X-ray analysis of the NMC-A beta-lactamase at 1.64-A resolution, a class A carbapenemase with broad substrate specificity
Texte intégral
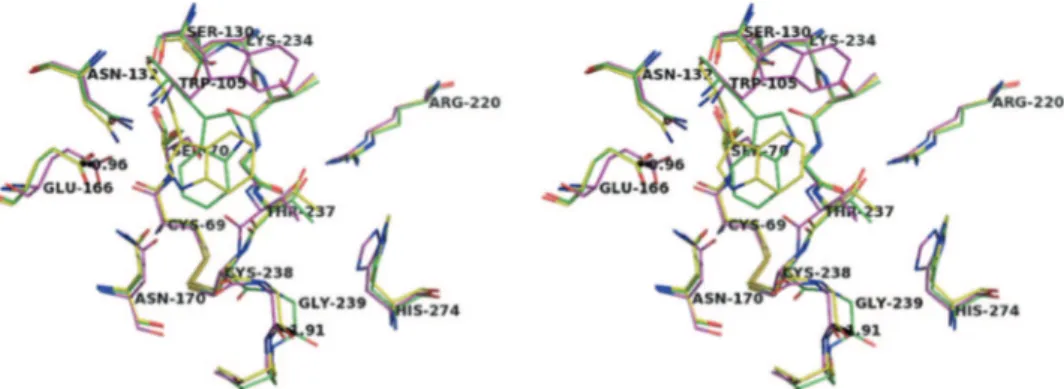
Figure
Documents relatifs
Pour montrer que cette conjecture est toujours vraie, on désigne le premier des quatre entiers par la lettre n.. Exprime alors les
c) de l'équipement de manutention ou de préparation d'aliments. 1) La tuyauterie dont le niveau ne permet pas l'écoulement par gravité dans un branchement d'égout
The text of the message displayed above also illustrates two other features of the Conversation Map system as a Usenet newsgroup browser: (1) Since
[r]
[r]
Given the steady, apparently hard emission, the X-ray flux variability may well arise purely from the soft component.. The VLBI data reveal an extended radio source with a size of
In this section, we describe three exact algorithms for solving the acyclic 2-coloring problem, namely a cycle-identification algorithm, a backtracking algorithm and a
La toute première étape du cycle de réplication viral s’effectue lorsque le virus rencontre une cellule hôte et qu’il s’y fixe. Lors de cette étape d’attachement,
