uNrvERSrrÉ
DU
euÉBEC À
Rrvrousrr
CoEXISTENCE
D'LINCoMPLEXE I'gSPÈCES
CRYPTIQUESÀ
pps
Écngrrps
LocALEs
pr
nÉcIoNALES
ruÈsp
pnÉsBNrÉs
COMME EXIGENCE PARTIELLE
DU DOCTORAT
ENBIOLOGIE
PAR
KAVEN
DIONNE
A
mes parents Chantal et André<< We need not mqrvel at extinction; if we must
marve|
letit
be at our own presumption inimaginingfor
a moment that we understand the many complex contingencies on which the existence of each species depends. >La
présente thèse estle
fruit
d'un
nombre incalculable d'heures passées aulaboratoire,
de
plusieurs
nuits
blanches d'analyse
et
de
quelques
momentsd'inspiration
pour
coucherle tout
sur papier.
La
plupart
desefforts
physiques et mentaux qu'arequis
la
production
de
ce
document sontde
moi,
maisje
n'aurais jamais réussi à terminer cette tâche colossale sans les apports de plusieurs personnesque
je
tiens à remercier de tout cæur.Merci
à Christian Nozais,mon
directeur de recherche, de m'avoir à nouveaufait
confiance pour mener à terme un projet d'une telle envergure. Merci pour ton aide surle
terrain et aulab
et merci pour tes conseils.Merci
également aux étudiants deton laboratoire, en
particulier
Geneviève Brouillet-Gauthier et Patricia Glaz, pour les conversations etle
partage de trucsqui
m'ont permis d'avancer. Bonne chance à tous dans vos projetsfuturs!
Merci
également à France Dufresne,ma
co-directrice,et
àtous ses étudiants,
pour
m'avoir encore permis de squattervotre
laboratoire pendant toutes ces années.Encore une
fois,
un merci
spécialà
RolandVergilino,
mon
mentoret
ami, pour tes conseils, ta patience et tous ces moments passés au lab ou devant une bonne bière méritée.Un
gros merci aux
étudiantsqui m'ont
aidé sur
le
terrain etlou
enlaboratoire
:
Laurence
Paquette,Michèle
Pelletier-Rousseau,Hubert
Philippon, Mirana Miharisoa Rakotomanga,Félix
St-Pierre etAstrid
Tempestini. Merci pour vosiii
efforts et
votre
dévouement.Merci
àAurore Levivier, Marion Trudel
etDavid Xie,
pour
I'intérêt que
vous
avez
manifestépour mon projet de
doctorat
et pour
lesHyalella
lorsquej'ai
eu
la
chancede
vous mentorer
dansvos
projets
respectifs. J'espère que ces expériencesont
été aussi enrichissantes pour vous qu'ellesI'ont
été pour moi!Merci
aux techniciens et au personnel de soutien deI'UQAR
et deI'ISMER:
Geneviève
Allard,
Louise
Amyot,
Mathieu Babin,
James Caveen, Jonathan Coudé, PaulineDubé,
Steeven Ouellet,Danie
Massé,Louiselle
Lévesque, Cécile Renault, PascalRioux,
Marie-AndréeRoy et
Mélanie
Simard.Merci
égalementà
Jonathan Gagnon de I'UniversitéLaval. Merci
pour tous ces coups de pouce que vous m'avez donnés chacun à votre tour.Merci
à Dominique Arseneault,Alain
Caton, GwénaëlleChaillou,
Robert Chabot, RichardCloutier, Mathilde
Couturier,Éric
Harvey, Yves Lemayet
Hacène Tamdrari. Pourvos informations, vos
conseils,votre
aide.Merci
égalementaux
gens
qui
ont
acceptéd'évaluer
les
différentes
épreuvesde
mondoctorat
:
Dominique Gravel, Richard Cloutier, Magella
Guillemette,
Jean-ÉricTremblay
et
GescheWinkler.
Un
merci particulier
à
Beatrix
Beisner,
MathewLeibold et
GescheWinkler
pour votre
temps passéà
évaluerla
présente thèse. Jepense
que
vos
conseilset vos
critiques
constructivesm'ont
permis
de
beaucoup m'améliorer en tant que scientifique.Merci
au personnel de la Réserve faunique Duchénier, de la Réserve faunique Rimouski, de la Réserve faunique Matane, du Parc national du lac Témiscouata, de la Zec Bas-Saint-Laurent, de la Zec Owen, de I'Organisme de Bassin Versant duNord-Est du
Bas-Saint-Laurentet aux
différentsvillégiateurs du
Bas-Saint-Laurent,pow
vos teffitoires.
Merci
au Conseil de recherches en sciences naturelles et en génie du Canada, à EnviroNord et à I'Association internationale des études québécoises pour le coup de poucefinancier; à
Boréaset
au Centre d'études nordiquespour le
prêt
du matériel de terrain.Enfin, un merci très spécial à ma
famille
qui m'a appuyé durant toute l'épreuve des études supérieures. Je remercie mes parents Chantal et André, mon frèreMichel,
ma
tarrte Denise, mon oncle Gaston et mes cousins Josée et Frangoispour
avoir cru enmoi
et enma
capacité à mener à termeun tel
projet.Merci
à mes beaux-parents Serge et Nicole pour leurs encouragements.Merci
aussi à Pascale Tremblay pour son amitié.AVANT-PROPOS
La
présente thèse de doctorat est présentée sousla
forme de cinq chapitres. Lepremier
et le
cinquième chapitre
correspondent respectivementà
l'introduction
générale et à la conclusion générale de la thèse. Les chapitres 2, 3 et 4 sont rédigés en anglais
et
sontécrits
sousla forme
de manuscrits soumisou
en préparation pour soumission à des revues scientifiques avec comités de lecture.Le
chapitre2
est enrévision pour publication dans la revue
Hydrobiologia,le
chapitre 3 est en voie d'être soumis pour publication à la revueEcologt
alors que le chapitre 4 est en préparation pour soumission à la revue Ecology Letters.Ce projet
a
été frnancé parle
Conseil de recherches en sciences naturelles et engénie
du
Canada(CRSNG)
grâceà
une subvention octroyéeà
ChristianNozais
etgrâce
à
une
bourse d'étude doctorale
accordéeà
Kaven Dionne.
K.
Dionne
aégalement reçu une bourse de recherche interdisciplinaire de la part du programme de
formation
FONCER
du
CRSNG
en
sciences
environnementales
nordiquesEnviroNord
ainsiqu'une
bourse de voyage dela
part del'Association
internationale des études québécoises.La
contribution de chacun des auteurs des articles présentés dansla
présente thèse est précisée au début de chaque chapitre.Bien
que Christian Nozais et FranceDufresne soient
co-auteurs
de tous les
articles
et
Dominique Gravel,
Rolandmanuscrit
doit
être considéré commela
thèse del'étudiant. L'apport
de C. Nozais etde
F.
Dufresne
en
a
été
essentiellementun de
supervision,
bien que
leurs commentaireset
ceux deD.
Gravel,R. Vergilino
etN.
Mouquet aient grandement amélioré la qualité du manuscrit.TABLES
DESMATIÈRES
REMERCIEMENTS
...ii
AVANT-PROPOS
...vLISTE
DESFIGURES...
...xiiLISTE
DESTABLEAUX...
...xviii
LISTE DES
ABRÉVIATIONS
...XXi nÉsuHaÉcÉNÉnar
..xxiw
CHAPITREI
TNTRODUCTTONCÉNÉnarE
...
... 11.1
Écologie descommunautés...
...11.2
Qu'est-ce qu'unecommunauté?...
...31.3
Nicheécologique
...41.4
Dérive écologique et théorieneutre
...91.5
Dispersion etmétacommunautés
... 111.6
Laspéciation...
...171.7
Distinguer les mécanismes liés à la formation des communautés... 181.8
Que sont les espècescryptiques?
...201.9
Le complexeHyalella
azteca...
...221.9.1
Coexistence des espèces du complexe ËL azteca à l'échelle locale ...231 . 1
0
Obj ectifs et hypothèses...:.
...251.10.1
Objectif
1 : Fractionnement isotopique et renouvellement des tissus des amphipodes...
...271JA.2
Objectif 2 : Diète des espèces cryptiques de ËL azteca dans les lacs...30CHAPITRE
II
HIGH
DIFFERENCES
IN
TROPHIC ENRICHMENT FACTORS
AMONG
FRESHWATER
AMPHIPODS FROM
THE HYALELLA AZTECA COMPLEX ....372.4
Résumé...
...382.1
Abstract....
...392.2
Introduction...
...402.3
Material andmethods...
...422.3.1
Collection ofanimals
...422.3.2
Foodpreparation...
...432.3.3
First experiment: TEFsmeasurement
...442.3.4
Second experiment: Turnover ratemeasurement
...462.3.5
Stable isotopeanalysis
...482.4
Results...
...512.4.1
Growth assessment andsurvivorship...
...512.4.2
Tumover rateexperiment...
...542.4.3
TEFsmeasurement
...572.5
Discussion
...612.6
Conclusion
...6823 Acknowledgements....
...69CHAPITRE
III
DO CRYPTIC SPECIESHAVE
DIFFERENT DIETSIN
LAKES?...713.0
Résumé...
...123.1
Abstract....
...733.2
Introduction...
...743.3
Material andmethods...
...783.3.1
Sampling
...78ix
3.3.3
Stable isotopeanalysis
...833.3.4
Dataana1ysis...
...843.4
Results...
...863.4.1
Speciesoccuffence
...863.4.2
C:N valuesof
amphipods...
...913.4.3
Isotopic composition ofamphipods...
...923.4.4
Diet ofamphipods...
...1003.5
Discussion
...1133.5.1
Isotopiccompositions...
... 1133.5.2
Diet ofamphipods...
...1153.5.3
Nichepartitioning
andoverlap
...1203.6
Conclusion
...1243.7
Acknowledgements....
...124CHAPITRE
IV
ARE
CRYPTIC
SPECIES
COMMUNITIES EXPLAINED
BY
SPATIAL.
ENVIRONMENTAL
ORBIOTIC
PROCESSES?...
...,.1274.0
Résumé...
...1294.1
Abstract....
...1304.2
Introduction...
...1314.3
Material andmethods...
...t34
4.3.T
Amphipodscollection
....1344.3.2
Speciesidentification
anddescription
...1394.3.3
Measurements of the physicochemicalparameters
...I43
4.3.4
Relative rolesof
spatial processes and environmental conditions... 1444.3.5
Speciesco-occu1rence...
...1474.3.6
Populationgenetics
...1484.4
Results...
...1484.4.2
Relative rolesof
spatial processes and environmental conditions...1514.4.3
Speciesco-occurrence...
...1544.4.4
Populationgenetic...
.... 1554.5
Discussion
...1614.6
Conclusion
...1674.1
Acknowledgements....
...168 CHAPITREV
CONCLUSION
GÉNÉRALE...
... T715.1
Contexte et originalité de l'étude...
...1715.2
Rappel desobjectifs...
...1735.3
Principauxrésultats....
...1775.3.1
Fractionnement isotopique et taux de renouvellement des tissus... 1775.3.2
Diète des espèces cryptiques dans leslacs...
...1815.3.3
Répartition des espèces cryptiques à l'échellerégionale
...1845.4
Portée del'étude
...1895.5
Limitations del'étude..
...1935.5.1
Fractionnement isotopique et taux de renouvellement des tissus... 1935.5.2
Diète des espèces cryptiques dans leslacs...
...1945.5.3
Répartition des espèces cryptiques à l'échellerégionale
...1965.6
Perspectives...
...1985.6.1
Critèred'invasibilité...
...1995.6.2
Co-occurrence avec d'autresinvertébrés
...2005.6.3
Dispersion des amphipodes entrelacs...
....2t1
ANNEXES
....203ANNEXE
A
...
...204ANNEXE
8...
...2A5ANNEXE
C...
...206Figure
1.1 Les quatre mécanismesqui
forment les communautés selon la théorie del'écologie
des
communautésde
Vellend (2010).
La
spéciation,
la
dérive écologique,la
niche écologique(ou
sélection selonla
terminologie de Vellend(2010)
et
la
dispersion
vont
interagir
pour
déterminer
la
dynamique
des communautésà
plusieurs échelles spatialeset
temporelles.Figure
adaptée deVellend
(2010).
...2Figure
1,2
La
niche
écologique d'une espècedécrite
selon ses besoins(a) et
son impact(b)
sur deux ressources substituablesRl
etR2.
Le
graphique (a) montreI'isocline
de
croissance
nette
nulle
(ICi.IN)
de
I'espèce, c'est-à-dire
les combinaisonsde
densitésde
Rl
et R2 pour
lesquelles I'espècea
une
valeur adaptative absoluenulle
alors que les taux de natalité et de mortalité sont égaux.La
zonegrise
à
droite de
I'ICNN
correspondaux
densitésde
Rl
et R2
pour lesquellesla
population a
une croissancepositive
et
inversémentpour
la
zone blanche à gauche deI'ICNN. Le
graphique montre que I'espèce est moins limitée parRl
que par R2 car sa population a besoin d'une densité moindre deRl
pour croître.Le
graphique(b)
représente I'impactper
capita
d'une espèce surR1
etR2.
Les flèches grises montrent I'impact de I'espèce sur chacune des ressources de façon indépendante alors quela
flèchenoire
représentela
somme des deux vecteurset
s'appellele
vecteur d'impact.Figure
adaptéede
Leibold
(1995)
etChase et
Leibold
(2003).
...5Figure
1.3ICNNs
et vecteurs d'impacts pour deux espècesA
etB
consommant deux ressources substituablesRl
et R2 (a et b). Les
vecteursd'impact
de
chaque espèce(Ia
et
Is)
sont placésà
I'intersection desICNNs
des deux espèces afin d'examinerla
stabilité
deleur
interaction.Les lignes
entirets
correspondent àI'inverse
des vecteursd'impacts; les
points
situéesà
I'intérieur
de
ces
lignesindiquent les
densitésen
ressourcespour
lesquellesles deux
espèces peuvent potentiellement coexister, alors que les points situés au-dessus ou en-dessous de ces lignes indiquent les densités en ressources pour lesquelles la compétition de I'une des espèces rejettera I'autre dumilieu. Le
graphique (a) montrela
situation dans laquelle chaque espèce a un plus grand impact sur la ressourcequi limite
leplus
la
croissancede
sapopulation, de
sorte quela
coexistencelocale
de ces espèces sera potentiellement stable.Le
graphique(b)
montre
la
situation danslaquelle chaque espèce a un plus grand impact sur la ressource
limitant
le plus la croissancede
I'autre
espèce, résultanten une
coexistencelocale instable
des espèces. Si les ICNNs ne se recoupent pas, I'espèceayatt
les plus faibles besoins en ressources existera seule (situation non montrée). Figure modifiée de Chase etxnr
Figure 1.4
Représentation schématiquedes
quatre
perspectives présentées parLeibold
etal.
(2004) etHolyoak
etal.
(2005) pour expliquerle
fonctionnement des métacommunautés: la
dynamique des parcelles(a),
le
tri
des espèces (b), I'effet de masse (c) et les modèles neutres (d). Les schémas présentent la situation dans laquelle deux espècesA
etB
compétitionnent entre elles pour les ressources dumilieu.
Les flèches corurectent les populations d'où originent les migrants avec les sites potentiels à coloniser, désignésici
par les grand rectangles ou ovales; lesformes
differentes
désignentdes
milieux
aux
conditions
environnementales différentes. Les flèches solides indiquent une dispersion plus forte que les flèchesen
tirets;
la
tête
des flèchesindique
si
la
dispersionest unidirectionnelle
ou bidirectionnelles entre les sites. La forme de la boîte entourant le nom de I'espèce indique que I'espèce est la meilleure compétitrice dans sonmilieu
lorsque la boîte est dela
mêmeforme
quecelle du
site.En
(a),la
dynamique des parcelles est montrée pour un cas où la coexistence des espèces est possible. Les deux espèces ont développé un compromis entrela
capacité à compétitionner et la capacité à sedisperser
:
I'espèceA
estla
meilleure
compétitrice alors que I'espèceB
est lameilleure
colonisatrice.La
parcellevide pourrait
éventuellement être coloniséepar I'une
des
deux
espèces.En
(b),
chaque espècea
une niche
écologiquedifferente.
Le
tri
des espècesfait
que chaque espèce occupeun
site
selon lesconditions environnementales rencontrées alors que les
taux
de dispersion sont trop faibles pour pennettre aux espèces de coloniser desmilieux
où elles ne sontpas les meilleures
compétitrices.En (c), I'effet
de
masse permetaux
espècesd'être
présentesdans
les
milieux
moins
favorables
en raison
des
taux
de dispersion élevés àpartir
des sources. En (d), les modèles neutres supposent que les espèces ont des valeurs adaptatives similaires. Dans ce cas, les communautés sont régits par les processus stochastiques liés à la démographie des populations des espèces,par
la
spéciation et parla
dispersion aléatoired'individus
entre les sites. Figure modifiée de Leibold etal.
(2004)...
... 16Figure
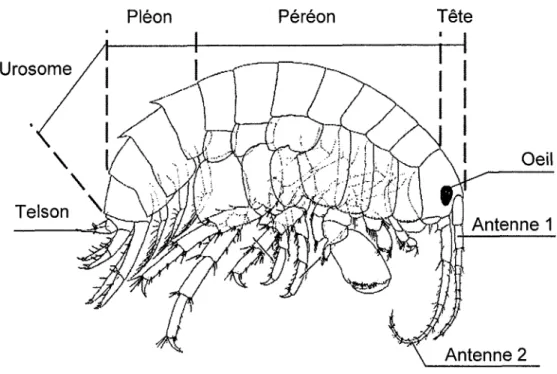
1.5 Schéma représentant les différentes structures du corps d'un amphipode du complexeHyalella
azteca. Figure modifiée de Bousfield (1973). ...23Figure
1.6 Diagramme schématique résumant la démarche de la présente thèse. ...26Figure
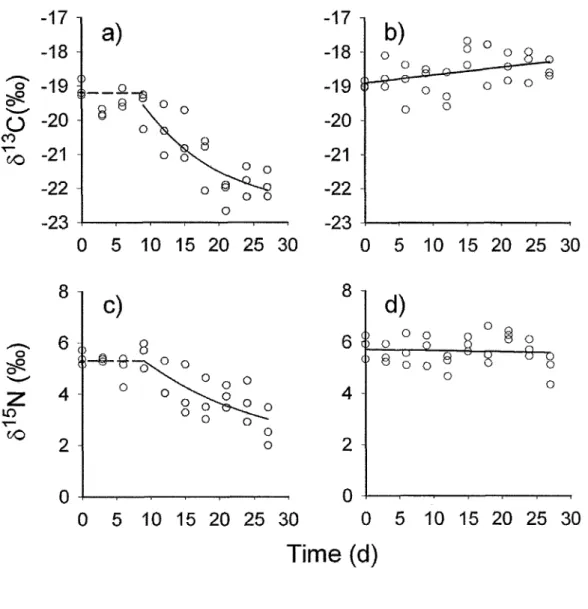
2.1Changesin
ôr3C and ô15Nin
Hyatella azteca amphipods (speciesB)
from
Lake
des Baies (Bas-Saint-Laurent, Quebec, Canada)during the
turnover rateexperiment,
when feeding
on
conditioned
birch
leaves
(a
and
c)
and
on periphytonin
the
controls(b
andd). For
a
andc,
solid
line
represents bestfit
nonlinear
regressionand
dashedline
representsthe delay during which
the isotopic compositionof
amphipods wasnot
yet changing;for
b
and d,solid
line represent least squares regression.dDays.
...56Figure 3.L
Carbon and nitrogen stable isotope compositionsof
amphipodsfrom
thedifferent
groups sampled at the three sitesof
Lac des Baiesin
June, August andOctober
2010. Circles
representH.
azteca
speciesA,
triangles
pointing
up representH.
azteca speciesB,
squares representH.
azteca species C, diamonds representH.
azteca speciesD;
signs areblack
for
amphipods sampledin
the shallow zone andwhite
for
amphipods sampledin
the deep zone. Letters at the leftof
signs denote amphipodswith
significantly different isotopic values...88Figure 3.2
Carbon and nitrogen stable isotope compositionsof
amphipodsfrom
thedifferent
groups sampledat
the three
sitesof
Lac
Fené
in
June,August
and October 2010.Circles
represent H. azteca speciesA
and squares represent 1Lozteca species C; signs are black
for
amphipods sampled in the shallow zone and whitefor
amphipods sampled in the deep zone. Letters at theleft
of
signs denote amphipodswith significantly
different isotopicvalues.
...89Figure 3.3
Carbon and nitrogen stable isotope compositionsof
amphipodsfrom
thedifferent
groups sampled at the three sitesof
Lac Neigettein
June, August andOctober
2010. Circles
representH. azteca
speciesA,
triangles
pointing
up representH.
azteca speciesB,
squares representH.
azteca species C, diamonds representH.
azteca speciesD
and trianglespointing
down represent G. lacustris;signs
are black
for
amphipods sampledin
the
shallow
zone
and
white for
amphipods sampled
in
the
deep
zone. Letters
at
the
left
of
signs
denote amphipodswith
significantly
different isotopicvalues.
...:.90Figure 3.4
Carbon and nitrogen
stable isotope compositionsfrom
potential
food sources(after
correctionfor
fractionation) andof
amphipodsfrom
the different groups sampled at the three sitesof
Lac
des Baiesin
June, August and October 2010. Red crosses represent detritus leaves,blue
crosses represent periphyton, green crosses representSOM, circles
representH.
azteca speciesA,
trianglespointing
up representH.
azteca species B, squares representH.
azteca species C anddiamonds
representH.
azteca speciesD;
signs
areblack
for
amphipods sampledin
the shallow zone andwhite for
amphipods sampledin
the deep zone.Black
crosses represent gastropods
and ('X))
represent
Daphnia Sp.,
for
xv
Figure 3.5
Carbon
andnitrogen
stableisotope
compositionsfrom
potential
food sources (after correctionfor
fractionation) andof
amphipodsfrom the
different groups sampled at the three sitesof
Lac Ferréin
June, August and October 2010. Red crosses representdetritus
leaves,blue
crosses represent periphyton, green crosses representSOM, cyan
crosses represent epiphyton,circles
represent 11.azteca species
A
and squares representH.
azteca speciesC;
signs are blackfor
amphipods sampled in the shallow zone and white
for
amphipods sampledin
the deep zone.Black
crosses represent gastropods and"X"
representDaphnia
sp.,for
comparison....
...109Figure 3.6
Carbon
andnitrogen
stableisotope
compositionsfrom
potential
food sources(after
correctionfor
fractionation) andof
amphipodsfrom the
different groups sampled at the three sitesof
Lac Neigettein
June, August and October 20T0. Red crosses represent detritus leaves,blue
crosses represent periphyton,green
crosses representSOM, circles
representH.
azteca speciesA,
trianglespointing
ap representH.
azteca species B, squares represent H. szteca species C, diamonds representH.
azteca speciesD
and trianglespointing
down represent G.lacustris;
signs areblack
for
amphipods sampledin
the
shallow
zone andwhite
for
amphipods sampled
in
the
deep
zone.
Black
crosses represent gastropods and'X"
represent Daphnia sp., forcomparison
...II2
Figure
4.1Map
of
the
locationsof
the 64
lakes sampledin
Bas-Saint-Laurent and Gaspésie (Eastern Quebec, Canada).Pie
charts representthe
fractionsof
each amphipod speciesin
the communitiesof
the study region as determinedfor
eachlake
for
30
individuals
(speciesA:
blue,
speciesB:
red,
speciesC:
green, speciesD:
purple, G.
lacustrls:
cyan).The
numbernext
to
a
pie
chartis
the number of the lake where the amphipods weresampled.
...135Figure 4.2
Scaling
2
biplot
of
the principal
component analysis(PCA) on
the morphological measurements made on the amphipods. Thefirst
and second PC axesexplain
59.60Â and 2Q.3oÂ, respectively,of
the
variation
in
the
data set.Arrows represent each morphological variables measured on the amphipods
(BL:
body
length,
HL:
head length,
ML:
mucronationslength,
EP:
eye
perimeter divided byHL, NM:
numberof
dorsal mucronations,TAL:
total antennae length divided byHL,
ANT2/1 : lengthof
antenna 2 dividedby
the lengthof
antenna 1).Circles
representH.
azteca amphipodsfrom
speciesA,
triangles
representI{
azteca amphipodsfrom
speciesB,
Crosses representH.
azteca amphipodsfrom
species C andXrepresent
H.
azteca amphipodsfrom
species D. The number next to a symbol is the number of the lake where the amphipods were sampled...150Figure 4.3
Correlation
between
the
log-transformed
Bray-Curtis
dissimilarity betweenamphipod communities
of
the
lakes
of
the
study
region
and
linearFigure
4.4Yariation partitioning
results showing the pure and shared influencesof
the
environmental component(upper
left
circle),
a
linear trend
(upper right
circle) and
the
spatial component
(lower circle)
on
amphipod
speciescomposition. Numbers represent adjusted
Rl
values;the empty fractions
have small negative adjustedRi
values..
... 153Figure 4.5
Correlation
for
eachH.
azteca species betweenthe
genetic
distance betweenpopulations
of
the
lakes
of
the
study region
calculatedas
Edwards distance and log-transformed geographicdistance.
...156Figure
4.6 Haplotype network
for
H.
azteca speciesA
in
the
lakesof
the
studyregion.
Eachpie
chart representsa
single
haplotype,with the
sizeof
the
pie proportionalto
the numberof
individualswho
possess that particular haplotype.The
romannumber next
to
the
pie
chart identifies the
haplotype.Each
color representa different
lake where haplotypes are present.The lines
between pie charts represent the relation between haplotypes; the numberof
black dots on aline represents the number of nucleotide differences between two haplotypes. 157
Figure 4.7
Haplotype network
for
H.
azteca speciesB
in
the
lakesof
the
studyregion.
Eachpie
chart representsa
single haplotype,with the
sizeof
the
pie proportionalto
the numberof
individualswho
possess that particular haplotype.The
romannumber next
to
the
pie
chart identifies the
haplotype.Each
color representa
difftrent
lake where haplotypes are present.The
lines between pie charts represent the relation between haplotypes; the numberof
black dots on aline represents the number of nucleotide differences between
two
haplotypes. 158Figure 4.8
Haplotype
network for
H.
azteca speciesC in
the
lakesof
the
studyregion.
Eachpie
chart representsa
single
haplotype,with
the size
of
the
pie proportionalto
the numberof
individualswho
possess that particular haplotype.The
romannumber next
to
the
pie
chart identifies the
haplotype.Each
color representa different
lake where haplotypes are present.The lines
between pie charts represent the relation between haplotypes; the numberof
black dots on aline represents the number of nucleotide differences between two haplotypes. 159
Figure 4.9
Haplotype network
for
H.
azteca speciesD
in
the
lakesof
the
studyregion.
Eachpie
chart
representsa
single
haplotype,with
the
sizeof
the
pie proportionalto
the numberof
individualswho
possess that particular haplotype.The
romannumber next
to
the pie
chart identifies the
haplotype.Each
color representa different
lake where haplotypes are present.The lines
between pie charts represent the relation between haplotypes; the numberof
black dots on axv11
Figure 5.1
Photos
à
l'échelle
d'amphipodes représentantles
différentes
espèces trouvées au Bas-Saint-Laurent et en Gaspésie. L'amphipodeH.
azteca de I'espèceA
provenait dulac
Caribou,celui
de I'espèceB
provenaitdu
lac Bébé,celui
deI'espèce
C
du lac
Ferré
et
celui de
I'espèceD
et
le
Gammarus lacustrisTableau 2.1 Isotopic
compositions(in
o/oo) andC:N
ratios
of
amphipodsfrom
thefield
andfrom
the fractionation experiment (mean+
SD,n
:
5
for all
samples exceptfor
amphipods feeding on cyanoperiphytonfrom
Lake France(n:
4) andfrom
Lake Ferré(n
:
1)). Valueswith
asterisks aresigni{icantly different from
values of amphipods from the
field....
...49Tableau 2.2Head
lengthsof
amphipods at the start and the endof
the fractionation experiment and numberof
survivors,for
each treatment....
...52Tableau
2.3 Two-wayANOVAs
to
assessif
amphipods grew over the courseof
the fractionation and tumover rateexperiment.
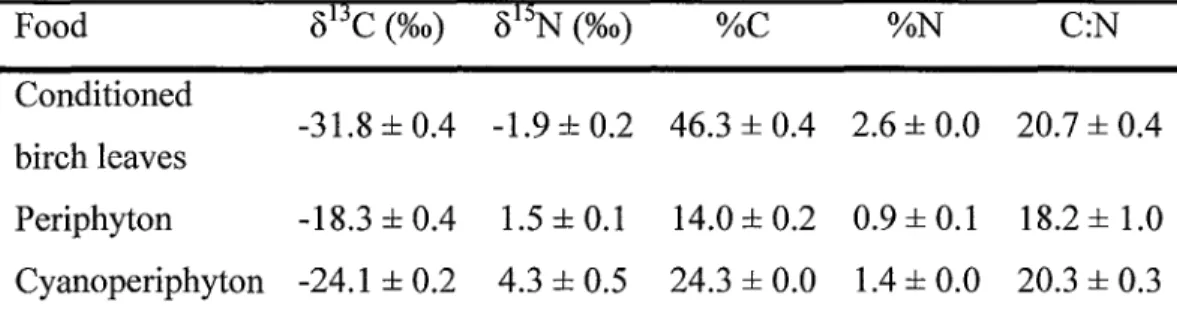
...53 Tableau 2.4 Carbon and nitrogen composition of foods (mean+
SD,n:3)
...57Tableau
2.5Trophic
enrichment factors(TEF) (in
%o)for
amphipodsfrom
thefirst
experiment (mean
+
SD,n
:
5for all
samples exceptfor
amphipods feeding on cyanoperiphyton from Lake France(n:
4) and Lake Ferré (n= 1))
...59Tableau
2.6Two-way
ANOVAs
to
assess the effectof
food type and lakeof
origin
of the amphipods on the fractionation
of
carbon and nitrogen isotopes...60 Tableau 3.1 Coordinates and site characteristics of lakes sampled in 2010. ...80Tableau 3.2
One-way
MANOVAs
results comparing
for
eachlake the
isotopic compositionsof
H. azteca amphipodsfrom
different
groups(different
species andlor sampling depths). Asterisks denote significant differences (P < 0.05). ...93Tableau 3.3 Two-way
MANOVAs
results comparing among monthsfor
each lakethe
isotopic
compositions
of
invertebratesfrom
different groups
(different speciesand/or sampling
depths).Groups:
G.
lacustris
from the
shallow
zone (GLs),H.
azteca speciesA
from the shallow zone(HAs)
or the deep zone(HAd),
H.
aztecq speciesB
from
the shallow
zone(HBs) or the
deep zone(HBd),
1Lazteca species C from the shallow zone (HCs) or the deep zone (HCd),
H.
azteca speciesD
from the shallow zone (HDs) or the deep zone (HDd). Asterisks denote significant differences (P <0.05).
...94XIX
Tableau
3.4
Mean isotopic
compositions
(in
o/oo)among sites
of
the
different potentialfood
sourcesof
amphipods sampledin
the lakesfor
each month (mean+
SD).
n
:
number
of
replicatesat the lake
scale; eachreplicate
exceptfor
Daphnia sp has a value based on 3 pseudo-replicates.
...
...l0l
Tableau
3.5
Bayesianmixing
model
(SIAR)
resultsfor
amphipods sampledfrom
each site
of
Lake des Baiesfor
each monthin
2010. Percentiles showthe
lst
to95th
range
of
potential
contribution.
Sourceswith
a
necessary contribution (minimal potentialcontribution>
}Yo) in bold. Groups:H.
azteca speciesA
from
the shallow zone (HAs),H.
azteca speciesB
from the shallow zone(HBs)
or the deep zone(HBd),
H. azteca speciesC
from the
deep zone(HCd),
H.
azteca speciesD
from
the
shallow zone(HDs)
or
the
deep zone(HDd). The
mention"out"
after the group name means that amphipods had isotopic compositions that were always completely outsideof
the
mixing
polygon
createdby
the
isotopic compositionsof
the food
sourcesafter
correctionfor
fractionation(nom
:
not enough organic matter detectedfor
analysis)....
...104Tableau
3.6
Bayesianmixing
model
(SIAR)
resultsfor
amphipods sampled from each siteof
Lake Ferréfor
each monthin
2010. Percentiles showthe
lst
to
95th rangeof
potentialcontribution.
Sourceswith
a necessary contribution(minimal
potential contribution
>
}Yo)in
bold.
Groups:H.
azteca speciesA
from
theshallow zone
(HAs)
or
the
deepzone
(HAd),
H. azteca speciesC
from
the shallow zone (HCs) or the deep zone(HCd)
(abs:
food
sourcenot
available atthe
site).
...107Tableau
3.7
Bayesianmixing
model
(SIAR)
resultsfor
amphipods sampled from each siteof
Lake Neigettefor
eachmonth
in
2010. Percentiles showthe
lst
to95th
range
of
potential contribution.
Sourceswith
a
necessary contribution(minimal potential contribution
>
0%o)in
bold.
Groups: G.Iacustris
from
the shallow zone(GLs),
H.
azteca speciesA
from
the shallow
zone(HAs)
or
the deep zone(HAd), H.
azteca speciesB from
the shallow zone (HBs),H.
azteca speciesC
from the
shallow zone
(HCs)
or
the
deep zone(HCd),
H.
azteca speciesD
from
the
deep zone(HDd).The mention
"out"
afterthe
group name meansthat
amphipods hadisotopic
compositionsthat
were always completely outsideof
themixing polygon
createdby the
isotopic compositionsof
the food sources after correctionfor
fractionation...
... 110Tableau
4.1
Coordinates, drainagebasin
and physicochemical parametersof
eachlake sampled
in
Bas-Saint-Laurent and Gaspésie (Eastern Quebec, Canada)(OM
:
organiô matter).(Cond:
pS cm-l,DOC
:
mg L-1, Chla:
mg m-3, TP:
pmolL-l,
TOCS:
mgof
organic matter per mgof
sediment) ... 136Tableau
4.3 Numberof
observed (Jo6r) and expected (J.,.0) occurencesfor
each pairof
amphipod specieswith
theprobability
that each pair could cooccur less (Px) ormore
(Pgt)if
their
distributions
weretruly
randomof
one
another. Asterisks denote significant differencesfrom
expected frequency (P < 0.05)...154LISTE
DESABREVIATIONS
abs
Food source absentfrom thelittoral
zoneAIC.
Corrected Akaike informationcriterion
ANT2/1
Length of antenna 2 divided by the length of antennaI
BL
Total body lengthCF-IRMS
Continuous-flow Isotope Ratio Mass SpectrometryChla
Chlorophyllaconcentration
COI
Cytochrome oxidaseI
Cond
Conductivitydf
Degree offreedomDOC
Dissolved organic carbonDs
Lake development indexEA
Elemental analysisEP
Eye perimeterGAs
Gasteropod sampled in the shallowpart
of thelittoral
zoneGIS
Geographic information systemGLs
Gammarus lacustris amphipod sampled in the shallowpart
of
thelittoral
zoneHAd
Hyolella
azteca amphipodfromprovisional
species A sampled in the deeppart
of thelittoral
zoneHAs
Hyatetla azteca amphipodfromprovisional
species A sampled in the shallowpart
of thelittoral
zoneHBd
Hyalella
azteca amphipodfromprovisional
speciesB
sampled in the deep
part
of thelittoral
zoneHBs
Hyalella
azteca amphipodfromprovisional
speciesB
sampled in the shallow
part
of thelittoral
zoneHCd
Hyalella
azteca amphipodfromprovisional
species C sampled in the deeppart
of thelittoral
zoneHCs
Hyalella
azteca amphipodfromprovisional
species C sampled in the shallowpart
of thelittoral
zoneHDd
Hyalella
azteca amphipodfromprovisional
speciesD
sampled in the deep
part
of thelittoral
zoneHDs
Hyalella
azteca amphipodfromprovisional
speciesD
sampled in the shallow
part
of thelittoral
zoneHL
Half-life
(chapitre2);
Total head length (chapitre 4)ICNN
Isocline de croissance nette nulleJexp
ExpectedfrequencyJobs
ObservedfrequencyLAl
Length of antennaI
LA2
Length of antenna 2MANOVA
Multivariate
analysisofvariance
MEM
Maran's eigenvector mapML
Mucronations lengthNM
Number of mucronations on thepleon
nom
No organic matter detectedfor the samplePC
Principal
componentPCA
Principal
component analysisPCR
Polymerase chain reactionPet
Probability
that two species are co-occurring morefrequently
than expectedif
the species were randomly associatedxxiii
Prt
Probability
thst two species are co-occurcing lessfrequently
than expected
if
the species were randomly associatedPZ
Pelagic zoneRDA
Redundancy analysisRFLP
Restrictionfragment length polymorphismSD
StandarddeviationSOM
Sediment organic matterSS
SumofsquareTAL
Total antennae lengthTEF
Trophicenrichmentfactor
TOCS
Total organic content of the sedimentsTP
Total phosphorusPlusieurs
mécanismesliés
à la
niche
écologique,
à la
dérive
écologique,
à
ladispersion
et à
la
spéciationparticipent
à la
formation
et
à la
dynamique
descommunautés
tant
à
des
échelles locales
que
régionales.Toutefois,
le
rôle
et fimportance
de
ces
mécanismessont
souvent
difficiles
à
distinguer
car
ces mécanismes peuventinteragir,
et
des
mécanismesdifférents peuvent mener
à
la formation de communautés semblables. Dans ce contexte, les communautés d'espècescryptiques, souvent
soupçonnéesd'être
écologiquement équivalentes,
semblent particulièrement intéressantespour
étudierle rôle
des mécanismesliés à la
dérive écologique et à la dispersion. Cette recherche de doctorat avait pourobjectifgénéral
d'approfondir nos connaissances surle
rôle respectif des mécanismes liés àla
niche écologique et ceux liés à la dérive écologique et à la dispersion dans la formation des communautés d'espèces cryptiques,tant
à l'échelle locale que régionale, en utilisant comme modèle les amphipodes du complexe d'espèces cryptiquesHyalella
azteca.Notre objectif
généralétait
divisé
principalementen deux
sous-objectifs. Premier sous-objectif : à l'échelle locale(le
lac),vérifier
si
les différentes espèces cryptiques du complexeH.
azteca occupaient des niches trophiques différentes. Préalablement à cetobjectif,
nous avonsvérifié si
des amphipodes du complexe -FI azteca provenant de differents lacs avaient des facteurs de fractionnement isotopique du carbone et de I'azote différents entre eux etlorsqu'ils
se nourrissaient sur des sources de nourriture différentes:
feuilles de bouleau conditionnées et deux types de périphyton. Ensuite,nous
avons échantillonné des
amphipodesdans
trois lacs
du
Bas-Saint-Laurent (Québec, Canada) àtrois
sitespar lac,
deux profondeurs ettrois
dates etutilisé
les isotopes stables du carbone et de l'azote pour comparer la composition isotopique des amphipodes avec celles de leurs nourritures potentielles. Deuxième sous-objectif:
àl'échelle régionale (plusieurs lacs),
déterminer I'importancerelative
des procédésspatiaux,
environnementaux
et
biotiques
pour
expliquer
la
composition
des communautés d'espèces cryptiques d'amphipodes Ë1. azteca. Pour cetobjectif,
nous avons échantillonné etidentifié
à I'espèce des amphipodes dans 64 lacs duBas-Saint-Laurent
et
de
la
Gaspésiepour
lesquelsnous
avons
également mesuré divers paramètres physicochimiques.Nous
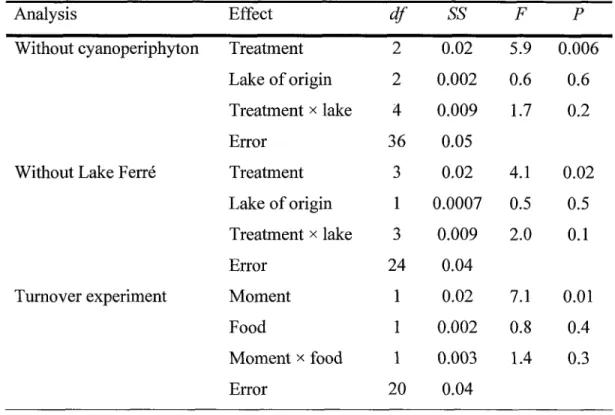
avons montré que les amphipodes avaient des differences significatives dans leurs facteurs de fractionnement isotopiques entre les traitements (environ 3,0%o à laXXV
fois
pour
le
carboneetl'azote). Le
fractionnement des amphipodes surle
carbone était davaîtage affectépar
le
type
denourriture
sur lequelils
se nourrissaient alorsque
le
fractionnementsur I'azote était
davantage affectépar
le
lac d'origine
des amphipodes.Nous
avons égalementmontré
que les. amphipodesont
renouvelé le carboneetl'azote
de leurs tissus en environ un mois.A
l'échelle locale, dans les lacs, les amphipodes de différentes espèces du complexeH.
azteca avaient des valeurs deô"C
et
de
ô"N
et
desutilisations de
leur
habitatqui
parfois
se chevauchaient et d'autresfois
se distinguaient. Comme des differences étaient souvent observées pour des amphipodes d'une même espèce échantillonnésà
différentes profondeurs, cela suggère que les amphipodesH.
azteca se nourrissaient probablement àpriori
sur les sources de nourritures qui étaient disponibles au site oùils
se trouvaient plutôt que d.ese répartir à I'intérieur des lacs en
fonction
d'une source de nourriture particulière.A
l'échelle régionale, en
utilisant
une approche de partitionnementde variation,
nous avonstrouvé
que les communautés d'amphipodesËI
azteca étaient faiblement mais significativement affectéespar les
conditions environnementaleset par
la
structure spatiale.La
compétition interspécifique
et
la
prédationpar les
poissons n'étaient probablement pas importantespour expliquer
la
présenceou
I'absence des espèces dans les lacs dela
région étant donné que les plupart des associations d'espèces ont été classées par un modèle probabiliste comme étant aléatoires.Le
fait
qu'il
y
ait
un niveau élevé de variation qui reste inexpliquée suggère que des procédés comme letri
des espèces et
le
déclin dela
similarité
des communautés enfonction
dela
distancegéographique
ont
probablement
joué
des rôles
modestes
pour
modeler
lescommunautés
de
la
région;
les
facteurs
environnementauxet
les
événements historiques non-structurés spatialementet
la
dérive écologiqueà I'intérieur
des lacsavaient
probablementdes rôles plus
importants.
Dans
I'ensemble,nos
résultats montrent que les espèces d'un complexe d'espèces cryptiques pouvaient avoir à la fois des differenceset
des chevauchements dans leurs niches trophiqueset
dans leurs besoins environnementaux.Cela
suggère que I'associationde
certaines espèces du complexe étaient davantage influencéepar
des procédésliés à
la
niche
écologiquealors
que
les
associations
d'autres
espèces
étaient
probablement
davantage influencées par des procédés liés à la dérive écologique.Mots-clés : Amphipodes, communauté, co-occurrence, détritus organiques, diète, espèces cryptiques, fractionnement isotopique,
Hyalella
azteca, isotopes stables, lac, niche trophiqueKeywords: Amphipods, community, co-occurence, cryptic species, diet,
Hyalella
UNIVERSITÉ DU QUÉBEC À RIMOUSKI
Service de la bibliothèque
Avertissement
La diffusion de ce mémoire ou de cette thèse se fait dans le respect des droits de son auteur,
qui a signé le formulaire « Autorisation de reproduire et de diffuser un rapport, un
mémoire ou une thèse ». En signant ce formulaire, l’auteur concède à l’Université du
Québec à Rimouski une licence non exclusive d’utilisation et de publication de la totalité
ou d’une partie importante de son travail de recherche pour des fins pédagogiques et non
commerciales. Plus précisément, l’auteur autorise l’Université du Québec à Rimouski à
reproduire, diffuser, prêter, distribuer ou vendre des copies de son travail de recherche à des
fins non commerciales sur quelque support que ce soit, y compris l’Internet. Cette licence et
cette autorisation n’entraînent pas une renonciation de la part de l’auteur à ses droits
moraux ni à ses droits de propriété intellectuelle. Sauf entente contraire, l’auteur conserve
la liberté de diffuser et de commercialiser ou non ce travail dont il possède un exemplaire.
CHAPITRE
I
INTRODUCTION GENERALE
1.1
Ecologie des communautésL'un des grands défis de la recherche en écologie est de comprendre comment les espèces s'assemblent en communautés. Traditionnellement, les chercheurs ont mis beaucoup d'effort à étudier les communautés à une échelle surtout locale et à
vérifier
I'influence des processus se déroulant à cette échelle : la relations entre les espèces etleur
environnementmais
égalementles relations entre
les
espècesentre
elles (compétition, prédation, mutualisme, parasitisme)(Ricklefs,
1987;Cottenie
et
al.,2003). Toutefois, depuis
quelques années,nombre de travaux soulignent que
les communautés observées à une échelle locale sont souvent structurées à la fois par des procédés se déroulant à l'échelle locale et par des procédés se produisant à l'échellerégionale,
comme
par
exemple
la
dispersion
d'individus provenant
d'autres communautés présentes dansla
région ou les
échanges de matière organique entre écosystèmes(Holt,
1994; Cottenie et a1.,2003; Polis e/ a1.,2004; Gravel et a1.,2010;Vellend,
2010;Logue
et a1.,2011). Aussi, I'importance des processus stochastiques est maintenantde
plus
en
plus
reconnuepour
expliquer, au moins en partie,
les assemblages d'espèces observés en nature(Gravel
eta1.,2006).
Dans sa théorie del'écologie des communautés,
Vellend
(2010) résume les mécanismes impliqués dansdérive écologique,
la
spéciation etla
dispersion(Fig.
1.1). Les espèces sont ajoutées dansune
communautévia la
spéciationet la
dispersionet
I'abondancerelative
de chaque espèce est ensuitemodifiée par
la
dérive,la
sélectionet
la
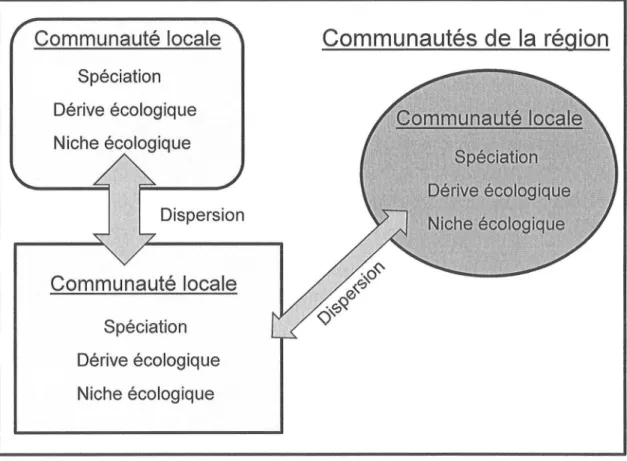
dispersion. Ces mécanismes peuvent agir tant à l'échelle locale (une seule communauté) qu'à l'échelle régionale (métacommunauté) (Vellend, 20 I 0).Figure
l.L
Les quatre mécanismesqui
forment les communautés selonla
théoriede
l'écologie des
communautésde Vellend (2010).
La
spéciation,
la
dérive écologique,la
niche
écologique(ou
sélection selonla
terminologie de
Vellend(2010) et
la
dispersion
vont
interagir
pour
déterminer
la
dynamique
descommunautés
à
plusieurs
échelles spatialeset
temporelles.Figure
adaptée deVellend (2010),
Communauté
loca'le
Spéciation
Dérive écologique
Niche écologique
Communagté
locale
Spéciation
Dérive écologique
Niche écologique
3
Un
mot
sur I'utilisation
du
terme
<
sélection
)
par Vellend.
Le
terme < sélection > désigne habituellementle
fait
que lesindividus
d'unepopulation
ayantdes traits
phénotypiquesdifférents
vont
avoir
des valeurs
adaptatives (/itness)differentes, c'est-à-dire
des
chancesde
survivre
et
de
se
reproduire
differentes(Darwin,
1859).Vellend (2010)
appliquele
conceptde
sélectionà
la
communauté d'espèces essentiellement commeun
synonyme du concept de niche écologique. Enfait,laniche
écologique des espèces est une conséquence de la sélection : la sélection détermine quels traits permettent à des espèces de persister dans unmilieu
et permet éventuellementà
ces espècesde
s'y
adapter(Holt,
2003). Dansla
présente thèse,j'utiliserai
le
terme < niche écologique >plutôt
que(
sélection > étant donné sa plus grande utilisation dans la littérature traitant de l'écologie des communautés.Qu'est-ce qutune communauté?
Le terme
(( communauté>
désigneun
groupeplus ou moins
défini
dans le temps et dans I'espace d'espèces interagissant ensemble dans unmilieu
(Leibold et al., 2004; Holyoak et aL.,2005). On peut distinguer les communautés selon la persistence de I'assemblage des espèces dans le temps. On parle de << coexistence stable >>, aussiappelé simplement < coexistence
)
par
certains auteurs(Leibold et
McPeek,2006;
Siepielskiet a1.,2010),lorsque la
persistence des espèces dansla
communauté estindéfinie
dansle
temps
et ne
montre pas
de
tendanceà long
terme;
elle
résulte nécessairement d'unedifferentiation
de niche écologique entre les espèces (Chesson,l99I;2000).
On parle de < coexistence instable > lorsquela
persistence des espèces dans la communauté n'est pas indéfinie et que des espèces sont vouées à être excluesdu
système à plus ou moinslong
terme; cetype
de coexistence est favorisé lorsqueles
espècesdu
systèmeont
des valeurs
adaptativessimilaires
(Chesson, 2000). Commeil
est souventdiffrcile
à court terme de distinzuer entre coexistence stable et 1.21.3
instable sans connaître les mécanismes ayarfi mené à la formation de la communauté, on
pouna utiliser le
terme(
co-occurrence)
pour
désigner globalementla
situation dans laquelle des espèces sont retrouvées ensemble dans unmilieu,
indépendamment dufait
que leur persistence soit indéfinie ou non (Leibold et McPeek,2006; Siepielski et aL.,2010).Niche écologique
La
niche écologique estun
concept pouvant renvoyer à plusieurs définitions(Silvertown 2004;
Soberôn2007).
D'un
côté,
elle
fait
référenceà
I'ensemble desconditions
environnementaleset
des ressourcesqui
doivent être
présenteset
des besoinsqui
doivent être remplis pour qu'une espèce puisse s'établir et persister dansun milieu
(nichede
Grinnell) (Grinnell,
1917;Hutchinson,
l95l).
À
I'intérieur
de cette perspective, on distingue généralement I'effet des conditions environnementales(par
exemple,la
températureou
le
pH), qui ne
peuventêtre
consomméspar
un organisme maisdont
les valeursvont
déterminersi
une espèce peutvivre
dans unhabitat
ou
non, de
I'effet
d'une ressourcequi
peut
êtreutilisée
et
épuiséepar
les consommateurs (Soberon,2007). Defait,
une ressource conespond à toute substanceou
facteurqui
estutilisé
parun
organismeet
dontla plus
grandedisponibilité
dans I'environnement peut potentiellement augmenter le taux de survie, de croissance etloude
reproduction
de
son
consommateur(Tilman,
1982).Une
ressourcepeut
donc correspondreà
des sources de nutriments,mais
aussi,par
exemple,à
des sites aux conditions environnementales adéquatespour
s'établir ou à I'espace-refuge contre les prédateurs sila
disponibilité
de ces sites peut êtrelimitante
pourla
croissance de lapopulation d'une
espèce.D'un
autre côté,
la
niche
écologiquepeut
aussi
faire référenceau rôle
d'une
espèce,à
I'impact qu'elle
a
sur
son
milieu lors de
son)
deux
conceptionsde
la
niche
vont
depair
et doivent
être considérées comme des aspects dela
< nichetotale
> des espèces.Elles font
référence respectivementà
la réponse(aux
besoins) d'une espèceface
à
son
environnementet à
I'impact
d'une espèce sur son environnement lors de son exploitation des ressources(Leibold,
1995; Chase et Leibold, 2003)(Fig. L2).
N
É.ôl
c)()
5
o
@ @o
É,(b)
Consommation de R1 -z.-t I
tla
,f lot./
13
?
.1,3 . vÉ.vecteur
I
uilnP.rur.
È NRessource
1
(R1)
F'igure
1.2La
niche écologique d'une espèce décrite selon ses besoins (a) et sonimpact
(b)
sur deux ressources substituablesRl
et R2.
Le
graphique(a)
montreI'isocline
de
croissance
nette
nulle
(ICI.IN)
de
I'espèce,
c'est-à-dire
lescombinaisons
de
densitésde
Rl
et R2 pour
lesquelles I'espècea
une
valeur adaptative absoluenulle
alors que lestaux
de natalité et demortalité
sont égaux.La
zonegrise
à droite de I'ICNN
correspondaux
densitésde
Rl
et R2
pour lesquellesla
population
a
une
croissancepositive
et
inversémentpour
la
zone blanche à gauche deI'ICNN.
Le
graphique montre que I'espèce est moins limitéepar
Rl
que parR2
car sa population a besoin d'une densité moindre deRl
pour croître.Le
graphique(b)
représente I'impactper
capita d'une espèce surRl
et R2.Les
flèches grisesmontrent I'impact de
I'espècesur
chacune des ressources defaçon
indépendantealors que
la
flèche
noire
représentela
somme des
deux vecteurset
s'appellele
vecteur d'impact.
Figure
adaptéede Leibold
(1995)
et Chase etLeibold
(2003).Selon
le
principe
de I'exclusion compétitive, dansun
milieu
homogène, des espècesne
peuvent coexister localementsi
elles
occupent desniches
écologiques identiques.Il
est attendu qu'une espèce qui aurait ne serait-ce qu'un léger avantage sur les autres pour accéder aux ressources limitantes devrait à termeproliférer
et exclure les autres dumilieu
dans lequel elle performele mieux
(Gause, 1934;Hardin,
1960;Connell,
1961). Pour coexister defaçon
stable, des espècesdoivent
nécessairementoccuper
des
niches écologiques sufisamment differentes
pour
minimiser
lacompétition entre
elles pour les
ressourceslimitées
du milieu
(Chesson,2000;
Leibold et
McPeek, 2006).
Lorsquedes
espèces coexistentde façon
stable, leurs populations respectives sont capables àpartir
d'une population réduite de réatteindre, en présence des autres espèces de la communauté, les densités de population qu'ellesavaient lorsque
la
communaûé était
à
l'équilibre;
il
s'agit d'une
condition
à
la coexistencequi
s'appellele
< critèred'invasibilité
> (Chesson,I99l;
Chesson,2000;
Siepielski
et
McPeek,
2010).Pour que cela soit
possible,il
faut
que
I'effet
de
lacompétition
interspécifiquesoit moins
important
pour
les populations
de
chaque espèce queI'effet
dela
compétition intraspécifique (Chesson, 2000). Dans une telle situation, cela veut dire que lesindividus
d'une espèce dont la population décroît sont avantagés car le relâchement de la compétition qu'ils subissent favorise leur accès aux ressourceset
donc
leur
chancesde survivre, de grandir
et
de
se reproduire.Afin
d'éviter d'être exclues du
milieu,
des espèces qui compétitionnent peuvent développer des différences écologiquesqui
leur
permettront d'occuper des niches écologiquesdifférentes. Ces differences
écologiques conespondentà
des compromis
qui
les avantagentsur
certains axesde leur niche
écologiques maisqui les
désavantagent nécessairement par rapport à leurs compétiteurs sur d'autres axes (Chesson, 2000).Pour que
des
espècescoexistent
à
l'échelle
régionale,il
suffrt pour
elles7
lesquels chacune performe
le mieux
soient présents(Leibold,
1998; Soberôn,2007).À
l'échelle locale, pour coexister de façon stable, les espèces doivent développer descompromis à
la fois
dansla
réponse qu'ellesont
faceà leur
environnement et dansI'impact
qu'ellesont
sur
ce
dernier
lorsqu'ellesexploitent les
ressources (Leibold, 1998; Chaseet Leibold,
2003).D'un
côté,pour
pouvoir
setrouver
dansun
mêmemilieu, les
espècesdoivent
avoir
des besoins similaires
en
terme
de
conditions environnementales.Toutefois,
pour
que
la
coexistencesoit
possible,elles
doiventavoir
des besoins differents en ressources de façonà
ce que chaque espèceait
unevaleur
adaptativeplus
élevée sous certaines conditions que celle des autres espèces(Leibold,
1995). Par exemple, dansun
système à deux espèces compétitionnant pour deux ressources,la
population d'une première espècedoit
être davantage limitée parla disponibilité
d'une ressourceI
alors que la population d'une deuxième espècedoit
être
davantagelimitée
par
la
disponibilité
d'une ressource2
(Leibold,
1995); la
situation est aussi valide si une espèce est davantage limitée par la disponibilité d'une ressourcealors que
la
deuxième
espèceest
davantagelimitée par
le
risque
de prédation d'un prédateur commun aux deux espèces(Leibold,
1996; 1998; Chase etLeibold.2003).
Alors
que les besoins de chaque espècevont
déterminersi la
coexistence des espèces est possible dans unmilieu,
les impacts de chaque espèce surla
disponibilité des ressources(ou
surla
densité d'un prédateur)vont
déterminer si cette coexistence sera stable(Leibold,
1995; Chaseet
Leibold,
2003). Dansle
casde deux
espècescompétitionnant