amérique du sud pendant la saison non reproductive, voyageant donc en moyenne 8200 km pour retourner sur son site de reproduction (Fuller et al. 1998b). Les populations des sous-espèces tundrius et anatum ont décliné vers la moitié du 19e siècle en Amérique du Nord principalement à cause de l’introduction de pesticides organochlorés (Cade et al. 1971). À l’aide de programmes de réintroduction et la prohibition de plusieurs pesticides au Canada et aux États-Unis (Heinrich 2009), les populations de faucon pèlerin ont récupéré grandement en Amérique du Nord depuis les années 1970. Toutefois, ils conservent le statut d’espèce préoccupante au gouvernement fédéral étant donné leur sensibilité aux contaminants environnementaux durant la saison de reproduction (Cooper et Beauchesne 2007). Des études récentes continuent de trouver des niveaux considérables de contaminants environnementaux chez des individus sauvages de cette espèce, mais ce ne soit pas suffisant pour altérer leur développement ou leur succès de reproduction (Franke et al. 2010, Smits et Fernie 2013). Des campagnes d’échantillonnage répétées ont lieu chaque cinq ans dans des secteurs spécifiques pour continuer le suivi des populations de cette espèce (Canadian Peregrine Falcon Survey; Rowell et al. 2003). En 2007, le Comité sur la situation des espèces en péril du Canada (COSEPAC) a estimé la taille de la population
minimale des sous-espèces anatum/tundrius à 1168 individus matures, dont seulement 119 des individus proviennent de la sous-espèce tundrius (Cooper et Beauchesne 2007). Cependant, le comité explique que ce nombre est probablement sous-estimé, car la plus grande partie de la répartition du faucon pèlerin se trouve dans les régions toundriques éloignées (Cooper et Beauchesne 2007).
Bien que le faucon pèlerin toundrique soit bien suivi dans certaines régions spécifiques, il y a relativement peu d’information sur sa répartition spatiale ou sur sa démographie à grande échelle. La répartition du faucon pèlerin dans le nord du Canada a été estimée quelques fois dans le passé grâce aux sites de reproduction connus et aux avis d’experts. Sur l’entièreté de la région du Nord canadien, des latitudes 55 à 73oN, Fyfe (1969) a estimé la superficie d’habitat de nidification optimal des faucons pèlerins à 128164 km2, définit à partir de caractéristiques topographiques et la proximité de cours d’eau. Cette estimation fut basée sur des observations de faucons et de leurs habitats effectuées par plusieurs biologistes et observateurs travaillant dans différentes régions du nord canadien. Plus récemment, une carte de la densité des rapaces dans la région du Kivalliq au Nunavut a été créée à partir de tous les sites de nidification connus pour les oiseaux de proie de la région (Campbell et al. 2012). Cependant, étant donné que l’échantillonnage des sites de nidification n’a pas couvert l’aire d’étude de façon uniforme, il s’agit probablement d’une sous-estimation de la répartition actuelle des rapaces. Ces deux méthodes d’estimation de la répartition (c.-à-d. à partir d’avis d’experts et d’une utilisation strict des données d’occurrences connues) ont chacune leur mérite, mais elles ont le potentiel de sur- ou sous-estimer la répartition spatiale. De plus, il est difficile de répéter des estimations se basant sur les avis d’experts en utilisant de nouvelles informations puisque ces estimations sont qualitatives. La modélisation de la sélection d’habitat et de la répartition des espèces se base sur une combinaison de ces deux approches : il est en effet important d’inclure dans le modèle des informations jugées pertinentes sur la base de notre connaissance de l’habitat de l’espèce (Austin 2007) ainsi que les occurrences locales des espèces pour comprendre la sélection d’habitat d’une espèce.
10
Objectifs et prédictions
Les objectifs de cette étude sont en premier lieu d’étudier la sélection de l’habitat de nidification du faucon pèlerin au sein du biome de la toundra dans l’est de l’Arctique canadien, et en second lieu d’utiliser des informations sur leur habitat préférentiel pour modéliser leur répartition actuelle durant la nidification sur l’ensemble de l’aire d’étude. Nous anticipons que la sélection de l’aire de nidification chez le faucon dans l’Arctique sera fortement influencée par des caractéristiques topographiques étant donné que la disponibilité des parois de falaise était considérée importante dans toutes les autres études de sélection de l’habitat du site de nidification chez le faucon à plus petites échelles spatiales (Gainzarain et al. 2000, Wightman and Fuller 2005, Brambilla et al. 2006, Bruggeman et al. 2015). Étant donné que les variables climatiques sont souvent des déterminants clés pour la répartition des espèces à l’échelle régionale et du paysage (Pearson et Dawson 2003), et puisque notre aire d’étude se trouve à la limite nordique de leur répartition mondiale, les faucons devraient également préférer les zones avec des températures relativement chaudes. Il a été démontré que les événements de fortes précipitations ont un effet négatif sur la productivité du faucon pèlerin dans le Bas-Arctique (Anctil et al. 2013), par conséquent nous avons anticipé que les faucons préféreraient les zones avec moins de précipitation estivale. De plus, Jenkins et Hockey (2001) suggèrent que la disponibilité des proies peut être un facteur déterminant pour la densité de faucons à grande échelle. Toutefois, nous avons une connaissance très limitée de l’abondance des proies à grande échelle dans le Nord. Il se peut également que ce soit moins instructif d’utiliser les estimations d’abondance de proies souvent mesurées à court terme, étant donné que l’abondance des oiseaux et petits mammifères peut varier grandement d’une année à l’autre dans le Nord (Gauthier et al. 2013, Robinson et al. 2014). Par conséquent, nous avions prédit que les faucons auraient une réponse indirecte positive aux régions relativement productives en terme de végétation (toundra graminéenne, présence de couvert arbustif et milieux humides). À l’échelle régionale, la végétation peut être le principal facteur déterminant la productivité total de l’écosystème (Callaghan et al. 2004). Nous avons également tester les effets d’autres variables environnementales qui étaient
importantes pour la sélection d’habitat et la productivité du faucon pèlerin dans d’autres régions : la proximité de la côte (L’Hérault et al. 2013), et l’élévation moyenne (Sergio et al. 2004). Avec notre connaissance des habitats préférés par les faucons pèlerins à grande échelle, nous avons estimé leur occurrence sur l’ensemble du territoire du Nunavut, incluant les zones où une information de suivi est manquante.
CHAPITRE 1
RÉPARTITION ET SÉLECTION D’HABITAT DE NIDIFICATION DU FAUCON PÈLERIN AU NUNAVUT, CANADA
1.1 RÉSUMÉ EN FRANÇAIS DU PREMIER ARTICLE
Dans les régions éloignées, la répartition spatiale de la plupart des espèces est inconnue, ce qui représente un défi pour leur gestion et conservation. Le faucon pèlerin (Falco peregrinus tundrius) est une espèce dont le statut est préoccupant dans le Nord. Cette dernière fait fréquemment l’objet de suivis dans le cadre de projets industriels à cause des obligations légales des entreprises en termes d’études d’impacts environnementaux. Cependant, mis à part ces études à l’échelle locale, il n’y a aucune évaluation rigoureuse des conditions nécessaires pour la nidification du faucon pèlerin dans le Nord à plus grande échelle. Puisqu’il y a très peu de connaissances a priori des risques de rencontrer des faucons reproducteurs dans l’Arctique, l’objectif de notre étude est d’investiguer la sélection d’habitat de ce prédateur pour ensuite prédire la répartition des nids au Nunavut, un vaste territoire canadien (>2M km2) comprenant une large portion du biome de la toundra arctique. Nous avons utilisé 60 ans de données de suivi de nidification et des informations sur l’habitat obtenues par télédétection afin de modéliser la sélection d’habitat des faucons à l’échelle régionale (100 km2). Tel que le laissait présager la littérature, les faucons pèlerins préfèrent nicher dans les aires dont la topographie est la plus accidentée. Cependant, nous avons aussi découvert qu’ils préfèrent une température relativement élevée, un haut pourcentage de végétation productive, une altitude moyenne basse et qu’ils évitent les zones avec des précipitations relativement élevées. De plus, la modélisation de leur répartition a permis de déterminer des aires de haute probabilité relative d’occurrence, incluant des zones qui n’ont jamais été inventoriées. Connaître quels types d’habitat les faucons pèlerins choisissent aidera à prédire leur réponse aux utilisations futures du
territoire et aux changements climatiques. Ces connaissances permettront aux gestionnaires de la faune de mieux identifier les aires prioritaires à protéger et de mieux anticiper les interactions potentielles avec le secteur industriel en croissance au Nunavut.
Mots clés : Arctique, sélection d’habitat, télédétection, modèle de la répartition d’espèce, prédateur d’un niveau trophique supérieur, faucon pèlerin de la toundra
Cet article, intitulé « Nesting habitat selection and distribution of the peregrine falcon (Falco peregrinus tundrius) in Nunavut, Canada », est en attente de soumission dans un journal scientifique, potentiellement « Biological Conservation ». Il s'agit d'une collaboration avec mon directeur Joël Bêty, professeur au département de biologie, chimie et géographie à l'Université du Québec à Rimouski, Alastair Franke, investigateur principal du programme de recherche Arctic Raptors, et Nicolas Lecomte, professeur au département de biologie à l’Université de Moncton. En tant que première auteure, ma contribution à ce travail fut de générer l’idée du projet, d’élaborer les hypothèses, de récolter une partie des données sur le terrain, de faire les analyses statistiques et de rédiger l'article. Dr Alastair Franke, second auteur, a fourni une partie des données, a aidé à la formulation de la question de recherche, a aidé à établir les grandes lignes des méthodes et a aidé à la révision de l’article. Dr Nicolas Lecomte, troisième auteur, a aidé au développement de l’idée, a fourni un lien avec le Gouvernement du Nunavut pour l’utilisation de leurs données, a aidé à établir les méthodes et les analyses et finalement, a aidé à la révision de l’article. Dr Joël Bêty, dernier auteur, a été impliqué dans toutes les étapes du processus menant à cet article. Il a aidé à la formulation de la question de recherche et des hypothèses, au développement de la méthodologie ainsi qu'à la révision de l'article. Une présentation portant sur le contenu de cet article a été faite en décembre 2014 à Ottawa dans le cadre d’« Arctic Change », une réunion scientifique internationale d’ArcticNet, de même qu'au congrès annuel du Centre d'études nordiques tenu à Québec en février 2015.
14
1.2 NESTING HABITAT SELECTION AND DISTRIBUTION OF THE PEREGRINE FALCON
(FALCO PEREGRINUS TUNDRIUS) IN NUNAVUT,CANADA
Peck, Kristen1*; Franke, Alastair2; Lecomte, Nicolas3,4; Bêty, Joël1
1Centre d’études nordiques et Département de chimie, biologie et géographie, Université du Québec à Rimouski, Rimouski, Québec
2Arctic Raptor Project, Rankin Inlet, Nunavut
3Chaire de recherche du Canada en Écologie Polaire et Boréale, Département de biologie, Université de Moncton, Moncton, New Brunswick
4Centre de la Science de la Biodiversité du Québec, Département de chimie, biologie et géographie, Université du Québec à Rimouski, Rimouski, Québec
*kristen.peck01@uqar.ca 1.3 ABSTRACT
In remote areas such as the Canadian Arctic, breeding distributions of many species are not well known, or even unknown, making it difficult for wildlife managers to predict how species will be impacted by land-use or climate changes. One species with a status of “special concern” in the North, the peregrine falcon (Falco peregrinus tundrius), is often monitored by industrial projects due to the legal mandates of environmental assessment certificates. Besides these local area studies, there has been no rigorous regional scale evaluation of peregrine falcon nesting distribution in the North. Since there is little to no a priori knowledge of the risk of encountering peregrine falcon nesting sites in most of the Arctic, the objective of our study was to determine the nesting habitat selection of this top predator and to predict its nesting distribution throughout most of Nunavut, a massive northern Canadian territory (>2M km2) encompassing a large and varied portion of the tundra biome. Using a 60-year dataset and remotely-sensed habitat information, we modeled the nesting habitat selection and the likely nesting distribution of peregrine falcons
at the regional scale (grain size of 100 km2). As expected from previous studies, peregrine falcons selected nests in areas with the most rugged topography. However, we also found that higher than average summer temperatures, greater percentages of productive vegetation, low mean elevations, and relatively low mean precipitation were also selected. When their expected distribution was mapped, our model identified several areas of high relative probability of peregrine falcon occurrence, including areas that, to our knowledge, have never been surveyed in the past. Knowing which environmental criteria peregrine falcons use to select their nesting areas may help to anticipate their responses to future land-use and climate changes in areas without previous studies in Nunavut. Such knowledge will allow managers to identify the areas that are the most critical for monitoring and mitigation, as well as the likelihood of future interactions between this species and the growing industrial sector in Nunavut.
Key words: Arctic, habitat selection, remote sensing, species distribution model, top-predator, tundra peregrine falcon
1.4 INTRODUCTION
The Arctic is among the places that have been, and will continue to be, the most affected by climate change. Not only has the mean surface air temperature in much of northern North America already gained an average of 2-3oC since the 1950s (ACIA 2004, Screen and Simmonds 2010), the climate in the Arctic is expected to change dramatically in the future in even the most conservative climate change scenarios (Larsen et al. 2014). In addition to the impacts of a changing climate, industrial development has also expanded in the North American Arctic in recent decades. Northern industrial development and exploration are also expected to expand to new areas as environmental changes allow greater access to marine shipping routes and a longer seasonal exploration window (Pearce et al. 2011). Detecting and planning for the impact on northern terrestrial ecosystems from multiple pressures may be limited by a lack of baseline information for many species in the North.
16
Indeed, regional or landscape scale studies of species diversity and distribution are rare in the North American Arctic, except for a few charismatic or economically important species (e.g. caribou and muskoxen (Campbell et al. 2012, Yannic et al. 2014), wolves (Heard and Williams 1992), wolverines (Copeland et al. 2010), and polar bears (Wilson et al. 2014)). Large citizen-science databases like eBird are also limited by the low density of data contributors in most areas of the North (Kelling et al. 2013) and thus are too limited in sample size to create accurate distribution maps. Several long-term studies in the terrestrial Canadian Arctic have captured species trends at the population or community level (e.g. Franke et al. 2010, Trefry et al. 2010, Gauthier et al. 2013), but the cost and logistical difficulty of conducting studies in the North limits the spatial extent of such research (Foy et al. 2014). Given the scale of changes that will affect northern ecosystems, tools to determine current species distributions and habitat selection are required to adequately detect and respond to future global changes.
Species distribution models are commonly-used research tools in conservation biogeography (Guisan and Thuiller 2005, Franklin 2013). Although the methods vary widely, the goal is to create an empirical model relating the occurrence, abundance, or physiological response of a species, or a group of species, to environmental features (Guisan and Zimmermann 2000). Since species distribution models are often applied successfully to broad scales, they are ideal for producing baseline species distribution maps at a coarse resolution, yet are powerful enough to delineating how species interact with their environment (Rushton et al. 2004). These analyses are often based on imperfect datasets, which is often the case when data are gathered opportunistically or for alternate purposes, for example, museum records (Graham et al. 2004), paleo records (Pearman et al. 2008), or mechanistic responses to environmental stimuli (Buckley et al. 2010). In remote locations, species distribution models are a viable option for defining the relationship between species and their environments at the regional scale by using various sets of information accumulated over time. This is especially useful in the tundra biome, since human centres are widely dispersed and high costs limit travel.
The peregrine falcon (Falco peregrinus, hereafter referred to as peregrines) is an avian top predator with a worldwide breeding distribution extending as far north as the Arctic Ocean (Ratcliffe 1980, White et al. 2002). The subspecies F. p. tundrius nests in the tundra biome of northern North America, where it forages in both terrestrial and marine arctic environments (Court et al. 1988, L’Hérault et al. 2013). Studies of peregrines in northern North America have monitored long-term population trends (Franke et al. 2010, Ritchie and Shook 2011), the influence of different environmental and human features (Anctil et al. 2013, Coulton et al. 2013, L’Hérault et al. 2013) or assessed their occupancy, habitat and natural history (Court et al. 1988, Carrière and Matthews 2013). There have been widespread efforts to assess population trends following population crashes due mainly to organochlorine contamination in the mid-1900s (Cade et al. 1971). Following the banning of harmful chemicals in Canada and the United States and re-introduction programs, this species has seen a significant recovery and de-listing from endangered and threatened species lists (Mesta 1998, White et al. 2002). Following its de-listing, 5- to 10-year monitoring plans were proposed to document its recovery (Mesta 1998). However, these surveys have mainly been recurring local studies (e.g. the Canadian Peregrine Falcon Survey; Rowell et al. 2003), rather than broad scale attempts to define peregrine distribution or breeding habitat. Peregrine occurrence and densities in northern Canada were estimated using known nest locations and expert knowledge in the 1960s (Fyfe 1969), but quantitative spatial methods were not used. More recently, a map of overall raptor density in the Kivalliq region of Nunavut was created based on densities of all known nest sites (Campbell et al. 2012), but since the sampling of nest sites was patchy, this was likely an underestimate of actual peregrine distribution. Rather than conducting further surveys to fill in knowledge gaps, we used an existing database of nesting information gathered from a variety of sources (Peck et al. 2012) in a species distribution model to estimate the current distribution of peregrines in the northeast Canadian Arctic.
The objectives of this study were to investigate habitat selection of peregrines at the scale of the home range in the tundra biome of the eastern Canadian Arctic, and then to use selected habitats to estimate their breeding distribution throughout the study area. We
18
expected that peregrine home range selection would be strongly influenced by topographical variability, as availability of nest sites was consistently important in peregrine habitat selection studies at smaller scales (Gainzarain et al. 2000, Wightman and Fuller 2005, Brambilla et al. 2006, Bruggeman et al. 2015). Since climatic variables are often key determinants of the distribution of species at regional or landscape scales (Pearson and Dawson 2003), and since this area is at the northern limit of this species’ distribution, we predicted that peregrines should prefer areas with relatively warmer temperatures. In contrast, extreme precipitation events have been shown to negatively influence the productivity of peregrines in the Low Arctic (Anctil et al. 2013). We thus expected that peregrines would prefer to breed in areas with overall less summer precipitation. Jenkins and Hockey (2001) suggest that prey availability may be a determining factor for peregrine falcon density at the regional scale. However, prey abundance can be difficult to estimate in the North, since there is often a huge variation in bird and small mammal abundance from one year to the next, even within the same ecosystem type (e.g. Gauthier et al. 2013). Therefore, we predicted that peregrines would have an overall positive response to areas with productive vegetation (graminoid tundra, shrubby tundra, and wetlands). Because vegetation is a driver of overall ecosystem productivity at this scale (Callaghan et al. 2004), and given the long temporal scale of our nesting data, we used the percent cover of these land classes as an indirect proxy for prey availability. We also tested the effects of other environmental variables that were important to peregrine habitat selection or productivity in other areas: the proximity to the coast (L’Hérault et al. 2013), and the mean elevation (Sergio et al. 2004). Using the knowledge of which habitats peregrine falcons selected at the broad scale, our final step was to estimate their relative probability of occurrence throughout the territory of Nunavut, including in areas where survey information was lacking.
1.5 METHODS
1.5.1 Study area
Our study area covers approximately 1 689 000 km2 (~80%) of Nunavut and consists of three Canadian ecozones: the Arctic Cordillera, the Northern Arctic, and the Southern Arctic (Marshall 1999), and all arctic bioclimatic subzones except the coldest, zone A (Walker et al. 2005). We included areas above the tree line according to Olthof et al. (2008), except for some islands north of the Parry Channel where survey effort was limited to absent (Figure 1). Some islands in the southern Hudson’s Bay (e.g. the Belcher Islands) were also excluded due to the lack of environmental information, as were some locations where remote sensing data were absent (e.g. a small strip of the Brodeur Peninsula on Baffin Island, see Figure 1). Due to some discrepancies among the limits of the environmental variables, we buffered the coastline within the study area by 1 km to be sure to include all land areas. Throughout our study area, the average summer temperature (May to Aug.) is 1.5oC (range from -16.2oC May minimum to a 17.2oC July maximum) and the average summer precipitation is 26.9 mm (range: 8.3 to 60.5 mm; calculated from Worldclim climate data [averaged from 1950 to 2000]; Hijmans et al. 2005). A large proportion of the study area is coastal but is continental to the southwest where it borders the Northwest Territories. The highest elevation, 2133 m, is found on Baffin Island (Natural Resources Canada 2000). The dominant vegetation land classes are: sparsely vegetated bedrock, prostrate dwarf shrub, and barren ground (25.4%, 11.9%, and 10.0% of land area, respectively; Olthof et al. 2008). Overall, the study area covers most of the tundra habitats and management areas within Nunavut, and is an appropriate scale to study a widely-distributed and highly mobile species like the peregrine falcon (Guisan and Thuiller 2005).
20
1.5.2 Nest sites and occurrence cells
We used data from the Nunavut and Northwest Territories Raptor Database, which contains raptor nest site records from the 1950s to the present (Shank 1997, Poole 2011, Peck et al. 2012). Since raptors often re-use nesting territories year after year (Booms et al. 2011, Viter 2013), this database is structured by the nest site location rather than by individual bird (Peck et al. 2012). However, different raptor species can also occupy the same site from one year to the next, thus there are nest visit records documenting the yearly breeding species and reproductive information, in addition to the more permanent nest site information. As of 2013, there were 3 861 total raptor nest sites in this database (Nunavut: 2 469, Northwest Territories: 1 338, Alberta: 51, Yukon: 3) and 18 473 visit histories (Nunavut: 11 377, Northwest Territories: 5 164, Alberta: 1 929, Yukon: 3). For the records in which nest visit methods were noted (15 099 records), 81% were surveyed by helicopter, 11% by ground transport, 8% by boat and <1% by fixed-wing aircraft. Breeding peregrines were found during 3 428 nest visits in Nunavut from 1951-2013. However, this included the same nesting sites re-used over multiple years. In our analysis, we included sites in which peregrine falcon occupancy was confirmed at least once from 1951-2013. The total number of unique peregrine falcon nesting sites in our study area was 1 110 (Figure 1).
Nesting data were gathered by different sources and search effort was not recorded in the Nunavut and Northwest Territories Raptor Database. Nest locations were therefore spatially clumped, with a disproportionately high density of sites in some well-surveyed areas of Nunavut. There are several ways to account for biased sampling efforts, including down-weighting records in areas that received more survey effort, adding more data by surveying areas that are under-represented, or sampling background or pseudo-absence data only from areas that were likely surveyed (e.g. Phillips et al. 2009). In the case of peregrine nesting habitat selection in Nunavut, it would be logistically difficult to add data with targeted surveys due to the spatial and temporal spread of information included in this analysis. For this reason, we simplified the nesting data (following Thuiller 2003) into occurrence cells on a 10x10 km grid to account for this spatial sampling bias. If at least one
nest fell inside the cell, it was considered an occurrence (see Figure A1 for an example). This 100 km2 cell size was chosen to represent an approximate and conservative size of breeding peregrine home range in the North (L’Hérault et al. 2013, Sokolov et al. 2014, Franke, unpublished data). According to Guisan et al. (2007), our predictions should be robust even if the cell size was varied 10-fold. This transformation to grid cells reduced the weight of nests found in areas that were searched more intensively, thereby accounting for some of the spatial autocorrelation and sampling bias. It also masked any spatial error in the nest position caused by variable search methods (e.g. helicopters versus ground surveys) and the variable accuracy of site recording technology (e.g. physical maps versus handheld GPS) over the years. After this reduction of data, 550 peregrine occurrence grid cells remained of the original 1 110 nests sites (approximately 50% of total nest sites), which represents a small proportion of the study area (prevalence: 550/41 440 = 0.01 of total grid cells).
1.5.3 Comparison points
Search effort and survey tracks were not available to determine areas in which peregrine nest sites were absent; we therefore generated pseudo-absence cells to compare to occurrence cells. Pseudo-absences were generated throughout the entire study area since we considered all of the study area available to breeding peregrine falcons. Given the distance that individuals of this northern subspecies travel during migration (Fuller et al. 1998a) and the delimitation of the boundaries of the study area to include only areas with nest records, this assumption is likely valid. This comparison is less robust than comparing occurrence and true absence locations, but the detection of animals or animal sign is rarely perfect and true absences can be difficult to determine (Mackenzie et al. 2003, Booms et al. 2010, Lobo et al. 2010). Pseudo-absence points were sampled from non-occurrence cells throughout the study area and re-sampled 10 times to cover a wider range of background environmental variation. Environmental information from an equal number of occurrence and pseudo-random cells were compared in each habitat selection model. This comparison was repeated with each of the 10 pseudo-absence datasets.
22
1.5.4 Habitat variables
We chose environmental variables based on previous studies of peregrine habitat selection and observations of peregrine falcons in Nunavut. Austin (2002) suggests that direct variables, or variables with a direct effect on a species’ biology, should be used over indirect variables whenever possible. In the case of peregrines, direct variables might include: the availability of nest site substrate, prey availability, or weather extremes. However, at the regional scale, species distributions are often driven indirectly by climate, while at the local scale occurrences are driven more by direct, small-scale processes such as interspecific competition and micro-topography (Pearson and Dawson 2003). The large scale of our study led us to choose several variables that were indirectly related to peregrine site occupancy as proxies of direct variables. For example, we included overall terrain ruggedness instead of nest site availability, and primary productivity instead of prey availability. The biological effects of indirect variables are more challenging to interpret than variables with a direct effect on the species (Austin 2002), but at this scale of study broad signals can still be detected and interpreted with care.
All original environmental information was transformed into model variables in 10x10 km occurrence or pseudo-absence cells (Table 1). The standard deviation of elevation (Ruggedness [m]), squared standard deviation of elevation (Ruggedness2 [m2]), and mean elevation (Elevation [m]) were derived from Canadian Digital Elevation Data (Natural Resources Canada 2000); the percentage of productive land cover classes (Cover [%]: graminoids, shrubs and wetlands - we assume greater prey availability in high productivity land classes [Callaghan et al. 2004]) out of all land classes including water and ice was derived from the Northern Land Cover of Canada: Circa 2000 (Olthof et al. 2008); mean summer temperature (Temperature [oC]: May-Aug., or the entire breeding period) and precipitation (Precipitation [mm]: May-Aug.) were calculated from Worldclim data (monthly averages from 1950-2000; [Hijmans et al. 2005]); and whether the cell was inland or coastal (Coastal: >15 km or ≤15 km from the centre of the cell to the nearest coast, respectively) was calculated using CanVec political boundaries (Natural Resources Canada
2014). The cut-off of 15 km from the coast was used since there would be at least one full cell (or estimated home range) between the centre of the cell and the coast. We attempted to represent this variable both as truncated after 15 km and as a decay variable, but the distribution of the values of the cells was approximately bimodal, so we maintained this variable in its categorical form. Ruggedness2 was included since topography was consistently important to nesting peregrines in previous studies and we wanted to test if peregrines responded non-linearly to extremes in ruggedness in the study area. We also applied two logarithmic transformations to Elevation and Ruggedness (Ruggedness2 was the square of log-transformed Ruggedness) to reduce the influence of extremely high values on the model results (Figure A2). To aid in the comparison of coefficients and the interpretation of first and second order polynomials within the same model, we centred all variables on the mean and standardized by the standard deviation (Schielzeth 2010). Variables were tested pair-wise for collinearity with a Pearson’s correlation coefficient and for multicollinearity with Variance Inflation Factor (VIF; Zuur et al. 2010) using the R package Car (Weisberg and Weisberg, 2011) in R version 3.1.2 (R Core Team 2014). Following Dormann et al. (2007), we left out variables that had a Pearson’s correlation coefficient of over 0.70 or a VIF >10. According to our criteria, the variables used in the analysis were not collinear: the largest pairwise Pearson’s correlation coefficient was 0.60 between Cover and Temperature (see Figure A3 for all pair-wise correlations) and the highest VIF score was 2.44 for Temperature. According to Dormann et al. (2013), this collinearity should not be high enough to cause type II errors.
1.5.5 Data analysis
We estimated peregrine habitat selection using resource selection functions (RSFs; generalized linear models with a binomial family and logit link). Competing models were divided into groups of variables representing the influence of climate (Temperature and Precipitation), topography (Ruggedness, Ruggedness2, and Elevation), and prey availability (Cover), as well as one model potentially combining a number of environmental effects: the proximity to the coast (Coastal). This partitioning allowed us to compare the relative