Couts et bénéfices de l’association post-sevrage chez la chèvre de montagne (Oreamnos americanus) Mémoire Karina Charest Maitrise en biologie Maitre ès sciences (M.Sc.) Québec, Canada © Karina Charest, 2016
RÉSUMÉ
Les couts et bénéfices des soins parentaux ont été largement démontrés durant la période de lactation et croissance des jeunes, mais rarement après le sevrage. Pour nous permettre de mieux comprendre l’effet de ces soins à plus long terme, nous avons étudié les associations post-sevrage chez une population de chèvres de montagne (Oreamnos americanus) suivie depuis plus de 25 ans. Nous avons évalué l’effet des caractéristiques maternelles et de la taille de population sur l’occurrence et la fréquence des associations post-sevrage et avons étudié les couts potentiels de la reproduction et les bénéfices pour la mère et le juvénile âgé de 1 à 3 ans. Les femelles non reproductrices s’associent plus souvent avec les juvéniles d’un an que les mères reproductrices qui perdent leur chevreau et les mères lactantes, tandis que l’occurrence des associations avec un juvénile de 2 ans est plus élevée pour les femelles que pour les mâles. Un indice d’association élevé avec un juvénile de 2 ans augmente la probabilité qu’une mère ne se reproduise pas l’année suivante. Nous n’avons pas détecté de bénéfices des associations sur la croissance à court terme et la survie des juvéniles associés. Les associations post-sevrage induisent des couts reproductifs pour la mère, qui augmentent avec l’âge du juvénile.
ABSTRACT
The costs and benefits of parental care have been largely demonstrated during the lactation and offspring growth periods, but rarely after weaning. To better understand the effects of post-weaning parental care, we examined post-weaning associations in a population of mountain goats (Oreamnos americanus) monitored for > 25 years. We assessed the effect of maternal characteristics and population size on the occurrence and frequency of post-weaning associations and investigated potential reproductive costs and benefits for mothers and juveniles aged 1 to 3 years old. Barren mothers associated more frequently with yearlings than summer yeld mothers, and lactating mothers, whereas the occurrence of associations was higher for 2-year females than 2-year males. A higher association index with 2-year-olds increased the probability that a mother would be barren the following year. We did not detect benefits of associations on the short-term growth and survival of associated juveniles. Post-weaning associations induced reproductive costs for mothers, which exacerbated as associated juveniles aged.
TABLE DES MATIÈRES
RÉSUMÉ ... III ABSTRACT ... V TABLE DES MATIÈRES ... VII LISTE DES TABLEAUX ... IX LISTE DES FIGURES ... XI REMERCIEMENTS ... XIII AVANT-PROPOS ... XV
INTRODUCTION ... 1
Facteurs favorisant les associations post-sevrage ... 2
Couts potentiels de l’association post-sevrage pour la mère ... 7
Bénéfices de l’association post-sevrage pour le juvénile ... 7
Problématique ... 8
Objectifs ... 8
Population de chèvres de montagne à l’étude ... 9
Méthodologie ... 10 CHAPITRE PRINCIPAL ... 13 RÉSUMÉ... 15 ABSTRACT ... 17 Introduction ... 19 Methods ... 22 Study area ... 22 Captures ... 22 Post-weaning associations ... 23 Kid survival ... 24 Reproductive status ... 24 Social rank ... 24 Body mass ... 25 Statistical analyses ... 25 Results ... 28
Factors favoring post-weaning associations ... 29
Costs of post-weaning associations for mothers ... 30
Benefits of post-weaning associations for juveniles ... 31
Discussion ... 33
Factors favoring post-weaning associations ... 33
Costs of post-weaning associations for mothers ... 35
Benefits of post-weaning associations for juveniles ... 37
Acknowledgments ... 38
References ... 40
CONCLUSION GÉNÉRALE ... 49
BIBLIOGRAPHIE ... 55
LISTE DES TABLEAUX
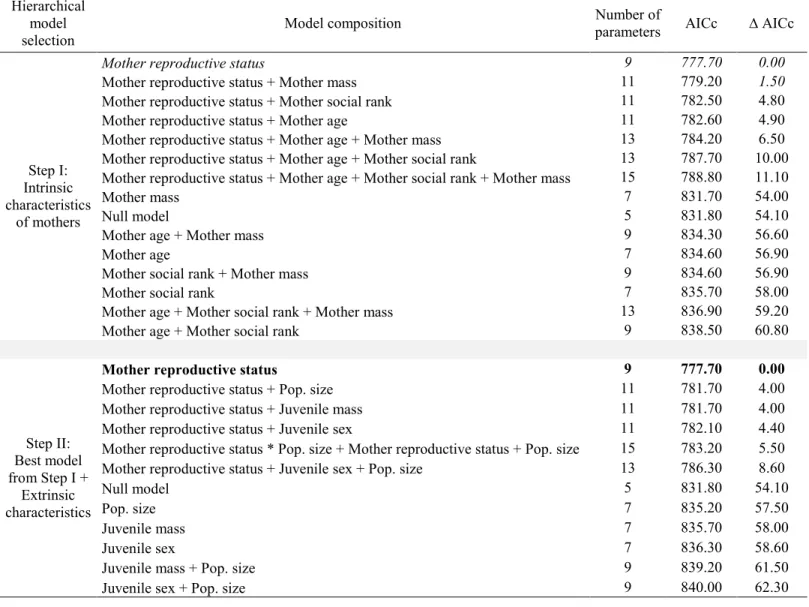
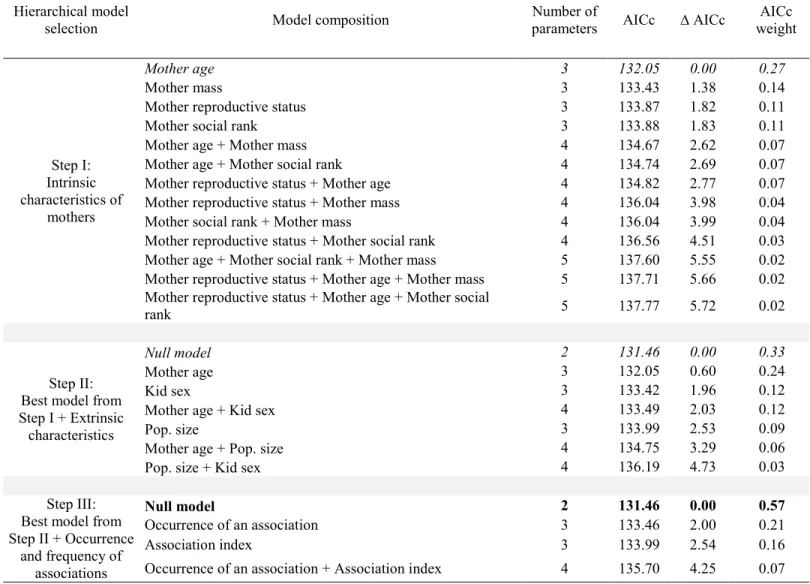
Table 1. Relative support received by hierarchical a priori zero-inflated negative binomial models explaining the occurrence and frequency (measured using an association index) of post-weaning associations of mothers with their yearling (n = 103) in a population of mountain goats (Oreamnos americanus) at Caw Ridge, Alberta, Canada (1991-2012). ... 63 Table 2. Relative support received by hierarchical a priori zero inflated negative binomial models explaining the occurrence and frequency (measured using an association index) of post-weaning associations of a mother with a 2-years-old (n = 72) in a population of mountain goats (Oreamnos americanus) at Caw Ridge, Alberta, Canada (1991-2012). ... 64 Table 3. Relative support received by hierarchical a priori generalized linear mixed models assessing the influence of associations with a yearling (n = 89) on the probability that a mother was barren the following year in a population of mountain goats (Oreamnos
americanus) at Caw Ridge, Alberta, Canada (1991-2012). ... 65
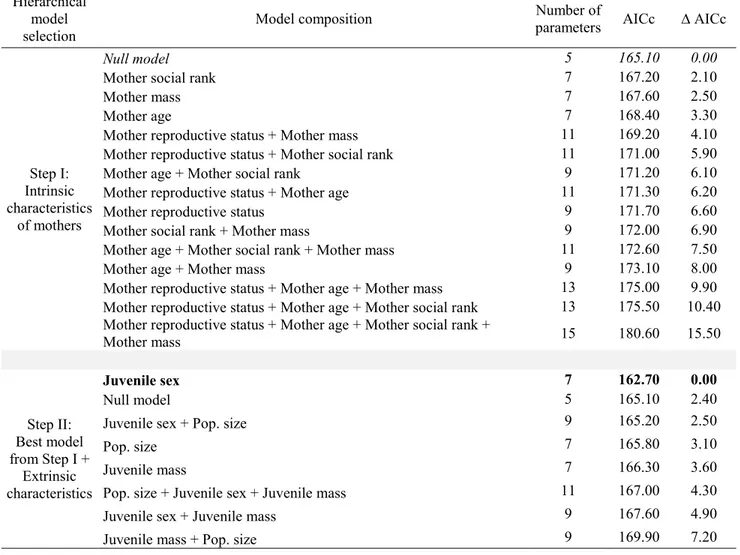
Table 4. Relative support received by hierarchical a priori generalized linear mixed models assessing the influence of associations with a 2-years-old (n = 64) on the probability that a mother was barren the following year in a population of mountain goats (Oreamnos
americanus), at Caw Ridge, Alberta, Canada (1991-2012). ... 67
Table 5. Relative support received by hierarchical a priori linear models assessing the influence of associations with a yearling (n = 27) on the age-specific body mass of kids in a population of mountain goats (Oreamnos americanus) at Caw Ridge, Alberta, Canada (1991-2012). ... 68 Table 6. Relative support received by hierarchical a priori linear models assessing the influence of associations with a 2-years-old (n = 21) on the age-specific body mass of kids in a population of mountain goats (Oreamnos americanus) at Caw Ridge, Alberta, Canada (1991-2012). ... 69 Table 7. Relative support received by hierarchical a priori generalized linear mixed models assessing the influence of associations with a yearling (n = 102) on the probability of yearling survival in the following year in a population of mountain goats (Oreamnos americanus) at Caw Ridge, Alberta, Canada (1991-2012). ... 70 Table 8. Relative support received by hierarchical a priori generalized linear mixed models assessing the influence of associations with a 2-years-old (n = 72) on the probability of the 2-years-old survival in the following year in a population of mountain goats (Oreamnos
americanus) at Caw Ridge, Alberta, Canada (1991-2012). ... 71
Table 9. Relative support received by hierarchical a priori linear mixed models assessing the influence of associations with a yearling (n = 101) on its age-specific body mass in a population of mountain goats (Oreamnos americanus) at Caw Ridge, Alberta, Canada (1991-2012). ... 73 Table 10. Relative support received by hierarchical a priori linear mixed models assessing the influence of associations with a 2-years-old (n = 71) on its age-specific body mass in a population of mountain goats (Oreamnos americanus) at Caw Ridge, Alberta, Canada (1991-2012). ... 75
LISTE DES FIGURES
Figure 1. Association post-sevrage entre une mère et un juvénile à Caw Ridge en juillet 2013 : mère # 368 avec son juvénile mâle âgé d’un an # 432. Photo par K. Charest Castro. ... 9 Figure 2. Relationship between the annual maternal reproductive status and the association index in a population of mountain goats (Oreamnos americanus) at Caw Ridge, Alberta, Canada (1991-2012). Post-weaning associations occurred between a mother and its (A) yearling(s) or (B) 2-year-old(s). We measured the association index by dividing the number of associations with the number of times the mother-juvenile dyad was observed during summer. Boxplots show the median (centerline), the 1st (lower box) and 3rd quartiles (upper box), and the data points within the range quartile ± 1.5* (interquartile range, shown using whiskers). Empty circles represent outliers. Letters contrast significant differences (P < 0.05). The relationship was significant and retained in the best model for yearlings only. ... 33
REMERCIEMENTS
Avant tout, j’aimerais d’abord remercier deux professeurs durant mon baccalauréat, soit le Dr. William Wickery pour m’avoir acceptée et supportée pour mon premier travail de recherche, précurseur de maitrise, et le Dr. Éric Lucas pour m’avoir aussi encouragée à poursuivre mes études au niveau supérieur, volontairement ou involontairement, par nos nombreuses discussions entre autres autour d’un feu lors des sorties terrain ! Merci de m’avoir fait confiance à ce moment, de croire que j’étais une bonne candidate pour la maitrise et de toujours m’avoir encouragée à poursuivre, persévérer et croire que j’y parviendrais. Également, sitôt mon baccalauréat complété, je souhaite grandement te remercier, Dr. Steeve Côté, pour m’avoir généreusement ouvert les portes de ton laboratoire suite à une rapide rencontre au congrès de la SQÉBC après avoir discuté de chiens et de Bella bien sûr ! Haha ! Merci de m’avoir fait confiance si rapidement et de m’avoir permis de compléter ce rêve d’études de maitrise dont je parlais déjà tant au début du baccalauréat. Merci de m’avoir proposé les chèvres de montagne pour lesquelles j’ai dû avouer ne rien connaitre au départ et de m’avoir aussi laissé choisir mon sujet, à travers lequel je me voyais bien : la mère et les juvéniles, quoi de mieux ! Merci enfin pour tes nombreuses histoires de terrain qui font de Caw Ridge un endroit tout aussi unique, mémorable où tu as accepté de m’envoyer après avoir tant tenté de m’en dissuader lors de notre première entrevue !
Un merci particulier à Joëlle Taillon et Julien Mainguy qui m’ont donné d’excellents commentaires quant à l’avancement de mon projet à son début. Malgré que je fusse nouvelle au laboratoire, merci Joëlle pour ton temps sans hésiter et tes commentaires précieux sur la façon de rédiger, j’ai beaucoup appris en te lisant.
Merci particulièrement à Jean Marchal pour ton aide à la création de mon indice d’association, qui dura plusieurs semaines de problèmes, sans ton aide, mon départ de projet aurait été des plus tumultueux dans R Studio, alors que je n’y comprenais rien.
Merci à Sonia De Bellefeuille pour ton sourire de tous les jours, ton support et tes corrections en français, bien que discrète, tu es toujours là pour nous aider et ô combien efficace !
Merci à Caroline Hins pour ton aide toi aussi sur la rédaction en français et tes nombreux bons commentaires pour notre lutte de bien écrire. Merci pour ton support moral et tous tes encouragements dans les moments moins évidents.
Merci à mes nombreux collègues, autres étudiants, qui m’ont aidé à parcourir ce long chemin qu’est la maitrise en recherche. Merci aux « chèvres » : Patrick Morissette, Julien Hénault-Richard; aux « caribou » Alexandre Rasiulis, Glenn Yannic, Mael Le Corre pour ton aide précieuse en codage statistique et toutes ces soirées seuls au labo à jaser, Sabrina Plante pour les nombreuses discussions en statistiques où tu m’as plus que souvent éclairée dans mon brouillard et aussi ta patience pour tous mes petits dérangements, et Mathieu Leblond merci infiniment pour toute ta patience, volonté et ô combien enrichissantes corrections; merci aux « chevreux » Michael Bonin, Amélie Drolet bien qu’une licorne, et Nicolas Houde; merci à la « plante » Émilie Champagne pour ton aide sur les présentations orales, et merci aux tous « nouveaux » incluant Andrea Panagaski pour ta correction sur mon anglais. Le « laboratoire Côté » ne serait rien sans cette merveilleuse équipe que vous formez tous et dont j’ai été si choyée de pouvoir côtoyer. Vous êtes plus que des collègues, mais êtes devenus de bons amis. Merci.
Finalement, d’une manière plus personnelle, merci à mon papou Alain Charest et ma mouma Caroline Larouche, ma marraine Katie Charest et Marquis Bradette pour vos encouragements sans relâche et votre support moral. Votre foi inébranlable en moi m’a donné tout l’espoir pour me rendre à mes fins dans ce projet entouré de mes nombreuses activités extra-universitaires. Sans votre écoute et votre présence, mon parcours aurait été beaucoup plus ardu moralement. Merci à vous, je vous aime.
« […] quand tu veux quelque chose, tout l’Univers conspire à te permettre de réaliser ton
AVANT-PROPOS
Ma maitrise est présentée avec l’insertion d’un article scientifique rédigé en anglais dans le but de le publier dans une revue scientifique internationale. Je suis la principale auteure de ce mémoire et de cet article et j’étais donc responsable du développement des hypothèses et prédictions, de la compilation et l’analyse des données et enfin, de la rédaction. Mon directeur de maitrise, Steeve D. Côté professeur au département de biologie de l’université Laval sera coauteur de l’article.
Introduction
Plusieurs écologistes s’intéressent à l’investissement parental à cause de son influence marquée sur le succès reproducteur des individus (Trivers, 1972; Clutton-Brock, 1984). L’investissement parental peut se définir comme toute caractéristique ou action parentale augmentant la valeur adaptative (fitness) de la progéniture. Cet investissement entraine un cout sur le succès reproducteur futur, la survie ou la fécondité du parent, et sa valeur reproductive résiduelle (c.-à-d. les opportunités de reproduction future) s’en trouve donc diminuée. Il est possible d’évaluer l’investissement parental en mesurant, par exemple, la masse corporelle perdue par la mère après une période de lactation et l’effet sur sa survie, ou le taux de croissance des juvéniles en lien avec leur survie (Clutton-Brock, 1991). L’intervalle de temps entre les naissances est aussi un indice de l’investissement parental (Lee and Moss, 1986; Hewison and Gaillard, 1999; Bercovitch et al., 2000). Chez le mâle, on peut aussi observer une probabilité d’acquisition réduite d’un autre partenaire sexuel (Schwagmeyer et al., 2012).
Le fait de prodiguer des soins parentaux, comme préparer un nid, creuser une tanière, toiletter, réchauffer, allaiter ou protéger les jeunes contre les prédateurs n’influence pas nécessairement la capacité des parents à allouer des ressources pour la production d’un autre jeune (Oftedal, 1985; Clutton-Brock, 1991). Par définition, le soin parental représente toute forme de comportement parental qui vise à assurer la croissance et la survie du jeune, sans réduire la valeur adaptative subséquente du parent (Clutton-Brock, 1991). On parle d’investissement parental quand les soins parentaux ont un cout en termes de valeur adaptative. Les soins parentaux varient grandement selon les espèces : ils sont absents chez la majorité des invertébrés et des poissons, alors que plusieurs espèces d’oiseaux et de mammifères prodiguent des soins variés (Clutton-Brock, 1991). Cette variabilité dépend de divers facteurs tels que le moment de la fertilisation des œufs, la dépendance des juvéniles aux soins, les conditions environnementales ou encore les composantes biodémographiques individuelles (Clutton-Brock, 1991). Par conséquent, ces différents facteurs influenceront la masse, la croissance et la survie des jeunes (Bernardo, 1996). De plus, les soins parentaux
peuvent aussi varier selon le système d’accouplement (Davies et al., 2012). Chez la majorité des espèces polygynes, puisque les mâles adultes sont ségrégés spatialement, ils investissent plutôt dans l’effort reproducteur (Oliveras and Novak, 1986; Patris and Baudoin, 2000). Les soins aux jeunes sont alors principalement prodigués par la femelle. Les liens qui se créent alors entre la mère et sa progéniture sont déterminés par l’atteinte d’un équilibre entre les besoins de la mère et ceux de son(ses) jeune(s). Lorsque cet équilibre est débalancé, le sevrage du jeune survient (Trivers, 1974). Lorsque la mère continue d’offrir des soins parentaux aux jeunes après le sevrage, on parle alors d’association post-sevrage entre la mère et son juvénile, comme les associations post-sevrage observées, par exemple, chez les bisons (Bison bison) (Green et al., 1989; Brookshier and Fairbanks, 2003), les hyènes tachetées (Crocuta crocuta) (Holekamp et al., 1997) et les sangliers (Sus scrofa) (Kaminski et al., 2005). Ces associations peuvent se créer lorsque les bénéfices pour le(s) juvénile(s) sont élevés et que les couts pour la mère demeurent faibles (Green et al., 1989).
Facteurs favorisant les associations post-sevrage
La qualité et la quantité des soins offerts après le sevrage peuvent varier selon différentes caractéristiques individuelles maternelles (Green, 1986; Bernardo, 1996) ou selon les conditions environnementales (Hauser and Fairbanks, 1988; Therrien et al., 2007). La formation d’associations post-sevrage peut donc être favorisée ou non selon ces caractéristiques et conditions.
Chez les mammifères, la lactation est considérée comme l’un des soins maternels les plus directs que la mère puisse offrir à son jeune. Toutefois, la lactation engendre des couts énergétiques élevés, plus que la gestation elle-même (Oftedal, 1985; Clutton-Brock et al., 1989; Tucker, 1994). Ces couts peuvent engendrer, entre autres, une diminution de la masse corporelle et des réserves lipidiques de la femelle (Simard et al., 2010) et peuvent parfois se traduire par une pause reproductive chez les jeunes femelles lorsque la densité de population est élevée (Hamel et al., 2010). Ainsi, les femelles adultes peuvent être séparées en trois
catégories de statut reproducteur, des couts de reproduction les plus faibles vers les plus élevés (adapté de L’Heureux et al., 1995) : (1) femelle non reproductrice, (2) mère reproductrice dont le jeune de l’année meurt avant le sevrage (mortalité juvénile), et (3) mère lactante ayant produit un jeune qui survit au moins jusqu’au sevrage. Ces différentes catégories sont associées à des couts énergétiques distincts reliés à la reproduction : la mère non reproductrice n’en aura aucun ; certaines auront les couts associés à la gestation en plus de ceux de la lactation, soit partielle ou complète.
Chez les grands ongulés, les femelles non reproductrices sont plus souvent observées en association post-sevrage avec leurs jeunes que les mères lactantes, entre autres chez les bisons (Green et al., 1989), les cerfs élaphes (Cervus elaphus) (Guinness et al., 1979) et les moutons domestiques (Ovis aries) (Lawrence, 1990; Rowell, 1991). De façon similaire, les femelles mouflon d’Amérique (O. canadensis) ayant perdu leur agneau à la naissance ont tendance à s’associer plus souvent avec le jeune de l’année précédente que les femelles non reproductrices. Ceci pourrait être dû au fait que le lait demeure disponible suite à la mort de l’agneau (L’Heureux et al., 1995). Ainsi, le statut reproducteur de la femelle favorise ou non l’association avec son(ses) jeune(s) des années précédentes selon les couts énergétiques engendrés.
Le statut reproducteur de la mère influence les soins maternels prodigués, le développement des jeunes et la formation d’associations post-sevrage, mais ces soins et associations peuvent aussi être modulés par l’âge de la mère. Contrairement aux femelles âgées, les jeunes femelles doivent faire face à différents compromis en ce qui a trait à l’allocation de leur énergie (Côté and Festa-Bianchet, 2001b; Hamel and Côté, 2009). Les jeunes femelles doivent allouer leur énergie à la croissance, la survie, la lactation et le succès reproducteur futur, tandis que les femelles plus âgées, ayant atteint la masse corporelle asymptotique, n’ont pas à investir dans la croissance (Bérubé et al., 1999). Par conséquent, la qualité des soins maternels prodigués peut être plus faible pour les jeunes mères, tel qu’observé chez le
mouflon d’Amérique, où elles présentent des périodes d’allaitement plus courtes et effectuent moins de toilettage de leur jeune que les mères plus âgées (Festa-Bianchet, 1988). En ce qui concerne les effets de l’âge sur les associations post-sevrage, chez les bisons, par exemple, ce sont les plus jeunes mères qui entretiennent des liens plus étroits par contacts physiques fréquents avec leurs juvéniles (Green, 1993). Elles peuvent ainsi compenser leur manque d’expérience maternelle à prodiguer des soins (Kaminski et al., 2005) et pourraient atténuer certains désavantages physiques ou sociaux (Green, 1993). Contrairement aux bisons, l’âge maternel chez le mouflon d’Amérique n’influence pas la probabilité d’association post-sevrage entre la mère et les juvéniles (L’Heureux et al., 1995). Il semble donc que l’âge maternel puisse, chez certaines espèces, avoir une influence sur la formation d’association post-sevrage alors que chez d’autres, non.
Un rang de dominance élevé peut s’avérer être une caractéristique maternelle déterminante dans les relations mères-juvéniles. Ainsi, la mère pourrait augmenter le taux de survie post-sevrage de ses jeunes bénéficiant d’un accès prioritaire aux ressources (Watts et al., 2009). Selon Holekamp et al. (1997), le rang social influence fortement l’association entre mères et juvéniles. Chez les hyènes tachetées, par exemple, les mères de rang social supérieur ont un indice d’association plus élevé avec leurs jeunes sevrés que les mères de rang social faible, permettant ainsi de conserver la stabilité de la dominance dans la hiérarchie sociale des femelles. En effet, les femelles apparentées regroupées peuvent ainsi gagner plus facilement les interactions agonistiques avec les femelles non apparentées de rang social plus faible (Holekamp et al., 1997). Chez plusieurs espèces d’ongulés, le rang social est fortement corrélé à des caractéristiques maternelles comme l’âge [bison (Rutberg, 1986), cerf élaphe (Clutton-Brock et al., 1984), mouflon d’Amérique (Festa-Bianchet, 1991), chèvre de montagne (Oreamnos americanus) (Côté, 2000)] ou la masse corporelle [cerf élaphe (Clutton-Brock et al., 1984)]. Par exemple, chez les espèces où le rang social est corrélé positivement à l’âge, les plus jeunes mères peuvent avoir tendance à s’associer avec leurs juvéniles plus souvent que les mères plus âgées afin de compenser pour leur faible statut social lors des interactions agonistiques, comme démontré pour les mères chez le singe vert
(Cercopithecus aethiops sabaeus) (Fairbanks and McGuire, 1985) et le cerf élaphe (Clutton-Brock et al., 1982).
Bien que l’influence de la masse maternelle sur la formation d’associations post-sevrage ait été peu étudiée chez les ongulés, L’Heureux et al. (1995) ont néanmoins étudié l’effet de l’association sur le gain de masse de la mère associée chez le mouflon d’Amérique sans toutefois observer de diminution de gain de masse. La masse maternelle est toutefois reconnue comme un facteur critique pour la survie du jeune (Clutton-Brock 1991, Festa-Bianchet et al. 1998), particulièrement à la mise bas ainsi qu’au sevrage (Cameron et al. 1993; Taillon et al. 2012). En déterminant la masse du jeune, la masse maternelle est indirectement reliée à la survie estivale et hivernale qui dépendent généralement de la masse chez les jeunes (Théoret-Gosselin et al., 2015). De plus, une mère ayant une masse corporelle élevée prendrait plus de risques pour défendre ses jeunes qu’une mère plus petite, comme observé chez le cerf de Virginie (Odocoileus virginianus) (Smith, 1987).
Chez certaines espèces d’ongulés polygynes qui présentent un fort dimorphisme sexuel, le sexe du jeune est un facteur déterminant dans l’allocation de l’énergie pour les soins parentaux (p. ex. cerf élaphe, mouflon d’Amérique), alors qu’il ne l’est pas chez d’autres (p. ex. bison d’Amérique, caribou [Rangifer tarandus]) (Hewison and Gaillard, 1999). Puisque le succès reproducteur des mâles ongulés dépend généralement de leur habileté à combattre ainsi que de leur taille corporelle, leur succès reproducteur est fortement influencé par la quantité et la qualité des soins parentaux lors de la croissance. Les mères devraient alors allouer plus d’énergie dans la croissance de leurs jeunes mâles que dans celle de leurs jeunes femelles (Trivers and Willard, 1973; Bercovitch et al., 2000). La mère peut ajuster le nombre de périodes d’allaitement selon le sexe du jeune en accordant en moyenne plus de périodes d’allaitement à ses jeunes mâles qu’à ses jeunes femelles, comme c’est le cas chez le mouflon à manchettes (Ammotragus lervia) (Cassinello, 1996). En plus d’influencer les soins aux jeunes, le sexe peut aussi influencer l’association post-sevrage. Par exemple, chez le mouflon
d’Amérique, les femelles d’un an s’associent plus souvent à leur mère que les mâles du même âge (L’Heureux et al., 1995). Chez les cerfs élaphes, les mâles d’un an sont moins associés à leur mère que les femelles, possiblement parce que la mère dirige plus d’agressions envers eux qu’envers leurs filles ou parce qu’ils sont plus explorateurs que les jeunes femelles (Guinness et al., 1979). Bien que l’effet semble variable selon les espèces, le sexe du jeune pourrait moduler la quantité et la qualité des soins maternels prodigués, ce qui peut se répercuter directement sur la croissance, la survie et le succès reproducteur de ce jeune.
En plus des caractéristiques individuelles maternelles, la densité de population peut aussi influencer la qualité et quantité des soins parentaux. De ce fait, les effets d’une densité de population élevée sont bien connus chez les mammifères, pouvant entrainer par exemple une masse plus faible à la naissance (Kjellander et al., 2006), un délai dans la croissance (Mobaek et al., 2013), une diminution de la masse corporelle des jeunes avant le sevrage (Gaillard et al. 1996) ou suite au sevrage (Hewison et al., 2002), et compromettre les chances de survie des juvéniles (Côté and Festa-Bianchet, 2001a; Coulson et al., 2001; Couturier et al., 2009; Hamel et al., 2010). Les effets dépendants de la densité peuvent influencer les juvéniles jusqu’à l’âge adulte en affectant leur condition corporelle (Pettorelli et al., 2002; Bonenfant et al., 2009; Hamel et al., 2010), en entrainant un délai dans l'âge de primiparité (Langvatn et al., 1996; Martin and Festa-Bianchet, 2012) ou en réduisant le succès reproducteur (Forchhammer et al., 2001). La probabilité de formation d’associations post-sevrage peut ainsi être influencée par la densité de population à travers une multitude de traits biodémographiques. À faible densité de population, les associations post-sevrage entre mère et juvéniles étaient plus rares chez le mouflon d’Amérique tandis qu’à forte densité, les mères ayant perdu leur agneau peu après la naissance s’associaient plus fortement avec leur jeune de l’année précédente (L’Heureux et al., 1995). Cette tactique atténue possiblement les effets négatifs d’une forte densité de population en augmentant le fitness des jeunes sevrés. Chez le cerf élaphe, la densité de population était toutefois inversement liée aux associations post-sevrage, en réponse à la compétition directe pour les ressources (Albon et al., 1992).
Couts potentiels de l’association post-sevrage pour la mère
Bien que les couts de la reproduction courante soient bien documentés pour la mère (Hamel et al., 2009), les couts d’une association post-sevrage avec ses juvéniles sont moins connus. L’Heureux et al. (1995) n’ont pas détecté d’effet de l’association mère-juvénile sur la masse corporelle de la mère chez le mouflon d’Amérique, mais Johnson (1986) a démontré que le succès reproducteur de la mère wallaby à cou rouge (Macropus rufogriseus banksianus) diminuait avec le temps passé avec ses juvéniles. De plus, peu d’études ont considéré les couts de l’association post-sevrage pour la mère chez les rongeurs (Moses and Millar, 1994) et les oiseaux (Earnst and Bart, 1991).
Bénéfices de l’association post-sevrage pour le juvénile
En s’associant avec sa mère après le sevrage, le juvénile peut obtenir plusieurs bénéfices. Un juvénile associé bénéficie d’une plus grande probabilité de survie par rapport à un juvénile non associé, comme observé chez l’orignal de l’Alaska (Alces alces gigas) (White et al. 2001). De plus, un juvénile qui s’associe à sa mère après le sevrage peut acquérir un rang social élevé et croitre plus rapidement qu’un juvénile non-associé (Holekamp et al., 1997). Un juvénile associé se fait moins souvent déplacer par les autres membres du groupe et obtient de meilleures positions dans le groupe (p. ex. position centrale plutôt que périphérique) qu’un non-associé, lui conférant un meilleur accès aux sources de nourriture (Green et al., 1989). Une position centrale peut aussi diminuer le risque de prédation (Green et al., 1989; Côté et al., 1997; Brookshier and Fairbanks, 2003). Enfin, l’association post-sevrage peut favoriser l’apprentissage de la localisation des ressources dans l’espace, tel que la localisation des sites d’alimentation préférentiels et des trajets de déplacements saisonniers (Lawrence, 1990).
Problématique
La majorité des études réalisées à ce jour sur les soins parentaux ont été faites durant la période de lactation. Très peu d’études se sont attardées aux associations post-sevrage. De plus, les études sur les associations post-sevrage considèrent rarement l’influence des caractéristiques maternelles sur la formation de ces associations, ainsi que les effets de l’association elle-même sur la capacité des mères à se reproduire et sur les caractéristiques du jeune produit. La chèvre de montagne est une espèce chez qui les associations post-sevrage entre la mère et son juvénile sont fréquemment observées. Ongulé des milieux alpins, la chèvre de montagne est une espèce longévive et polygyne. Dans les populations naturelles, les femelles ne se reproduisent pour la première fois qu’à environ 4 ans et n’ont habituellement qu’un seul chevreau annuellement. La quantité et la qualité des soins maternels sont alors déterminantes pour la survie du jeune (Théoret-Gosselin et al., 2015). Puisqu’il y a ségrégation des sexes chez les adultes et que les chevreaux et les juvéniles (≥ 1 an jusqu’à 3 ans) restent dans le groupe maternel, la mère s’avère être le seul parent à offrir les soins parentaux durant la période où la majorité de la croissance a lieu. Plusieurs études sur la chèvre de montagne ont traité des soins maternels envers les jeunes de l’année (Côté and Festa-Bianchet, 2001a; Gendreau et al., 2005; Théoret-Gosselin et al., 2015) mais aucune de ces études ne s’est attardée aux soins post-sevrage.
Objectifs
Cette étude a pour but de documenter les effets à court et long terme des soins maternels post-sevrage sur la capacité reproductive de la mère chez la chèvre de montagne, ainsi que sur les caractéristiques phénotypiques de son juvénile. Nous voulons identifier les facteurs qui déterminent la formation et l’abondance d’associations post-sevrage ainsi que l’impact de ces associations sur les individus impliqués afin d’améliorer nos connaissances sur l’écologie des soins maternels post-sevrage chez une population d’ongulés. Avec cette étude, nous pourrons, entre autres, mieux comprendre si la continuité post-sevrage des soins maternels est une forme de soin ou plutôt un investissement par la mère. À l’aide de données issues d’un suivi à long terme d’une population naturelle de chèvres de montagne marquées,
trois questions sont abordées pour mieux comprendre l’occurrence des associations post-sevrage : (1) De quelle manière les caractéristiques individuelles maternelles et la densité de population favorisent ou non la formation des associations post-sevrage ? (2) Y a-t-il des couts en termes de fitness pour la mère à prodiguer des soins post-sevrage à ses juvéniles ? et (3) Quels sont, chez le juvénile, les bénéfices de la formation d’associations post-sevrage ?
Figure 1. Association post-sevrage entre une mère et un juvénile à Caw Ridge en juillet 2013 : mère # 368 avec son juvénile mâle âgé d’un an # 432. Photo par K. Charest Castro.
Population de chèvres de montagne à l’étude
La population de chèvres de montagne à l’étude habite un complexe montagneux nommé Caw Ridge se trouvant du côté est des montagnes Rocheuses en Alberta (Canada). Le site d’étude de Caw Ridge est constitué majoritairement de quatre montagnes reliées par des sentiers créés par les chèvres au fil des années. Au sommet de ces montagnes, plusieurs pentes rocailleuses et falaises servent de terrains de fuite afin d’échapper aux prédateurs (p. ex. l’ours grizzly, Ursus arctos, le loup, Canis lupus, et le cougar, Puma concolor) (Festa-Bianchet and Côté, 2008). Les conditions climatiques sont typiques des milieux alpins, c’est-à-dire ardues, avec une saison estivale courte et fraiche et une saison hivernale longue, froide
et aride avec de forts vents. Des précipitations sous forme de neige ont lieu durant toute l’année. La disponibilité de ressources alimentaires de qualité s’étend donc sur une brève période de l’année en raison d’une saison de croissance courte. La population de chèvres de montagne utilise une superficie d’environ 28 km2 de cet habitat, entre 1,750 et 2,185 m d’altitude.
Méthodologie
Le suivi de la population de chèvres de montagne de Caw Ridge est réalisé depuis 1989 et est basé sur la capture et le marquage d’individus d’âge connu (Festa-Bianchet and Côté, 2008). Depuis 1993, 98% de la population est marquée. Les chèvres sont capturées à l’aide de trappes Stephenson en bois dans lesquelles un bloc de sel est déposé afin de les attirer. Les captures se font à partir de la fin mai jusqu’à la mi-septembre, mais particulièrement en juin. Les juvéniles d’un et deux ans sont capturés et marqués individuellement à l’aide d’une combinaison d’étiquettes de plastique colorées apposées aux oreilles. Les femelles de deux ans et plus sont marquées avec des colliers visuels pour faciliter l’observation à distance. Tout au long de l’été, la masse corporelle est prise à l’aide de balances électroniques (50 x 130 cm) (± 0,5 kg) lorsque les individus visitent le site de capture, sans avoir à manipuler les individus (Bassano et al., 2003). Afin de confirmer les assignations parentales des jeunes âgés d’un an, un échantillon de peau de chaque individu est recueilli et des analyses génétiques sont effectuées (Mainguy et al., 2009). La taille de population est évaluée annuellement comme étant le nombre total d’individus dans la population.
La récolte de données comportementales servant à décrire les associations mère-juvénile se fait en observant quotidiennement les groupes de femelles à distance à l’aide de jumelles (10 x) ou de télescopes (15–45 x) du 15 mai environ au 15 septembre. L’emplacement du groupe ainsi que la taille du groupe sont notés. Le statut reproducteur des mères est assigné selon 3 catégories distinctes : (1) mère en lactation, femelle dont le chevreau survit jusqu’au 15 septembre, (2) mère reproductrice dont le chevreau meurt avant le sevrage et (3) mère en
pause reproductive. Le sexe des chevreaux est déterminé par l’observation de la région anogénitale lors des périodes d’allaitement ou selon la position qu’il prend lorsqu’il urine (Côté et Festa-Bianchet 2001a). La survie hivernale est établie au 1er juin de chaque année et la survie estivale au 15 septembre. La date de sevrage considérée est le 15 septembre puisque les allaitements sont alors peu fréquents et courts et la survie des jeunes qui perdent leur mère au début de l’automne ou non est similaire (Côté et Festa-Bianchet 2001a).
Lors de l’observation de chaque groupe de femelles, les associations post-sevrage entre mères et juvéniles âgés entre un et trois ans sont identifiées comme étant une mère et son juvénile situés dans un rayon d’environ 1,5 m, à peu près équivalent à la longueur d’une chèvre adulte. Ces associations sont observées principalement lors des périodes de repos/rumination. À ces moments, l’association est claire, la mère ne tolère aucun autre juvénile que le sien à ses côtés. Une mère suivie de près par un de ses juvéniles lors d’un déplacement peut aussi représenter une association. Au total, 234 dyades juvéniles d’un an, 167 dyades mère-juvéniles de 2 ans et 113 dyades mère-mère-juvéniles de 3 ans ont été suivies entre 1991 et 2012.
Le rang social des femelles adultes est évalué en observant les interactions agonistiques entre elles de manière ad libitum (Altmann, 1974). L’identité de l’individu qui initie l’interaction et celle de l’individu qui la subit, en plus de l’issue de l’interaction (c.-à-d., celui qui la remporte) sont enregistrées. Le gagnant est déterminé lorsqu’un individu s’éloigne à plus de 1,5 mètres et démontre des comportements de soumission (Locati and Lovari, 1991). Le nombre d’interactions totales remportées et perdues par individu est calculé et permet ensuite de produire une matrice de dominance. La linéarité annuelle de la hiérarchie de dominance est calculée en utilisant l’indice h’ (De Vries, 1995), variant entre 0 (non linéaire) et 1 (parfaitement linéaire). Par la suite, une méthode itérative (10000 réitérations) (De Vries, 1995) est utilisée pour vérifier la significativité de la valeur h’. Finalement, un rang est attribué aux femelles par la méthode de De Vries (1998).
Afin de répondre à mes objectifs, j’ai étudié plusieurs variables maternelles telles que le statut reproducteur annuel, l’âge, le rang social et la masse de la mère. La taille de la population de chèvres a aussi été mise en relation avec la présence et le nombre d’associations post-sevrage. Afin de vérifier si l’association mère-juvénile engendre un cout pour la mère, j’ai étudié la présence d’une pause reproductive l’année suivant l’association ainsi que la masse du chevreau la même année de l’association avec un juvénile de 1 à 3 ans. Enfin, pour évaluer les bénéfices potentiels de l’association chez le juvénile, sa masse lors de l’année de l’association et sa survie à l’année suivante ont été analysées.
CHAPITRE PRINCIPAL
Reproductive costs of post-weaning associations for mountain goat mothers
Karina Charest Castro1,2 and Steeve D. Côté1,2
1. Département de biologie, Université Laval, Québec, Canada 2. Centre d’études nordiques, Québec, Canada
RÉSUMÉ
Les impacts des caractéristiques maternelles et des conditions environnementales sur la croissance et la survie ont été principalement étudiés tôt durant le développement des petits, mais rarement après le sevrage. Afin de mieux comprendre les effets des soins maternels suite au sevrage, nous avons évalué le rôle de ces soins prolongés dans une population de chèvres de montagne (Oreamnos americanus) qui est suivie depuis plus de 25 ans. Pour chaque dyade mère-juvénile (âgé de 1 à 3 ans), nous avons calculé la fréquence d’association. Nous avons examiné (1) l’influence des caractéristiques de la mère et de la taille de population sur la formation des associations post-sevrage entre mères et juvéniles, (2) les couts à court terme des associations sur le succès reproducteur de la mère, et (3) les bénéfices à court terme des associations sur les composantes biodémographiques des juvéniles. La formation d’associations post-sevrage avec les juvéniles d’un an dépend du statut reproducteur de la mère : les femelles non-reproductrices s’associent plus fréquemment avec les juvéniles d’un an que les mères reproductrices qui perdent leur chevreau et les mères reproductrices lactantes. La probabilité des mères à s’associer avec un juvénile âgé de 2 ans augmente lorsque le juvénile est une femelle. Les associations avec un juvénile de 2 ans augmentent aussi la probabilité que la mère soit en pause reproductive l’année suivante, ce qui suggère un cout reproductif des associations post-sevrage pour les mères. Les associations post-sevrage avec les juvéniles d’un an ou plus n’influencent pas la masse des chevreaux. Nous n’avons pas détecté de bénéfices des associations sur la croissance à court terme et la survie des juvéniles associés. Nous concluons que donner des soins à des juvéniles plus âgés est plus couteux pour les mères qu’offrir des soins à des juvéniles plus jeunes.
ABSTRACT
The impacts of maternal characteristics and environmental conditions on juvenile growth and survival have been mainly studied during early offspring development, but rarely after weaning. To better understand the effects of post-weaning parental care, we assessed the role of prolonged maternal care in a population of mountain goats (Oreamnos americanus) that has been monitored for > 25 years. For each mother and juvenile (aged 1 to 3 years) dyad, we calculated the frequency of association. We investigated (1) the influence of maternal characteristics and population size on the formation of post-weaning associations between mothers and juveniles, (2) the short-term costs of associations on maternal reproductive success, and (3) the short-term benefits of associations on juvenile life-history traits. The formation of post-weaning associations with yearlings depended on the maternal reproductive status: barren mothers associated more frequently with yearlings than summer yeld mothers and lactating mothers. The probability of mothers to associate with a 2-years-old juvenile was higher for daughters than sons. Associations with 2-year-2-years-olds also increased the probability that mothers would be barren the following year, suggesting a reproductive cost of post-weaning associations for mothers. Post-weaning associations with ≥ 1year-old juveniles did not influence the mass of newborn kids. We did not detect benefits of associations on the short-term growth and survival of associated juveniles. We conclude that caring for older weaned offspring is costlier for mothers than caring for younger ones.
K. Charest Castro & S. D. Côté
Département de biologie and Centre d’études nordiques, Université Laval, Québec, QC, G1V 0A6, Canada
Introduction
Kinship interactions may provide several advantages for gregarious mammals, including learning of traditional traveling routes and safe areas (Festa-Bianchet, 1991; Di Fiore and Suarez, 2007), accessing food resources and learning their distribution (Valsecchi et al., 1996; Ratcliffe and ter Hofstede, 2005), cooperating during agonistic interactions with conspecifics (Fairbanks and McGuire, 1985), alliance formation (Connor et al., 2001; Parsons et al., 2003), defense against predators (Ross, 2001; Wright et al., 2001; Graw and Manser, 2007), and caring for offspring (Clutton-Brock et al., 2001, 2002). In contrast, increased intraspecific competition may decrease breeding success and offspring survival rate as group size increases (Stewart et al., 2005; Richard et al., 2014). Consequently, individuals should tradeoff between group size benefits (e.g., lower scanning rate, and higher rates of predator detection and foraging) and costs (e.g., higher intraspecific competition for resources, higher rate of interactions) (see Elgar, 1989, and Roberts, 1996 for a review).
When resources are limited, life history theory predicts that adults should trade off energy allocation among growth, reproduction, and survival to maximize their lifetime reproductive success (Stearns, 1992). Conservative reproductive strategies often dominate in long-lived species, as fitness usually increases with longevity (Clutton-Brock, 1988; Festa-Bianchet and Côté, 2008). When resources are scarce, individuals may reduce their reproductive input or postpone reproduction to a subsequent year to favor their own survival (Cameron, 1994). Similarly, individuals may adjust energy allocation for offspring care depending on environmental conditions such as food quality (Hauser and Fairbanks, 1988), abundance of conspecifics (Therrien et al., 2007), and predation risk (Ghalambor et al., 2013). Parents should consequently adjust their level of offspring care depending on variations in benefits and costs for their offspring as well as themselves (Trivers, 1974).
In a variety of species, including birds (Covas and Griesser, 2007), mammals (Silk et al., 2009) and insects (Park and Choe, 2003), mothers may continue caring for their young after
the weaning period. This strategy was shown to increase offspring survival (Covas and Griesser, 2007; Silk et al., 2009) and growth (Park and Choe, 2003). Prolonged care after the weaning period (hereafter post-weaning association) has been observed in many ungulates [bison (Bison bison) (Green et al., 1989; Green, 1993; Brookshier and Fairbanks, 2003); wild boars (Sus scrofa) (Kaminski et al., 2005); bighorn sheep (Ovis canadensis) (L’Heureux et al., 1995)]. For the post-weaning association to persist, it should be beneficial for the juvenile and not too energetically costly for the mother (Trivers, 1974); otherwise, the association is expected to cease (Higashi and Yamamura, 1993).
Post-weaning associations mainly provide benefits for the associated juveniles. They may be less displaced by conspecifics during foraging and may show increased feeding efficiency (Green et al., 1989; Brookshier and Fairbanks, 2003). Associated juveniles may also obtain support during agonistic interactions (Jonker et al., 2012) or gain a higher social status (Holekamp et al., 1997) as well as higher survival rates (Moses and Millar, 1994). Additionally, they may have a more central position in the group (Green et al., 1989), leading to a higher protection against predators (Sherman, 1977).
The costs of parental care on the current and future reproduction are well documented (see Clutton-Brock (1991), and Kölliker (2012) for a review); the costs of post-weaning associations, however, are poorly studied. L’Heureux et al. (1995) found no effect of post-weaning association on the maternal body mass in bighorn sheep. Johnson (1986) showed that the reproductive success of red-necked wallaby (Macropus rufogriseus banksianus) mothers decreased as they spent more time with their weaned offspring. Moses and Millar (1994) did not find any cost for mothers wild bushy-tailed woodrats (Neotoma cinerea) in sharing their natal site with their yearling daughters.
To assess the influence of post-weaning associations in large mammals, we investigated the influence of maternal intrinsic characteristics (e.g., body mass, age) and extrinsic characteristics (e.g. population size) on the occurrence and frequency of post-weaning associations with juveniles 1 to 3 years of age. We also estimated several proxies of the potential costs of post-weaning associations for mothers (e.g. reproductive output), as well as the potential benefits of post-weaning associations for juveniles (e.g., body mass, survival). We addressed these questions using a capital breeder, the mountain goat (Oreamnos americanus), which has been studied at Caw Ridge, Alberta (Canada) for over 25 years. To our knowledge, no comparable long-term data have been used to assess the possible costs and benefits of post-weaning associations with juveniles up to 3 years old.
Because maternal characteristics (e.g. body mass) affect offspring traits during growth (Côté and Festa-Bianchet, 2001a; Gendreau et al., 2005), and because caring of offspring imply reproduction and high lactation cost for mothers (Clutton-Brock et al., 1989), we hypothesized that non-reproductive mothers and those with a high body mass (corrected for age) would be involved in more post-weaning associations than other females. Similarly, we hypothesized that high population size (used as an index of intraspecific competition) would increase the probability of observing post-weaning associations. Because the costs of caring for an associated juvenile should add up to the high energetic demands of gestation and lactation for reproductive mothers, we predicted a negative relationship between current kid body mass and the frequency of associations for lactating mothers. Finally, we predicted that post-weaning associations would benefit juveniles by increasing their body mass during the year of the association, and their survival during the following year.
Methods
Study area
We studied a native mountain goat population at Caw Ridge (54° N, 119° W), in the foothills of the Rocky Mountains in west central Alberta, Canada. The population uses an area of 28 km2 composed of alpine tundra and open subalpine spruce (Picea engelmanii) forest ranging from 1 750 to 2 180 m in elevation. The area includes steep grassy slopes, hills, and cliffs that mountain goats use as escape terrain (Festa-Bianchet and Côté, 2008). The subarctic-arctic climate is characterized by long and harsh winters followed by short and cool summers. Snowfalls can occur anytime during the year. High quality food resources are available only during a short period of the year (Festa-Bianchet and Côté, 2008), usually from June to August. Total population size (in June) increased from 76 individuals in 1991 to a maximum of 164 goats in 2008, and then decreased to 93 in 2012. Main predators of mountain goats at Caw Ridge are grizzly bears (Ursus arctos), wolves (Canis lupus), and cougars (Puma concolor). Other potential predators are black bears (Ursus americanus), wolverines (Gulo gulo), coyotes (Canis latrans) and golden eagles (Aquila chrysaetos) (Festa-Bianchet and Côté, 2008).
Captures
To assess the association between a mother and its juvenile(s), we used long term observational data of marked mothers with surviving juveniles of 1, 2 and 3 years of age. Beginning in 1988, we captured goats from mid-May to mid-September (n = 870 captures of 443 individuals; 235 females, 208 males) using a drop net in 1988 and thereafter in remotely controlled wooden box traps and self-tripping Clover traps baited with salt (Côté et al., 1998a). Capture and marking procedures lasted between 20 and 40 minutes. From 1996 onward, all individuals were captured and marked at 1 year of age with coloured Allflex plastic ear tags. All females 2 years of age and older were also marked with visual canvas collars that allow identification from long distances. For goats that were first captured as adults prior to 1996 (ca. 10% of the total population), age was estimated according to the number of horn annuli, a technique reliable up to 7 years of age (Côté et al., 1998b). Since
1993, 98% of ≥ 1 year-old goats have been marked and are of known age. More details on capture and marking techniques can be found in Côté et al. (1998a).
Post-weaning associations
From mid-May to mid-September (1991-2012), we located nursery groups almost daily and identified all individuals, including adult mothers and their juveniles of 1 to 3-year-olds. Adults were defined as individuals aged ≥ 4-years-old, juveniles as young aged 1 to 3-year-olds, and kids as 0 to 4-month-olds. We defined post-weaning associations as associations between adult mothers and their offspring aged 1 to 3-year-olds (i.e., excluding kids). We recorded the behaviour of all mothers and their offspring (1 to 3-year-olds) using binoculars (10x) and spotting scopes (15x-45x) between 0600 and 2300 h, weather permitting. From 1991 to 2012, the number of adult females ranged from 18 (1991) to 55 (2007) (mean ± SE: 40.0 ± 1.85). Parturition mainly occurred between 20 May and 3 June (Côté and Festa-Bianchet, 2001a). We determined kid sex by observing urination posture and by looking for a vulvar patch in females (Côté and Festa-Bianchet, 2001a). No case of twinning were observed during the study period.
We observed the presence of post-weaning associations mostly during rumination and resting periods when mothers laid down next to their associated juveniles and did not tolerate the close presence of other individuals except for their associated juveniles and kid (Festa-Bianchet and Côté, 2008). For each nursery group, we first evaluated whether juveniles were in the same group as their mother or not. When it was the case, we evaluated whether a juvenile was within a distance of ca. 1.5 m from its mother (a distance equivalent to the length of an adult goat), and if so we defined the post-weaning association as present. Associations were also recorded when juveniles were closely following their mother during walking. Each observation of a nursery group was considered a single observation for each dyad when the two individuals were present and represented a possibility for mothers and juveniles to be together. In addition, we considered that all proximity behaviour observed between mothers
and juveniles represented mother-juvenile associations, considering the total number of times the mother and juvenile were seen in the same group as a weighting factor. The association between a mother and its juvenile older than 1-year-old does not necessary imply that the dyad was associated the previous year(s).
Kid survival
We considered that kids had survived through summer if they were still present at weaning, which we set as 15 September (Côté and Festa-Bianchet, 2001b). At this date, suckling periods are much shorter and less frequent. Winter survival was evaluated on 1 June the following year. We identified mothers whose kids survived the winter by observing nursing behaviours of yearlings the following spring (see details in Gendreau et al., 2005) and by confirming parental links with DNA analysis from ear samples taken during marking (see details in Mainguy et al., 2009).
Reproductive status
We assigned a maternal reproductive status annually: (1) barren, i.e. mothers without a kid and that did not lactate; (2) summer yeld, i.e. mothers whose kid died before 15 September; and (3) lactating, i.e. mothers whose kid survived up to 15 September (weaning). The reproductive status of lactating mothers was determined by the observation of suckles.
Social rank
We used ad libitum sampling to record agonistic interactions between adult females from late May to mid-September each year (Côté, 2000). For each interaction, the identities of the initiator, winner, and loser were recorded. An individual was classified as the loser when it moved away > 1.5 m from its antagonist (Locati and Lovari, 1991; Drews, 1993) and showed submissive behaviour such as orientation avoidance (avoiding the antagonist by walking or
staring) and rush avoidance (quickly moving away from the antagonist) (Chadwick, 1977). We used the h’ index (De Vries, 1995), varying between 0 (nonlinear) and 1 (perfectly linear), to calculate the annual linearity of dominance hierarchies. We compared matrices using randomization (10 000 iterations) to verify whether h’ values were significantly linear (De Vries, 1995). Because annual dominance hierarchies were always significantly linear (Côté, 2000), we ordered adult females in annual hierarchies using De Vries' method (1998; see Côté, 2000 for details). All procedures were implemented in Matman 1.0 for Windows (Noldus Information Technology, 1998). Because social rank is highly correlated with age (r > 0.9; Côté, 2000), we used the residuals of a polynomial regression between social rank and age to obtain annual age-specific social ranks.
Body mass
Captured goats were weighed (± 0.5 kg) using a spring scale. Since 2001, three remotely controlled electronic platform scales baited with salt were also used to record body mass
ad libitum during the summer (Bassano et al., 2003). This procedure allowed the recording
of goat body mass remotely. Body mass of mothers, juveniles, and kids was annually adjusted to 15 July. We averaged daily summer mass gain separately for each age classes, up to 6 years old, and females ≥ 7 years old were averaged together (Hamel et al., 2010). Because body mass increased with age (up to 7 years old; Festa-Bianchet and Côté (2008)), we used the residuals of the regression of mass on age to calculate annual age-specific body mass by individual (but for kids age-specific body mass was adjusted based on birthdate).
Statistical analyses
We noted the presence of post-weaning association for each mother-juvenile dyad for every observation during summer. We defined occurrence of association when the mother associated at least 10% of the time with one of its juveniles to minimize potential biases on association indices generated by small sample sizes. We also calculated the frequency of associations for every mother-juvenile dyads creating an association index by dividing the
number of associations seen within a dyad by the number of times the dyad was observed during summer. We considered potential associations (dyad mother-juvenile) with 1-year-old (YL) (n = 234), 2-years-1-year-old (2Y) (n = 167) and 3-years-1-year-old juveniles (3Y) (n = 113). We excluded data from mothers whose age, age-specific social rank, age-specific body mass or reproductive status was uncertain. We also excluded individuals that died from unnatural causes or had juveniles of unknown sex and no available body mass. Using these criteria, we retained a maximum of 102 potential associations with 1-year-old (44% of all potential associations, 72 with 2-year-olds (43%), and 53 with 3-year-olds (47%). Four maternal intrinsic characteristics were tested: i) reproductive status (3 categories), ii) age, iii) age-specific social rank, and iv) age-age-specific body mass. We considered 3 extrinsic variables: i) population size (because adult males and females are segregated most of the year, population size was defined as the sum of adult females and 1 to 3-year-old individuals on 1 June, thus excluding adult males and kids), ii) juvenile sex, and iii) juvenile mass.
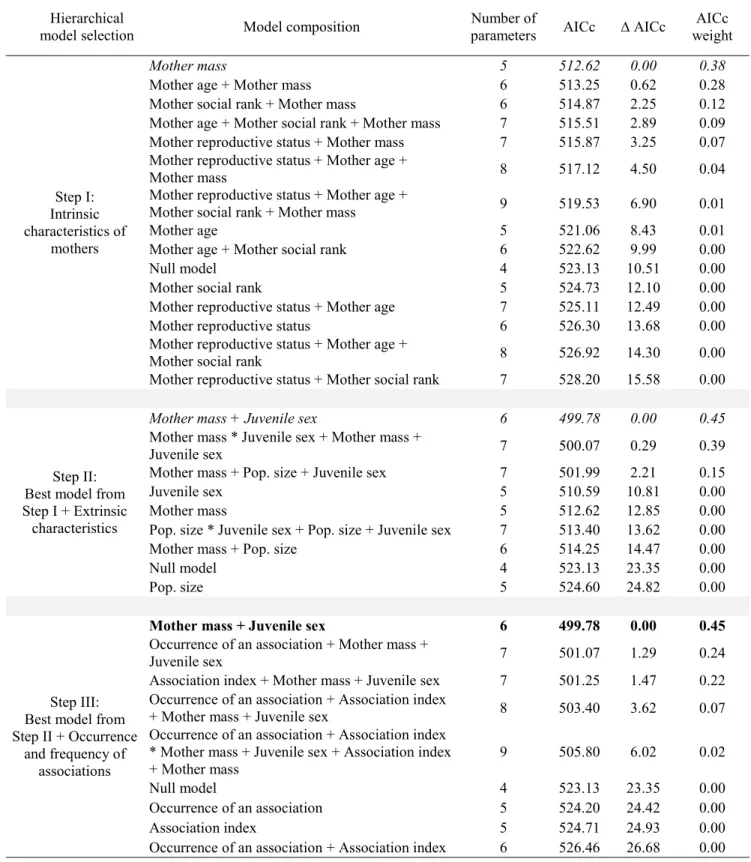
For all analyses, we developed a set of hierarchical candidate models to test a priori hypotheses (Appendix 1). Because we had several estimated parameters relative to sample size, we used the Akaike Information Criterion corrected for small sample sizes (AICc) to select the most parsimonious model (Hurvich and Tsai, 1995). We considered that models that differed in AICc by ≤ 2 received equivalent support (Burnham and Anderson, 2004). When multiple models were equivalent, we selected the least parameterized as parameters included in a more complex model selected within 2 AIC units should be considered as uninformative parameters (Arnold, 2010). We used maximum likelihood for all mixed model selection because models contained different fixed effect structures (Bolker et al., 2009).
For model selection, we used a separated steps approach adapted from Hosmer and Lemeshow (2004), because this method is recommended when multiple variables may be correlated. Using the “AICcmodavg” package (Mazerolle, 2015) in R software version 3.2.0, we first determined the most influential maternal intrinsic characteristics, and added the
selected variables in the second round of selection that included models with extrinsic characteristics unrelated to mothers. Finally, we added the most influential variables determined in the first two steps of selection in the final selection procedure to assess the influence of the occurrence of post-weaning associations and their frequency. In mixed effect models, we refitted the final selected model with restricted maximum likelihood to estimate the parameters of fixed effects and we obtained the p-values using the “lmerTest” package (Kuznetsova et al., 2015) in R. Model selection for the ZINB analysis (see details hereafter) was only performed for the first two steps of selection.
Because the association variable was binary and included many counts of zero, we used a zero-inflated negative binomial model (ZINB) (Yau et al., 2003) to evaluate the effect of the various covariates on the occurrence and frequency of post-weaning associations. We preferred the ZINB to the Poisson regression because it is most suitable for count data with excess zeros and overdispersion (Yau et al., 2003). The ZINB, also called mixture models of two distributions, first models the probability that an event occurred with a logistic regression, and then models the frequency of these occurrences with a negative binomial regression (Hilbe, 2011). Therefore, the model selection is performed on the two parts of the analysis (occurrence and frequency), as the frequency is necessarily linked to its occurrence. Using the hierarchical selection procedure explained above, all variables, including random variables, were included in both parts of the model, i.e. the zero-inflated and the count part. The ZINB analysis requires to have at least one observation in each category, however, for the analysis with yearlings only, there were no females in the barren and unassociated category. Thus, in order to perform the ZINB analysis and model the probability of association, we followed the “empirical logit method” (Agresti, 2013) and added a "new" female in the barren and unassociated category. To minimize the impact of this "new" female on the response variable, we used the mean value of each explanatory variable for this individual (Agresti, 2013). We performed the ZINB analysis using the NLMIXED procedure in SAS software version 9.4 (SAS Institute Inc., 2013).
We used generalized linear mixed models (GLMM) to assess the influence of the occurrence of associations and the association index on the probability that a mother did not reproduce the following year. We performed the analysis with the “glmer” function with a logit link function using the “lme4” package (Bates et al., 2014). Sample sizes (YL: n = 27; 2Y: n = 21; 3Y: n = 20) were too low to use mixed models to test the influence of the occurrence of associations and the association index on age-specific kid body mass. We thus performed a linear regression using maternal age, maternal specific social rank, and maternal age-specific body mass as covariates as well as population size, and kid sex using the “stats” package (R Core Team, 2015) in R. We used linear mixed models (LMM) to assess the influence of the occurrence of associations and the association index on yearling (n = 101), 2-years-old (n = 71) and 3-years-old (n = 51) age-specific body mass. We fitted hierarchical models including maternal characteristics, population size and juvenile sex as covariates. The probability of survival of yearlings (n = 102) and 2-year-olds (n = 72) the year after the post-weaning association was assessed using a GLMM with logit link. The occurrence of associations and the association index, maternal characteristics, population size, juvenile mass and juvenile sex were used as covariates again using a hierarchical selection procedure. Based on preliminary power analyses, we decided to exclude associations with 3-year-olds because they were too rare in the population. More than one juvenile could be associated with a mother during a summer, leading to multiple association counts for some mothers. To account for these repeated measures, we set mother identity as a random effect in all models. We also set year as a random effect to account for possible annual variation in other covariates that were not included in the models. Significance level was set at P < 0.05 for all analyses.
Results
From 1991 to 2012, 163 out of 234 mothers (70%) were associated at least 10% of the time with their yearling, 33 out of 167 mothers (20%) were associated at least 10% of the time with their 2-years-old, and 15 out of 113 mothers (13%) were associated at least 10% of the
time with their 3-years-old. The relatively low sample size precluded analyses of associations with 3-year-olds.
Factors favoring post-weaning associations
Maternal reproductive status was the only intrinsic characteristic explaining post-weaning associations with yearlings (Table 1, step I). We thus included this covariate in models assessing the influence of extrinsic characteristics on the occurrence and frequency of associations (which were grouped in the ZINB analysis). Again, maternal reproductive status was the best model (Table 1, step II). The occurrence of an association was 2.1 times higher for barren mothers than summer yeld mothers (t = -3.60; df = 56, p = 0.0007), and 1.8 times higher than for lactating mothers (t = -3.54; df = 56; p = 0.0008). Barren mothers also had significantly more associations with yearlings (as measured by the association index) compared to other mothers (4.8 times more than summer yeld mothers: t = -4.42; df = 56,
p < 0.0001; 3.1 times more than lactating mothers: t = -5.64; df = 56, p < 0.0001; Figure 2A).
Because barren mothers were set as the reference reproductive status in the analysis, we performed a post hoc Wald multiple comparison test to identify differences with the other reproductive status. The multiple comparison tests revealed that the occurrence of an association as well as the association index did not differ between summer yeld mothers and lactating mothers (all p-values ≥ 0.31).
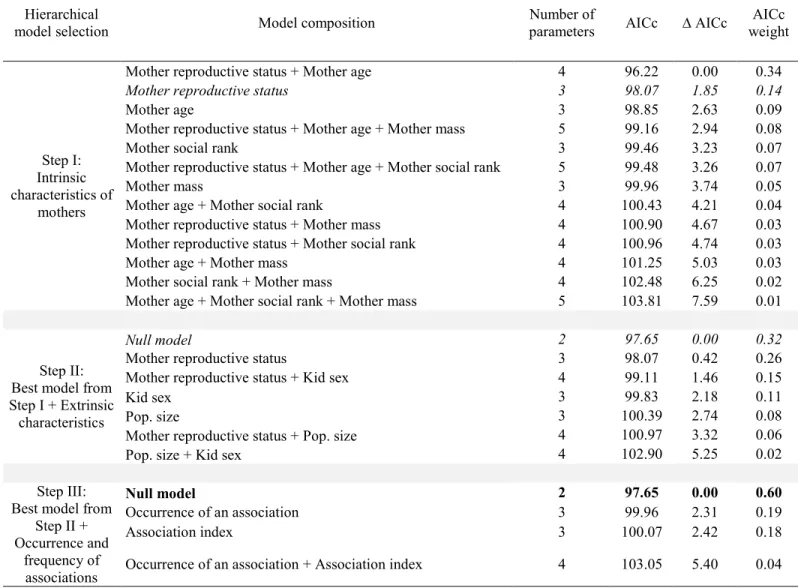
In post-weaning associations with 2-year-olds, no maternal intrinsic characteristics affected the association (i.e., the null model was retained; Table 2, step I). After adding additional models to assess the influence of extrinsic characteristics on the occurrence and frequency of associations, the best model explaining post-weaning associations with 2-year-olds included juvenile sex only (Table 2, step II). Mothers were more likely to be associated with 2-years-old when the juvenile was a female (females: 82%, males: 18%; juvenile sex: t = 2.05; df = 41; p = 0.047), but the association index did not vary with juvenile sex (t = 1.40; df = 41;
Costs of post-weaning associations for mothers
Reproductive status in the year following the association
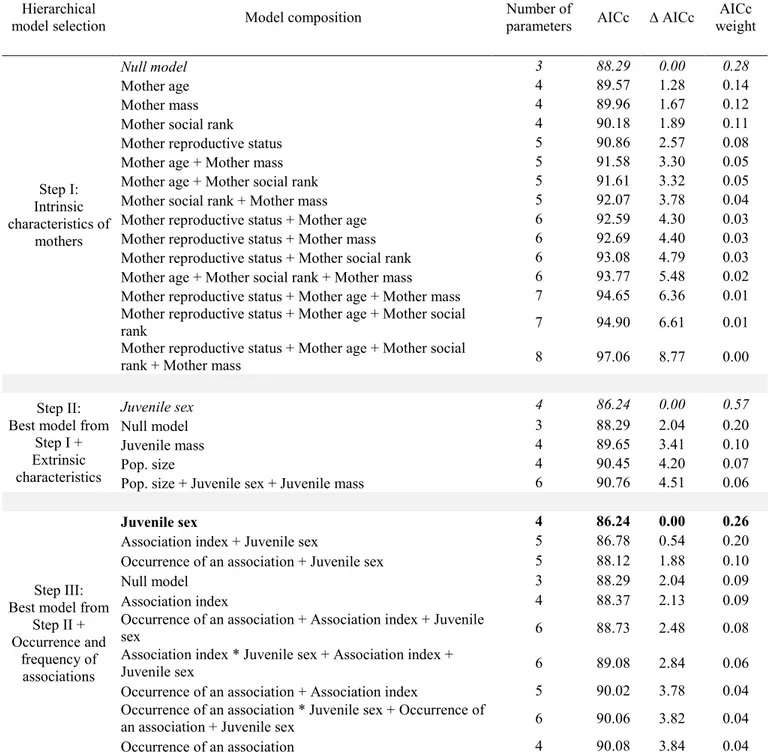
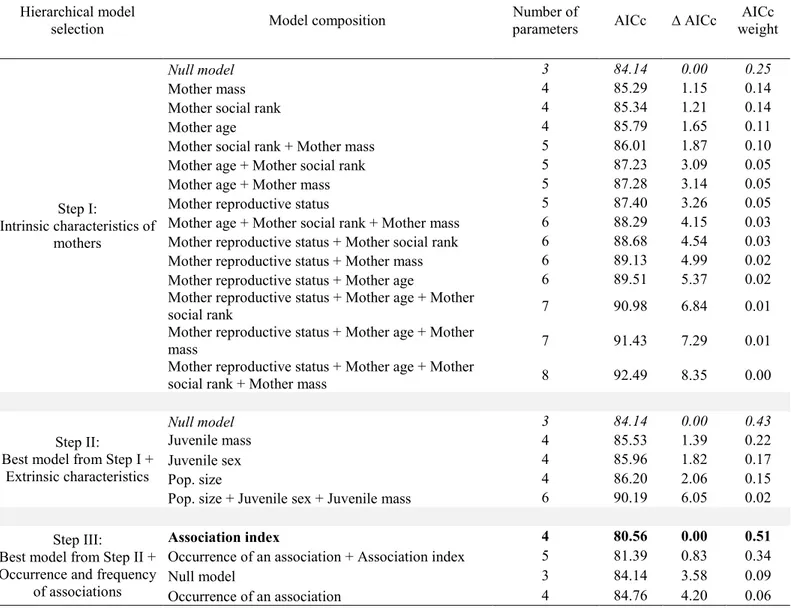
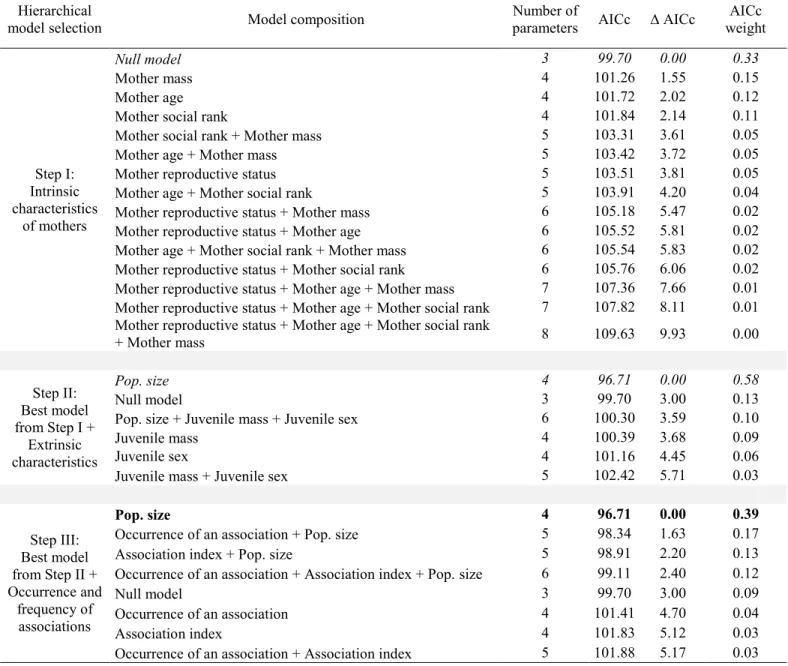
For potential associations with yearlings, the null model was selected when testing the effect of maternal intrinsic characteristics on their reproductive status the following year (Table 3, step I). When adding models composed of maternal extrinsic characteristics and the occurrence and frequency of associations, only juvenile sex was retained (Table 3, steps II and III). Mothers had a 35% probability of being barren the following year when they had a juvenile yearling male, compared to 17% when they had a juvenile yearling female, no matter if they were associated with it or not (male sets as reference juvenile sex, effect of juvenile sex: n = 89; β = 1.4 ± 0.7; z = 2.0; p = 0.05). For 2-years-old, the null model was also selected when testing the effect of maternal intrinsic characteristics on the maternal reproductive status the following year (Table 4, step I). When adding models composed of maternal extrinsic characteristics, no variables were selected (i.e., the null model was retained; Table 4, step II). We found no support for any of the models assessing the effect of an association with a 2-years-old on maternal reproductive status the following year. In contrast, the association index with 2-year-olds was retained as an influential covariate (Table 4, step III), showing a tendency for mothers to have a greater probability of being barren the following year if the association index with a 2-years-old was high (n = 64; β = 5.1 ± 2.8; z = 1.8; p = 0.07). Although the association did not significantly influence the reproductive status, 25% of females were barren when associated with a yearling compared to 23% of females that were barren when not associated; and 50% of females were barren when associated with a 2-years-old compared to 27% of females that were barren when not associated.
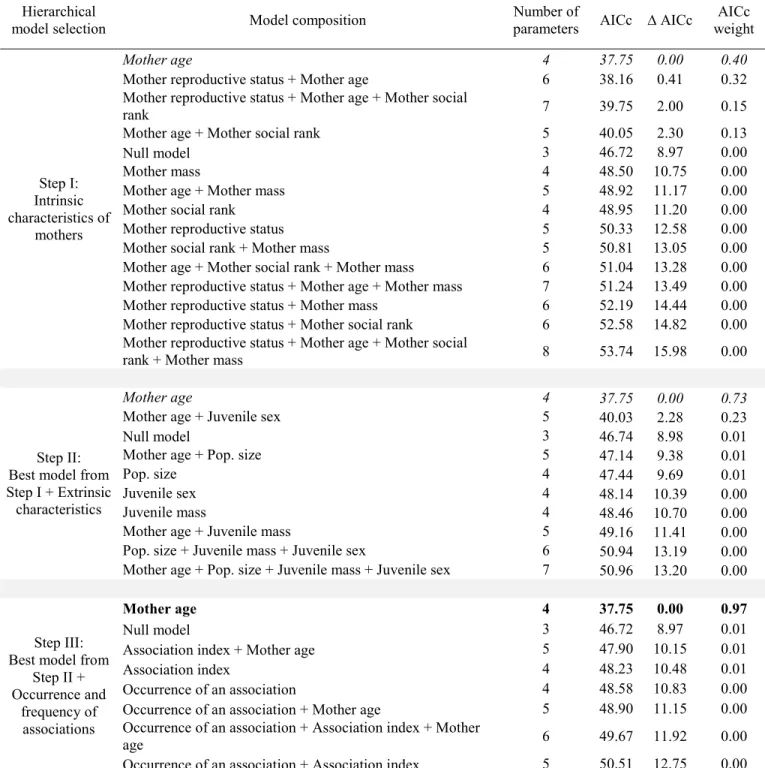
Age-specific body mass of kids during the year of association with a juvenile
The best model testing the effect of maternal intrinsic characteristics on age-specific body mass of kids was a model composed of maternal age only for yearlings (Table 5, step I), and