Approche écologique et multi-modale pour étudier
l’observation d’actions
Thèse
Sébastien Hétu
Doctorat en psychologie
Philosophiae Doctor(Ph.D.)
Québec, Canada
© Sébastien Hétu, 2014
iii
Résumé
Lors de l‟observation de mouvements, le système perception-action permet le couplage entre les représentations visuelle et motrice du mouvement observé. Ce couplage produit des effets de facilitation motrice qui pourraient être utilisés pour entraîner le système moteur chez différentes populations cliniques. Cette thèse se propose comme un pont entre la recherche fondamentale et l‟utilisation clinique de l‟observation d‟actions. Son objectif principal était de vérifier si les effets produits par l‟observation de mouvements simples pouvaient être reproduits en utilisant des mouvements de la vie courante. Pour ce faire, les effets de l‟observation d‟actions ont été étudiés chez des sujets sains à l‟aide d‟approches tenant compte des intérêts de la réadaptation. Cette thèse devait également fournir des données sur les différences dans la réponse à l‟observation d‟actions entre les individus et entre les méthodes utilisées pour la mesurer.
Les résultats suggèrent que le système perception-action est surtout recruté lors de l‟observation de mouvements peu familiers. Aussi, nos données indiquent que l‟observation de mouvements de la vie courante peut produire des patrons de facilitation anatomiquement congruents avec les mouvements observés. Nos données montrent également que la magnitude et le patron de réponse sont très variables entre les individus. De plus, les effets mesurés peuvent dépendre de la méthode utilisée pour étudier l‟observation d‟actions. Ainsi, il ne semble pas y avoir de relation directe entre les marqueurs comportementaux et physiologiques de la facilitation motrice induite par l‟observation d‟actions.
À l‟instar de ce qui avait été montré lors d‟études utilisant des mouvements simples, l‟observation de mouvements de la vie courante peut produire un patron de facilitation où la représentation du mouvement observé est associée à sa représentation dans le répertoire moteur de l‟observateur. Les importantes différences inter-individuelles dans les effets suggèrent que certains individus répondent plus facilement que d‟autres à l‟observation d‟actions. Par conséquent, identifier quels patients sont susceptibles de bénéficier le plus de l‟observation d‟actions en réadaptation pourrait permettre une utilisation plus efficiente de cette approche. Nous proposons qu‟une étape importante du passage de l‟observation
d‟actions vers la réadaptation sera l‟identification des marqueurs qui peuvent le mieux prédire les améliorations cliniques.
v
Abstract
During action observation, the perception-action system is responsible for the coupling between the visual and motor representations of observed movement. This coupling induces motor priming effects that could be useful for training the motor system of different clinical populations. This thesis main objective was to test if the effect induced by the observation of simple movements could be obtained during the observation of everyday movements. In order to answer this question, the effect of action observation was studied in healthy subjects using approaches in line with the interests of the rehabilitation domain. Furthermore, we aimed at investigating inter-individual differences in the response to action observation and differences pertaining to the method used to measure this response. Results suggest that the perception-action system is recruited to a greater extend by unfamiliar actions. Our data also indicate that observation of everyday movements can produce anatomically congruent facilitation patterns. Importantly, the magnitude and the pattern of response to action observation greatly vary between individuals. Results also indicate that the facilitation effects can vary depending on the method used to study action observation. Indeed, there seem to be no relation between the behavioral and physiological markers of motor facilitation induced by action observation.
Similar to what was shown in studies using simple movements, observing everyday movements can induce a pattern of motor facilitation where the visual representation of the observed movement is associated with its representation in the observer‟s motor repertoire. The important inter-individual differences in the effects produced by action observation suggest that some individuals could respond more easily to action observation than others. Hence, identifying patients for whom rehabilitation using action observation could be the most beneficial could ensure a more efficient use of this approach. An important future step in the clinical use of action observation will be the identification of markers that could best predict the clinical improvements produced with action observation.
vii
Table des matières
Résumé ... iii
Abstract ... v
Table des matières ... vii
Liste des tableaux ... ix
Liste des figures ... xi
Liste des abréviations ... xiii
Avant-Propos ... xvii
Chapitre 1 Introduction ... 1
1.1 Le système perception-action : description ... 2
Figure 1.1. Réponse motrice et visuelle d‟un neurone miroir de préhension. ... 3
Figure 1.2. Analyse de conjonction et système perception-action. ... 7
1.2 Le système perception-action : mesures ... 8
Figure 1.3. Effet spécifique à l‟effecteur produit par l‟observation d‟action. ... 14
Figure 1.4. Facilitation motrice dans une tâche d‟imitation automatique. ... 19
1.3 Le système perception-action : utilisation en clinique ... 20
Figure 1.5. Méta-analyse sur les effets de la thérapie miroir vs. tous les autres types d‟interventions sur les fonctions motrices. ... 22
1.4 Le fossé entre la recherche fondamentale et la clinique ... 23
Figure 1.6. Le pipeline réitératif et bidirectionnel de recherche biomédicale. ... 24
1.5 Objectifs et hypothèses ... 26
Chapitre 2 Variability in the effector-specific pattern of motor facilitation during the observation of everyday actions: implications for the clinical use of action observation ... 31
2.1 Résumé/Abstract ... 32 2.2 Introduction ... 34 2.3 Experimental procedures ... 39 2.4 Results ... 44 2.5 Discussion ... 46 2.6 Conclusion ... 52 2.7 Acknowledgments ... 53
Figure 2.1. Experimental setup used to present visual stimuli, creating the impression that the subject‟s own limb was performing the action. ... 54
Figure 2.2. Schematization of the time sequence during the action observation task. ... 55
Figure 2.3. Average amplitude of the normalized motor evoked potentials (nMEPs) in each condition for each muscle in arbitrary units (AU). ... 56
Figure 2.4. Relation between the effector-specific motor evoked potentials scores (esMEPs) for the biceps-OP (A) and biceps-FDI (B) dyads. ... 57
Chapitre 3 Modulation of brain activity during action observation: Influence of perspective, transitivity and meaningfulness ... 59
3.1 Résumé/Abstract ... 60 3.2 Introduction ... 62 3.3 Methods ... 66 3.4 Results ... 73 3.5 Discussion ... 76 3.6 Conclusion ... 85
3.7 Competing Interests ... 85
3.8 Funding ... 85
Figure 3.1. Timeline of an experimental trial. ... 86
Figure 3.2. Activation sites during action observation, execution and their overlap... 88
Figure 3.3. Action observation parameters estimates results for regions of the perception-action system. ... 90
Chapitre 4 Lack of relationship between behavioral and TMS markers of motor facilitation induced by action observation ... 97
4.1 Résumé/Abstract ... 98 4.2 Introduction ... 100 4.3 Methods... 103 4.4 Results ... 110 4.5 Discussion ... 112 4.6 Conclusion ... 117 4.7 Acknowledgments... 117
Figure 4.1. Illustration depicting the experimental trials for the behavioral and transcranial magnetic stimulation (TMS) tasks. ... 119
Figure 4.2. Group results for the behavioral task... 120
Figure 4.3. Group results for the transcranial magnetic stimulation (TMS) task. ... 121
Figure 4.4. Relation between the behavioral and physiological markers of motor facilitation induced by action observation. ... 123
Supplementary Figure 4.1. Stability of the performance across the two behavioral task sessions. ... 124
Supplementary Figure 4.2. Individual results of motor facilitation. ... 125
Chapitre 5 Discussion ... 127
5.1 Similarités et différences entre les résultats des études utilisant des approches écologiques et celles utilisant des approches plus contrôlées ... 128
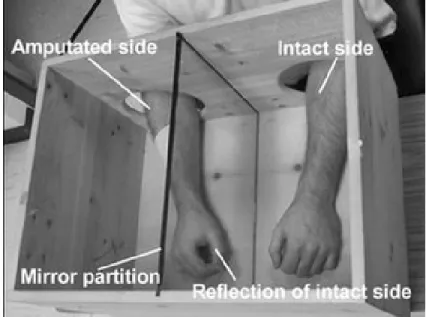
Figure 5.1. Exemple de boîte miroir. ... 132
5.2 Variabilité inter-individuelle ... 139
5.3 Différences inter-méthodes ... 140
Chapitre 6 Limites... 143
Chapitre 7 Conclusion... 147
ix
Liste des tableaux
Chapitre 3
Table 3.1. List of the 6 different everyday movements that were presented to the
participants. ... 91 Table 3.2. List and coordinates of the regions showing greater blood oxygenation
level-dependent signal response obtained from the contrast analyses... 94 Supplemental Table 3.1. Coordinates of the areas resulting from the conjunction analysis.
xi
Liste des figures
Chapitre 1
Figure 1.1. Réponse motrice et visuelle d‟un neurone miroir de préhension. ... 3
Figure 1.2. Analyse de conjonction et système perception-action. ... 7
Figure 1.3. Effet spécifique à l‟effecteur produit par l‟observation d‟action. ... 14
Figure 1.4. Facilitation motrice dans une tâche d‟imitation automatique. ... 19
Figure 1.5. Méta-analyse sur les effets de la thérapie miroir vs. tous les autres types d‟interventions sur les fonctions motrices. ... 22
Figure 1.6. Le pipeline réitératif et bidirectionnel de recherche biomédicale. ... 24
Chapitre 2
Figure 2.1. Experimental setup used to present visual stimuli, creating the impression that the subject‟s own limb was performing the action. ... 54Figure 2.2. Schematization of the time sequence during the action observation task. ... 55
Figure 2.3. Average amplitude of the normalized motor evoked potentials (nMEPs) in each condition for each muscle in arbitrary units (AU). ... 56
Figure 2.4. Relation between the effector-specific motor evoked potentials scores (esMEPs) for the biceps-OP (A) and biceps-FDI (B) dyads. ... 57
Chapitre 3
Figure 3.1. Timeline of an experimental trial. ... 86Figure 3.2. Activation sites during action observation, execution and their overlap. ... 88
Figure 3.3. Action observation parameters estimates results for regions of the perception-action system. ... 90
Chapitre 4
Figure 4.1. Illustration depicting the experimental trials for the behavioral and transcranial magnetic stimulation (TMS) tasks. ... 119 Figure 4.2. Group results for the behavioral task... 120 Figure 4.3. Group results for the transcranial magnetic stimulation (TMS) task. ... 121 Figure 4.4. Relation between the behavioral and physiological markers of motor facilitation induced by action observation. ... 123 Supplementary Figure 4.1. Stability of the performance across the two behavioral task
sessions. ... 124
Chapitre 5
xiii
Liste des abréviations
AC-PC: commissure antérieure-postérieure ANCOVA: analyse de covariance
ADM: abducteur digiti minimi AIP: aire intra-pariétale antérieure ANOVA: analyse de variance AU: unité arbitraire
AVC: accidents vasculaires cérébraux BC: condition congruente biologique BI: condition incongruente biologique
BOLD: signal dépendant du niveau d‟oxygène sanguin ECR: extenseur carpi radialis
EEG: électroencéphalographie EMG: électromyographie EPI: imagerie echo-planaire
esMEPs: scores spécifiques à l‟effecteur de potentiels moteurs évoqués FCR: fléchisseur carpi radialis
FDI: premier dorsal interosseux ICC: corrélation intra-classe
ICF/ICI: facilitation/inhibition intracorticale IFG: gyrus frontal inférieur
IPL: lobule pariétal inférieur
IRMf: imagerie par résonnance magnétique fonctionnelle MEPs: potentiels moteurs évoqués
MEG: magnétoencéphalographie
MI: mouvements intransitifs avec signification MLI: mouvements intransitifs sans signification MT: mouvements transitifs avec signification MVPA: analyse de patrons multi-voxels MT: temps de production du mouvement M1: cortex moteur primaire
nBC: condition congruente non-biologique nBI: condition incongruente non-biologique nMEPs: potentiels moteurs évoqués normalisés OP: opponens pollicis
PMv/PMd: cortex prémoteur dorsal/ventral RMS: racine des carrés moyens
rMT: seuil moteur au repos ROI: régions d‟intérêt RT: temps de réaction
SRC: procédure de compatibilité entre stimulus et réponse SEM: erreur standard de la moyenne
SD: écart type
STS: sulcus temporal supérieur TE: temps d‟écho
TR: temps de réaction
TMS: stimulation magnétique transcrânienne VP: perspective visuelle
xv
Au golden géologue, à la mathématicienne qui voulait voler, à la pianiste, terreur des TUACs et à ma petite rousse. Vous avez été et serez toujours la source et le but de toutes ces petites choses qui forment ma vie.
xvii
Avant-Propos
Si le chemin le plus court entre deux points est la ligne droite, je suis bien content de ne l’avoir pas suivi.
Entre l‟enfant qui rêvait d‟être ingénieur en robotique et l‟homme qui soumet cette thèse en neurosciences se situent de multiples détours, certes pas toujours agréables, mais qui ont su forger une véritable passion pour la recherche. J‟espère que votre lecture saura vous transmettre une partie de cette passion.
Cet ouvrage se présente comme une thèse par articles organisée en sept chapitres. Une introduction générale (chapitre 1) précèdera trois articles scientifiques (chapitres 2, 3 et 4). Ces articles ont été réalisés dans le cadre de mon doctorat sous la direction du Dr. Philip Jackson et du Dre. Catherine Mercier. Deux de ces articles ont déjà été publiés et le troisième a été soumis pour évaluation par mes pairs. En dehors de la mise en page, les articles insérés dans la thèse sont identiques aux articles publiés/soumis. Je suis l‟auteur principal de ces trois articles et j‟ai mené en grande partie la recension des écrits, l‟élaboration des devis de recherche, les expérimentations, l‟analyse des résultats et la rédaction pour ces trois études. Cependant, j‟aimerais souligner l‟important apport des coauteurs dans l‟écriture de ces articles : mes directeurs (qui ont aussi participé à l‟élaboration des projets), ainsi que Martin Gagné, Pierre-Emmanuel Michon, Fanny Eugène, Vincent Taschereau-Dumouchel et Hadj Boumediene Meziane. Leurs commentaires et corrections n‟ont pas seulement permis de bonifier ces articles, elles m‟ont également obligé à toujours chercher à me dépasser et je suis certain que je suis aujourd‟hui un meilleur chercheur grâce à leur usage exhaustif du correcteur Word. Les chapitres 5 et 6 présenteront respectivement une discussion générale et les limites des travaux qui auront été présentés. Une conclusion (chapitre 7) viendra clore la thèse.
J‟aimerais maintenant remercier les personnes qui ont eu la chance de m‟aider dans le cadre de mes études doctorales. Premièrement j‟aimerais souligner l‟immense contribution de mes directeurs à ma formation et à mes succès en recherche. En plus d‟être des mentors extraordinaires, ils auront été d‟incroyables modèles de vie : deux chercheurs qui savent conjuguer succès scientifiques et vie de famille épanouie. Le plaisir associé à ces six
années de discussions et d‟argumentations, de joie et de déceptions, de travail acharné et de fêtes à peine arrosées est en grande partie expliqué par la qualité de la relation que j‟ai avec Catherine et Philip. J‟aimerais également remercier les membres de mes laboratoires pour leurs apports scientifiques et ludiques : Martin Gagné, Joëlle Dubé, Jason Bouffard, Pierre-Olivier Lauzon, Sandra Therrien, Maxime Lee-Vézina, Fanny Jeté, Suzy Ngome, Mélanie Lamothe, Hadj Meziane, Marie-Claude Bilodeau, Guillaume Léonard, Fanny Eugène, Sarah-Maude Deschênes, Rosée Bruneau-Bhérer, Dora-Linsey Canizales, Béatrice Tousignant, Arnaud Saimpont, Marie-Audrey Lavoie, Damien Vistoli, Amélie Achim, Shirley Fecteau, Martin Noël, Jérôme Bibeau, Claire Gane, Midas Millien, Karine Morasse, Karen Reilly, Julien Voisin, Elsa Massicotte, et en particulier l’osti de crew : Mathieu Grégoire, Vincent Taschereau-Dumouchel, Louis-Alexandre Marcoux, Michel-Pierre Coll, et Michel-Pierre-Emmanuel Michon.
Je tiens également à remercier ma mère Monique, mon père Gilles ma petite sœur Marie-Christine et son conjoint Stéphane pour leur appui inconditionnel lors de mes années de purgatoire. Mes remerciements vont également à mes amis Mélissa, Guillaume, et Fred ainsi qu‟à Justine, Raphaëlle et ma filleule Antoinette pour leurs petits sourires. Enfin j‟aimerais dire merci à ma petite princesse rousse Julie pour son amour, son appui, ses encouragements et le fait qu‟elle ait accepté de marier un vulgaire B.A.
J‟ai mené mes travaux doctoraux grâce à l‟aide financière du Centre interdisciplinaire de recherche en réadaptation et intégration sociale (CIRRIS), des Fonds de la recherche en santé du Québec (FRSQ), et des Instituts de recherche en santé du Canada (IRSC).
1
Chapitre 1 Introduction
Pour plusieurs populations cliniques qui présentent des incapacités motrices, l‟entraînement ou la réadaptation basée sur des exercices physiques peut être difficile voire impossible. De nouvelles approches proposent d‟entraîner ces individus en activant indirectement les représentations cérébrales du mouvement par l‟entremise de l‟observation d‟actions. En effet, depuis plus de 15 ans, des travaux tablant sur diverses techniques de neuroimagerie et des tâches comportementales ont permis de démontrer l‟existence d‟un système cérébral responsable de faire le pont entre la perception et l‟exécution de mouvements. Ces travaux ont de plus montré que l‟observation d‟actions facilite la production de mouvements. Parallèlement à ces découvertes, certaines données provenant de la recherche clinique suggèrent que l‟observation de mouvements pourrait être utile en réadaptation, mais le niveau de preuve en ce sens reste limité. La grande disparité entre les tâches étudiées en recherche fondamentale et les approches utilisées dans la recherche et la pratique cliniques pourrait être un des facteurs les plus importants expliquant la lenteur du passage des données fondamentales aux pratiques cliniques. Afin de combler ce fossé, la présente thèse propose d‟étudier les effets physiologiques et comportementaux de l‟observation d‟actions à l‟aide d‟approches qui tiennent compte des intérêts et des objectifs du domaine de la réadaptation. Grâce à une approche combinant plusieurs plateformes de neuroimagerie et des tâches écologiques, cette thèse permettra d‟accroître nos connaissances sur les différents effets de l‟observation d‟actions sur le cerveau tout en facilitant le passage de ces connaissances au monde clinique.
Afin de donner un cadre de référence précis au lecteur, le système cérébral qui permet de faire le lien entre la perception de mouvements et les représentations motrices sera décrit en introduction. En plus de relater la découverte de neurones miroirs (qui seront décrits dans la section suivante) chez le primate, cette section permettra de dresser un portrait du système perception-action chez l‟humain qui permet de coupler la représentation visuelle à la représentation motrice d‟un mouvement. L‟introduction permettra également de présenter certaines approches/tâches qui ont, jusqu‟à ce jour, permis d‟étudier les différents effets de l‟observation d‟actions en recherche fondamentale grâce à la neuroimagerie et aux
paradigmes comportementaux. Il est à noter que cette thèse ne comporte pas de section méthodologique à proprement dite mais que les différentes approches/méthodologies utilisées seront présentées en détail dans les articles inclus dans la thèse. Enfin, l‟utilisation possible de l‟observation d‟actions comme approche thérapeutique sera abordée en mettant l‟accent sur le défi toujours présent d‟assurer un bon maillage entre les données issues de la recherche fondamentale, les études cliniques et la pratique en réadaptation.
1.1 Le système perception-action : description
Il y a près de 15 ans, des neurones déchargeant à la fois lors de la production et de l‟observation d‟une action ont été découverts chez le macaque dans l‟aire F5 du cortex préfrontal (Gallese, Fadiga, Fogassi, & Rizzolatti, 1996; Rizzolatti, Fadiga, Gallese, & Fogassi, 1996). Des études utilisant des enregistrements unicellulaires avaient d‟abord montré que cette région anatomique contenait des neurones qui déchargeaient lors de mouvements distaux et proximaux des membres supérieurs (Rizzolatti et al., 1988; Rizzolatti et al., 1987). D‟autres études avaient également montré que certains neurones de cette région étaient actifs lorsque les macaques observaient des actions produites par les expérimentateurs (di Pellegrino, Fadiga, Fogassi, Gallese, & Rizzolatti, 1992). Les décharges des neurones étaient par exemple associées à l‟observation de mouvements de préhension ou à des mouvements du bras. D‟une façon surprenante, ces neurones qui répondaient lors de l‟observation de mouvements, lorsque stimulés de façon extrinsèque, engendraient fréquemment la production de mouvements périphériques chez l‟animal. C‟est cette propriété d‟être à la fois impliqués dans la production et dans la perception de mouvements qui poussa l‟équipe de Giacomo Rizzolatti à proposer le nom de « neurones miroirs » (Gallese et al., 1996; Rizzolatti et al., 1996) (voir Figure 1.1). Les régions contenant des neurones miroirs chez le macaque ne se limitent pas au cortex préfrontal. Ainsi, des neurones avec propriétés miroirs ont également été identifiés dans le lobe pariétal, notamment dans la région du lobule pariétal inférieur (IPL) (Fogassi et al., 2005; Gallese, Fogassi, Fadiga, & Rizzolatti, 2002; Rozzi, Ferrari, Bonini, Rizzolatti, & Fogassi, 2008).
3 Figure 1.1. Réponse motrice et visuelle d’un neurone miroir de préhension.
Les conditions comportementales sont schématisées et représentées dans la partie supérieure de chaque panneau. Dans la partie inférieure est présentée une série de huit essais consécutifs et l‟histogramme des réponses neuronales relatives. (A) Une plaque avec un morceau de nourriture était présentée au singe, l‟expérimentateur exécutait un mouvement de préhension vers la nourriture puis amenait la plaque et la nourriture vers le singe qui la prenait. Les phases durant lesquelles la nourriture était présentée et amenée vers le singe sont caractérisées par une absence de décharge neuronale. Au contraire, une forte activation était présente durant les mouvements de préhension de l‟expérimentateur et de ceux du singe. (B) Même procédure qu‟en (A), à l‟exception du fait que l‟expérimentateur touchait à la nourriture avec des pinces. Dans cette condition, le neurone ne répondait que lorsque le singe produisait le mouvement de préhension. La ligne verticale indique le moment où l‟expérimentateur touchait la nourriture et le point noir le début de l‟essai. Tiré de Gallese et al. 1996 (Gallese et al., 1996).
Plusieurs éléments ont poussé les chercheurs à suggérer que les neurones miroirs sont impliqués dans un réseau fonctionnel permettant le couplage entre les représentations sensorielles et les représentations motrices utilisées lors de l‟exécution de mouvements.
Cette proposition, qui s‟inspire fortement de la théorie de l‟embodied cognition (Grafton,
2009), suppose que le « système des neurones miroirs » permet à la représentation d‟une
action observée d‟être immédiatement comparée aux représentations présentes dans le répertoire moteur du macaque. Premièrement, l‟aire F5 et le lobule pariétal inférieur, où les neurones miroirs ont été découverts, sont anatomiquement (Rizzolatti & Luppino, 2001; Tanji & Hoshi, 2008; Wise, Boussaoud, Johnson, & Caminiti, 1997) et fonctionnellement connectés (Nelissen et al., 2011). De plus, ce réseau fronto-pariétal contient diverses représentations motrices (Cooke, Taylor, Moore, & Graziano, 2003; Graziano, 2006; Nelissen, Luppino, Vanduffel, Rizzolatti, & Orban, 2005) activées lors de l‟exécution de mouvements du membre supérieur comme celui d‟atteinte ou de préhension (Hoshi & Tanji, 2007; Mutha, Sainburg, & Haaland, 2011). Enfin, des données récentes chez le macaque ont confirmé que lors de l‟observation de mouvements, l‟information visuelle encodée par le sulcus temporal supérieur (STS), une région jouant un rôle important dans le traitement des informations visuelles biologiques (Blake & Shiffrar, 2007), était par la suite relayée aux régions préfrontales et intrapariétales (Nelissen et al., 2011). Ce réseau de régions impliquées lors de la production de mouvements semble donc posséder plusieurs propriétés qui lui permettraient d‟assurer la correspondance entre la perception visuelle d‟une action et les représentations motrices présentes dans le répertoire moteur des primates.
De nombreux travaux se sont intéressés à identifier certaines caractéristiques des mouvements qui pourraient moduler l‟activité des neurones miroirs. Ces caractéristiques incluent la transitivité (c.-à-d., la présence d‟un objet (Gallese et al., 1996; Kraskov, Dancause, Quallo, Shepherd, & Lemon, 2009) ou d‟une cible (Tkach, Reimer, & Hatsopoulos, 2007)), la position dans l‟espace du mouvement par rapport au primate
(Caggiano, Fogassi, Rizzolatti, Thier, & Casile, 2009), le contexte dans lequel l‟action est faite (p.ex., mouvement de préhension pour manger ou pour placer) (Bonini et al., 2010; Fogassi et al., 2005), et la perspective visuelle dans laquelle le mouvement est vu
(Caggiano et al., 2011). Comme il sera présenté dans les sections suivantes, l‟influence de plusieurs de ces caractéristiques a également été étudiée chez l‟homme.
5 Les ressemblances anatomiques et fonctionnelles entre l‟humain et le macaque dans ces régions fronto-pariétales (Geyer, Matelli, Luppino, & Zilles, 2000; Peeters et al., 2009) ont rapidement amené les chercheurs à proposer l‟existence d‟un système similaire chez l‟humain (Rizzolatti & Craighero, 2004). L‟hypothèse actuelle soutient que le système miroir chez l‟humain s‟articule autour des régions du gyrus frontal inférieur (IFG) et/ou du cortex prémoteur dorsal/ventral (PMd/PMv), selon les auteurs, ainsi que des régions de l‟IPL incluant l‟aire intra-pariétale antérieure (AIP) (Fabbri-Destro & Rizzolatti, 2008; Iacoboni, 2009; Rizzolatti & Craighero, 2004; Rizzolatti & Sinigaglia, 2010). Cependant, vue la difficulté d‟utiliser l‟enregistrement unicellulaire chez l‟humain, les évidences de neurones ayant des propriétés miroirs chez l‟homme demeurent extrêmement minces (cependant voir (Mukamel, Ekstrom, Kaplan, Iacoboni, & Fried, 2010)). Chez l‟humain, l‟activité du cerveau est surtout mesurée à l‟aide d‟appareils d‟imagerie par résonnance magnétique fonctionnelle (IRMf). Contrairement à l‟enregistrement unicellulaire qui mesure directement l‟activité d‟un neurone, cette technique mesure indirectement l‟activité cérébrale avec une résolution de quelques millimètres cubes. Le signal mesuré représente donc l‟activité d‟une population de neurones. Les résultats issus des études en IRMf qui ont directement tenté de vérifier la présence de populations de neurones miroirs chez l‟humain restent à ce jour contradictoires. Ces études se basent sur le principe de suppression par répétition (Krekelberg, Boynton, & van Wezel, 2006) qui suppose que la répétition dans le temps d‟un même stimulus produit un phénomène d‟habituation au niveau des neurones sensoriels se traduisant par une diminution dans l‟activité cérébrale mesurée par l‟IRMf. Ainsi, comme proposé par Ilan Dinstein (Dinstein, 2008), si les neurones miroirs réagissent comme les neurones sensoriels, ils devraient s‟adapter lors de la répétition d‟un même mouvement observé; lors de la répétition d‟un même mouvement exécuté; et lorsqu‟un mouvement est perçu puis exécuté ET vice-versa (adaptation cross-modale). La découverte d‟une ou plusieurs régions répondant à cette dernière condition (c.-à-d., une diminution de l‟activité cérébrale montrant une adaptation cross-modale) serait une indication très forte de la présence de neurones miroirs chez l‟humain. En utilisant cette approche, un seul groupe a pu montrer une véritable adaptation cross-modale (observation suivie de l‟exécution et exécution suivie de l‟observation), et cela dans l‟IFG
effet, les autres études qui se sont intéressées à ce sujet n‟ont mesuré aucune adaptation
(Dinstein, Hasson, Rubin, & Heeger, 2007) ou des patrons d‟adaptation cross-modaux dans l‟IPL mais seulement dans un seul sens (p.ex., observation suivie de l‟exécution) (Chong, Cunnington, Williams, Kanwisher, & Mattingley, 2008; Lingnau, Gesierich, & Caramazza, 2009). Il est intéressant de noter que d‟autres approches d‟analyse du signal d‟IRMf qui pourraient potentiellement montrer un patron de réponse « miroir » chez l‟humain comme celle de multivoxel pattern analysis (MVPA) (Haynes & Rees, 2006; Norman, Polyn, Detre, & Haxby, 2006; Tong & Pratte, 2012) ont également produit des résultats contradictoires (Dinstein, Gardner, Jazayeri, & Heeger, 2008; Oosterhof, Wiggett,
Diedrichsen, Tipper, & Downing, 2010). Le manque flagrant de consensus sur la présence
de neurones miroirs chez l‟humain dans les régions fronto-pariétales nous pousse à proposer d‟utiliser plutôt le terme de système « perception-action » qui reflète mieux l‟état actuel des connaissances, c'est-à-dire l‟existence d‟un réseau incluant les régions de l‟IFG et de l‟IPL qui permet de faire le pont entre la perception d‟un mouvement et notre répertoire moteur (Jeannerod, 2001) et qui joue un rôle dans notre capacité à reproduire un mouvement observé lors de l‟imitation (Brass & Heyes, 2005; Iacoboni, 2009), dans le développement des habileté sensori-motrices (Casile, Caggiano, & Ferrari, 2011; Ferrari, Bonini, & Fogassi, 2009) et dans l‟apprentissage moteur par observation (Vogt & Thomaschke, 2007) (voir Figure 1.2b).
7 Figure 1.2. Analyse de conjonction et système perception-action.
A. Les neuroscientifiques ont souvent utilisé l‟imagerie par résonance magnétique fonctionnelle (IRMf) et les analyses de conjonction pour identifier les structures activées lors de l‟observation et de l‟exécution de mouvements. Exemple de ce type d‟analyse où la carte d‟activation de l‟observation est statistiquement superposée à celle de l‟exécution. B. Le système fronto-pariétal perception-action chez l‟humain. Le pôle frontal de ce système est l‟IFG et le pôle pariétal, le lobule pariétal inférieur. Modifié de Hétu & Jackson, 2012
1.2 Le système perception-action : mesures
Plusieurs approches ont permis de démontrer l‟existence d‟un système cortical impliqué dans le couplage entre la perception et la production d‟une action chez l‟humain. C‟est le cas de l‟IRMf, de l‟électro/magnétoencéphalographie (EEG/MEG), de la stimulation magnétique transcrânienne (TMS) et des études comportementales. Ces différentes techniques permettent respectivement d‟étudier l‟influence de l‟observation d‟un mouvement sur l‟activité hémodynamique (IRMf) et électrique (EEG/MEG) du cerveau, sur l‟excitabilité cortico-spinale (TMS) et sur les comportements moteurs. Dans les sous-sections suivantes, les données portant sur les effets de l‟observation d‟actions obtenues grâce à l‟IRMf, à la TMS et aux études comportementales seront présentées.
Pour ce qui est des études basées sur l‟EEG/MEG, elles ont notamment pu montrer que l‟observation d‟actions produisait des changements dans le rythme Mu (~ 10 Hz) (Lepage, Saint-Amour, & Theoret, 2008; Lepage & Theoret, 2006; Muthukumaraswamy & Johnson, 2004; Perry & Bentin, 2009), le rythme Beta (~12-30 Hz) (Muthukumaraswamy & Singh, 2008; Press, Cook, Blakemore, & Kilner, 2011), dans les potentiels somato-sensoriels évoqués (Avikainen, Forss, & Hari, 2002; Rossi et al., 2002) ainsi que dans la réponse somato-sensorielle steady-state produite lors d‟une stimulation mécanique (Voisin et al., 2010). Ces modulations dans l‟activité électrique sont similaires à celles produites lors de l‟exécution de mouvement, suggérant que les deux processus sont intimement reliés. Cependant, puisque l‟EEG ne sera pas utilisé dans le cadre de cette thèse, les résultats issus de travaux en électroencéphalograhie ne seront pas présentés plus en détails. Les lecteurs sont invités à lire la revue de Jaime A. Pineda (Pineda, 2005) pour plus de détails sur le lien entre l‟observation, l‟exécution d‟actions et le rythme Mu.
1.2.1 L’IRMf
L‟IRMf permet la mesure indirecte de l‟activité cérébrale par l‟entremise de l‟activité hémodynamique du cerveau. Cette technique se base sur le lien qui existe entre le débit sanguin cérébral, la demande en énergie et l‟activité neuronale. En effet, l‟activité des
9 neurones nécessite un apport en glucose qui parvient aux cellules nerveuses grâce aux vaisseaux sanguins qui irriguent le cerveau. Puisque la consommation en glucose est parallèle à celle en oxygène, et que l‟on sait que la consommation en oxygène est proportionnelle à l‟activité neuronale, la mesure de la concentration d‟oxygène est utilisée comme un marqueur de l‟activité du cerveau en IRMf (voir (Logothetis, 2002)). L‟IRMf utilise le contraste BOLD (blood oxygen level dependant) dont le principe a été démontré par les travaux de Seiji Ogawa (Ogawa, Lee, Kay, & Tank, 1990; Ogawa, Lee, Nayak, & Glynn, 1990). Ce contraste permet d‟identifier des régions où la concentration relative entre l‟hémoglobine contenant de l‟oxygène et la désoxyhémoglobine varie : une diminution de concentration en désoxyhémoglobine dans une région suggérant une augmentation de l‟activité neuronale dans celle-ci. Notons qu‟une étude utilisant l‟optogénétique (Fenno, Yizhar, & Deisseroth, 2011), une technique permettant de directement contrôler l‟activité de cellules nerveuses à l‟aide de stimulations lumineuses, a récemment démontré la relation entre l‟activité neuronale et le signal BOLD (Lee et al., 2010). Enfin, beaucoup de travaux en IRMf isolent le patron d‟activation relié à leur condition d‟intérêt en contrastant l‟activité hémodynamique durant cette dernière à l‟activité pendant une tâche contrôle.
Depuis les débuts des travaux sur l‟observation d‟actions et le système perception-action, l‟IRMf a été un outil de prédilection pour les chercheurs. En effet, ceux-ci ont su tirer profit de son excellente résolution spatiale afin d‟identifier les substrats neuroanatomiques du système perception-action, en plus de mieux comprendre comment ces différentes régions interagissent (p.ex., (Buccino et al., 2001; Emmorey, Xu, Gannon, Goldin-Meadow, & Braun, 2010; Kilner et al., 2009). Puisque l‟observation d‟actions n‟est pas associée à des manifestations comportementales observables proprement dites, l‟IRMf a aussi permis d‟étudier l‟influence de plusieurs variables sur le fonctionnement du cerveau pendant ce processus cognitif (p.ex., (Calvo-Merino, Grezes, Glaser, Passingham, & Haggard, 2006; Costantini et al., 2005). Ainsi, par l‟étude des changements dans l‟activité cérébrale, nous avons maintenant une meilleure compréhension du fonctionnement du cerveau lorsque l‟on observe une action et de comment cette perception peut être couplée à nos représentations motrices.
Suite à la découverte des neurones miroirs chez le macaque, les neuroscientifiques ont rapidement tenté d‟identifier les régions où se produirait le couplage entre l‟observation et l‟exécution d‟une action chez l‟humain. Si la « véritable » démonstration d‟une réponse miroir chez l‟humain nécessite des paradigmes et des analyses assez poussées qui n‟ont vu le jour que depuis quelques années (voir la section sur la description du système perception-action), les premiers travaux ainsi que plusieurs recherches toujours actuelles ont tenté de décrire le réseau impliqué lors de l‟observation d‟actions et de vérifier si ce dernier incluait des régions associées aux comportements moteurs (p.ex., (Turella, Erb, Grodd, & Castiello, 2009). Dans ce type d‟études, l‟activité hémodynamique des participants est mesurée pendant que ces derniers observent et exécutent un mouvement. Ceci permet d‟obtenir une carte d‟activation (c.-à-d., une carte représentant les régions significativement plus actives que durant la condition contrôle) pour chacun de ces processus. Par la suite, ces cartes peuvent être superposées par des analyses de conjonction visant à identifier quelles régions sont impliquées lors de l‟observation ET de l‟exécution de mouvements (voir Figure 1.2a). De nombreux travaux utilisant cette approche ont confirmé l‟implication des régions fronto-pariétales comme le cortex prémoteur ventral, dorsal, l‟IFG, et l‟IPL (Gazzola & Keysers, 2009; Gazzola, Rizzolatti, Wicker, & Keysers, 2007; Gazzola, van der Worp, et al., 2007; Turella et al., 2009). Les données sur l‟implication de l‟IFG et de l‟IPL issues de différentes études ont récemment été confirmées par des méta-analyses visant à identifier les régions activées de façon constante à travers les différents travaux en IRMf (Caspers, Zilles, Laird, & Eickhoff, 2010;
Molenberghs, Cunnington, & Mattingley, 2012). Comme il a déjà été mentionné, le fait
que des régions où sont inscrites des représentations motrices impliquées lors de la production de mouvement soient également activées lors de l‟observation d‟actions permet d‟envisager qu‟elles pourraient bien être responsables du couplage entre les représentations perceptuelles et motrices du mouvement. De plus, une méta-analyse a également montré que ce réseau fronto-pariétal était aussi activé de façon constante lors de tâches d‟imitation motrice (Caspers et al., 2010), soit des tâches qui nécessitent qu‟une action observée soit transformée en commandes motrices.
En plus de permettre d‟identifier les régions recrutées lors de différentes tâches, l‟IRM peut aussi être utilisée afin d‟étudier la connectivité entre les régions. En effet, diverses
11 approches d‟analyse du signal d‟IRMf permettent d‟inférer sur la connectivité fonctionnelle ou effective à l‟intérieur d‟un réseau en identifiant les régions dont les patrons d‟activité sont corrélés ou cohérents (Ramnani, Behrens, Penny, & Matthews, 2004). Des techniques de tractographie peuvent aussi être utilisées afin d‟étudier les liens physiques (c.-à-d., les réseaux de fibres blanches) et donc la connectivité anatomique entre différentes régions de manière non-invasive (Mukherjee, Berman, Chung, Hess, & Henry, 2008). Relativement récentes, ces techniques n‟ont que peu été utilisées pour étudier le système perception-action. Cependant, les données issues de quelques études suggèrent que les régions fronto-pariétales fonctionnent de concert lors de l‟observation de mouvements
(Emmorey et al., 2010; Molinari et al., 2012) et que ces régions sont anatomiquement reliées (Molinari et al., 2012). Ces résultats soutiennent l‟idée que lors de l‟observation d‟actions, les régions frontales et pariétales ne traitent pas l‟information de façon indépendante mais forment bien un réseau fonctionnel. Il est donc probable que le couplage adéquat entre la perception et l‟exécution d‟un mouvement dépende du bon fonctionnement de chacune de ces régions ainsi que de l‟intégrité des connexions qui les relient.
L‟IRMf a également permis d‟étudier le couplage effectué par le système perception-action en relation avec différentes variables. L‟activité dans les régions du système perception-action peut en effet être modulée de façon top-down (sous contrôle cognitif) et bottom-up (sous contrôle des inputs sensoriels). De nombreuses études en IRMf suggèrent que le focus attentionnel (p.ex., évaluer la douleur exprimée ou l‟amplitude de mouvements faciaux (Budell, Jackson, & Rainville, 2010)) et les instructions (p.ex., observer passivement ou observer pour comprendre le mouvement (Molenberghs, Hayward, Mattingley, & Cunnington, 2012)), la motivation (Cheng, Meltzoff, & Decety, 2007) ainsi que des caractéristiques individuelles des observateurs comme l‟expérience avec le mouvement (p.ex., danseur observant des mouvements de danse (Calvo-Merino, Glaser, Grezes, Passingham, & Haggard, 2005; Calvo-Merino et al., 2006); fumeur observant quelqu‟un qui fume (Wagner, Dal Cin, Sargent, Kelley, & Heatherton, 2011), ou la dominance manuelle (Cabinio et al., 2010; Rocca, Falini, Comi, Scotti, & Filippi, 2008; Willems & Hagoort, 2009) peuvent modifier l‟activité des régions fronto-pariétales lors de l‟observation de mouvements. D‟autres travaux ont permis de mettre en lumière l‟effet sur
l‟activation du système perception-action de plusieurs caractéristiques reliées au mouvement observé (effet bottom-up). Les neuroscientifiques se sont notamment intéressés à la complexité d‟une action (Biagi, Cioni, Fogassi, Guzzetta, & Tosetti, 2009), au membre impliqué (Buccino et al., 2001), à la latéralité de la main (droite vs. gauche) (Shmuelof & Zohary, 2006), à la perspective visuelle (1ère vs. 3ème personne) (Shmuelof & Zohary, 2008; Vingerhoets et al., 2012), au contexte visuel entourant l‟action (Iacoboni et al., 2005; Wurm & Schubotz, 2012), à la transitivité (c.-à-d., la présence d‟un objet) (Agnew, Wise, & Leech, 2012; Corina et al., 2007; Menz, McNamara, Klemen, & Binkofski, 2009) et à la signification du mouvement observé (Knutson, McClellan, & Grafman, 2008; Newman-Norlund, van Schie, van Hoek, Cuijpers, & Bekkering, 2010; Villarreal et al., 2008). L‟observation d‟actions ne se limite pas au couplage entre les informations sensorielles et motrices effectué dans les régions du système perception-action. En effet, lorsqu‟un mouvement est observé, les régions prémotrices et pariétales modulent également l‟activité du cortex moteur primaire (M1) (Feurra et al., 2011; Koch et al., 2010; Lago et al., 2010). Il est en effet connu depuis près de deux décennies que l‟observation d‟action produit une augmentation de l‟excitabilité cortico-spinale (Fadiga, Fogassi, Pavesi, & Rizzolatti, 1995). L‟étude de ce phénomène s‟est surtout faite à l‟aide de la TMS, une technique de neuroimagerie qui permet de mesurer l‟excitabilité du cerveau.
1.2.2 La TMS
La TMS est une technique de stimulation non-invasive où un courant électrique passant dans une bobine génère un champ magnétique de haute intensité qui permet d‟exciter ou d‟inhiber une petite région du cortex située sous la bobine (Hallett, 2007). La plupart des études en TMS profitent du fait que la stimulation de M1 produit des potentiels moteurs évoqués (MEPs; décharges simultanées de plusieurs unités motrices sous contrôle de neurones corticaux de la région stimulée) qui peuvent être enregistrés par des électrodes de surface placées sur le muscle d‟intérêt. Cette réponse motrice directement observable et quantifiable permet entre autres de mesurer l‟excitabilité de la voie cortico-spinale, et donc d‟étudier l‟effet de différentes variables sur l‟activité de M1 et de la moelle épinière. Cette
13 approche a notamment été utilisée afin de montrer comment l‟observation d‟action peut avoir un effet excitateur sur M1, et donc potentiellement faciliter un mouvement.
C‟est au milieu des années 90 que Luciano Fadiga et Giacomo Rizzolatti ont démontré pour la première fois que l‟excitabilité de la voie cortico-spinale était augmentée lors de l‟observation de mouvements chez l‟humain (Fadiga et al., 1995). Ce résultat suggérait que le couplage entre la perception et l‟exécution d‟un mouvement induit également une facilitation motrice. De nombreux laboratoires (voir (Fadiga, Craighero, & Olivier, 2005) pour une revue sur le sujet) ont par la suite répliqué ce résultat en utilisant un paradigme similaire à celui de l‟étude originelle : les participants observent des mouvements des membres supérieurs alors que l‟activité électromyographique (EMG) des muscles des bras et/ou des mains est enregistrée afin de mesurer l‟amplitude des MEPs induits par la TMS. Des résultats, montrant que l‟amplitude du réflexe H n‟est pas influencée par l‟observation d‟actions (Baldissera, Cavallari, Craighero, & Fadiga, 2001; Patuzzo, Fiaschi, & Manganotti, 2003) alors que les phénomènes de facilitation/inhibition intracorticales (ICF/ICI) le sont (Strafella & Paus, 2000), ont permis de démontrer que l‟augmentation d‟excitabilité se produit au moins en partie au niveau cortical dans M1.
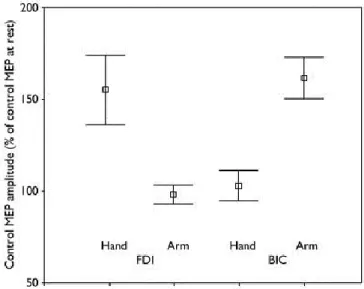
Un des résultats les plus intéressants obtenus à l‟aide de ce paradigme est que la facilitation motrice semble être spécifique à l‟effecteur. Ainsi, l‟observation d‟un mouvement de la main produit une augmentation dans l‟excitabilité de la région contrôlant les muscles de la main, mais pas dans ceux responsables des mouvements du bras et vice-versa (Strafella & Paus, 2000) (Figure 1.3). De nombreux travaux ont aussi pu mettre en évidence qu‟à l‟intérieur même de la représentation de la main, l‟effet de l‟observation d‟actions était spécifique au muscle impliqué dans l‟action (Avenanti, Bolognini, Maravita, & Aglioti, 2007; Catmur, Walsh, & Heyes, 2007; Cavallo, Becchio, Sartori, Bucchioni, & Castiello, 2012; Maeda, Kleiner-Fisman, & Pascual-Leone, 2002; Romani, Cesari, Urgesi, Facchini, & Aglioti, 2005; Sartori, Bucchioni, & Castiello, 2012). Par exemple, on observe de la facilitation motrice pour le muscle premier interosseux dorsal (FDI), qui participe notamment aux mouvements de l‟index, lors de l‟observation de mouvement de l‟index mais pas de l‟auriculaire. Ces résultats suggèrent un couplage fin entre les mouvements observés et les représentations motrices cérébrales. En effet, cette facilitation
anatomiquement congruente dans M1 supporte indirectement les résultats en IRMf de
Giovanni Buccino et ses collaborateurs (Buccino et al., 2001) qui montraient que le système perception-action était somatotopiquement activé lors de l‟observation de mouvements de différentes parties du corps.
Figure 1.3. Effet spécifique à l’effecteur produit par l’observation d’action.
Amplitude des potentiels moteurs évoqués (MEPs), relativement à une condition contrôle, enregistrés pour le premier interosseux dorsal (FDI) et pour le biceps (BIC) pendant l‟observation de mouvements de la main ou du bras. On peut voir que, pour un muscle donné, les MEPs sont plus grands lorsque le mouvement observé implique ce muscle. Tiré
de Strafella et Paus, 2000.
Il a été proposé que l‟expérience sensorimotrice de s‟observer exécuter nos propres mouvements, une activité présente dès notre naissance et qui se poursuit tout au long de notre vie, serait à l‟origine du patron de réponse spécifique à l‟effecteur (Ray & Heyes,
2011). L‟activation simultanée de neurones moteurs et de neurones perceptuels lors de ce
type d‟observation produirait un phénomène d‟apprentissage associatif visuo-moteur
(Heyes, 2001; Heyes & Ray, 2000). Ce pairage anatomique entre la perception et l‟exécution de mouvements se généraliserait par la suite aux mouvements des autres grâce aux expériences socio-culturelles des enfants (Ray & Heyes, 2011). Cette hypothèse suggère que même si le processus est inné (voir (Lepage & Theoret, 2007) pour une discussion sur le sujet), il n‟en demeure pas moins actif puisqu‟il est modelé par l‟expérience. En effet, les travaux de Carolyn Catmur (Catmur, Mars, Rushworth, &
15
Heyes, 2010; Catmur et al., 2007) ont montré que le patron de réponse spécifique à l‟effecteur pouvait être renversé par un apprentissage visuo-moteur incohérent. Ainsi, des participants qui présentaient une facilitation motrice somatotopique (facilitation pour le FDI pendant l‟observation de mouvements de l‟index et pas de l‟auriculaire), présentaient une réponse « contre-miroir » (facilitation pour le FDI pendant l‟observation de mouvements de l‟auriculaire et pas de l‟index) après avoir observé des mouvements de l‟auriculaire alors qu‟ils devaient produire des mouvements de l‟index (Catmur et al., 2007). Cette plasticité dans le patron de facilitation a également été observée lors d‟études s‟intéressant aux changements dans la direction habituelle de mouvements induits par la TMS à la suite d‟expériences motrices, visuelles ou visuo-motrices (Stefan, Classen, Celnik, & Cohen, 2008; Stefan et al., 2005). Si l‟expérience peut modifier le patron de réponse à l‟observation d‟actions, il est fort probable que l‟intensité et la spécificité (à l‟effecteur) de la facilitation motrice produite par l‟observation d‟actions varient entre les individus. Même si très peu de travaux se sont intéressés aux différences individuelles, quelques données semblent confirmer cette hypothèse (Alaerts, Swinnen, & Wenderoth, 2009).
À l‟instar de l‟IRMf, la TMS a été mise à profit pour étudier l‟effet de plusieurs variables sur la facilitation motrice produite par l‟observation d‟actions. Ainsi, la perspective visuelle (Alaerts, Heremans, Swinnen, & Wenderoth, 2009), la latéralité de la main (droite vs. gauche) (Aziz-Zadeh, Maeda, Zaidel, Mazziotta, & Iacoboni, 2002), la race (Desy & Theoret, 2007; Molnar-Szakacs, Wu, Robles, & Iacoboni, 2007) et le sexe (Desy & Theoret, 2007) de la personne observée, la transitivité (Cattaneo, Caruana, Jezzini, & Rizzolatti, 2009; Villiger, Chandrasekharan, & Welsh, 2011), l‟expertise/expérience avec un mouvement (Aglioti, Cesari, Romani, & Urgesi, 2008) ont tous été montrés comme pouvant modifier l‟excitabilité de M1. En considérant les résultats en IRMf et les données issues des travaux en TMS, on peut supposer que le couplage entre la perception de l‟action et les représentations motrices effectué par le système perception-action peut varier en fonction des caractéristiques du mouvement et que cette modulation se répercute sur la facilitation motrice induite par ce couplage.
Les approches de neuroimagerie comme l‟IRMf, la TMS ainsi que l‟EEG ont joué un rôle capital dans la description anatomique et fonctionnelle du système de perception-action. Il n‟en demeure pas moins que ces méthodes sont très coûteuses même pour des centres de recherche. Ainsi, peu de milieux cliniques possèdent l‟équipement nécessaire pour mener ce type de travaux. Si l‟étude plus fondamentale du système perception-action passe inévitablement par ces techniques de neuroimagerie, il est suggéré que l‟utilisation de paradigme comportementaux pourrait être une façon économique d‟étudier le système perception-action (Heyes, 2011). L‟utilisation de tâches comportementales pourrait particulièrement être utile lors de recherches cliniques qui s‟intéressent à la possibilité d‟entraîner les patients par l‟entremise de l‟observation d‟actions mais qui ne disposent pas des ressources nécessaires pour faire de la neuroimagerie. De plus, dans le cadre de programmes de réadaptation, les cliniciens pourraient bénéficier de tâches faciles à implémenter dans leur pratique afin, par exemple, d‟évaluer directement l‟effet de l‟observation d‟actions sur les capacités de leurs patients. Plusieurs études fondamentales qui ont utilisé des tâches comportementales ont permis de confirmer que l‟observation d‟actions induit bel et bien une facilitation motrice (motor priming) et que cette facilitation peut influencer nos comportements moteurs.
1.2.3 Les études comportementales
Nous avons tous déjà vécu un moment où, bien malgré nous, nous imitons les gestes ou les expressions faciales d‟une personne avec qui l‟on interagit. Ce phénomène de mimiquerie motrice a notamment été mesuré en laboratoire (Chartrand & Bargh, 1999). Ainsi, il semble que l‟observation d‟une action puisse avoir un effet sur nos propres comportements, suggérant du même coup un couplage direct entre la perception et l‟exécution de mouvements. Depuis plus de 15 ans, plusieurs approches ont été développées afin de mieux étudier l‟effet de l‟observation de mouvements sur les comportements moteurs.
La plupart de ces approches se basent sur le principe d‟imitation automatique où un individu aura tendance à copier un mouvement observé même si ce dernier n‟est pas relié à
17 la tâche à effectuer (Heyes, 2011). Cette tendance peut interférer avec la performance motrice de l‟individu et peut donc être mesurée (c.-à-d., en termes d‟amélioration ou de diminution). On parle d‟imitation automatique, puisqu‟il semble bien que cet effet se produise hors du champ de conscience de l‟individu (Heyes, 2011; Tipper, 2004). Les études classiques ont utilisé une version modifiée de la procédure de compatibilité entre stimulus et réponse (SRC) (Proctor & Vu, 2006). Ces tâches superposent aux stimuli qui doivent être traités des images photographiques ou vidéo de mouvements qui ne sont pas utiles à la tâche. Ces images/vidéos sont habituellement similaires aux mouvements que les participants doivent produire pour répondre. Par exemple, lors de la tâche les participants doivent identifier le plus rapidement possible la couleur d‟un petit cercle présenté en même temps qu‟une image d‟une main ouverte ou fermée. Les participants reçoivent comme instruction de répondre en fermant la main si le cercle est rouge et de l‟ouvrir s‟il est bleu. Ces études ont ainsi démontré que même si les images de mains n‟étaient pas reliées à la tâche (c.-à-d., la détection de couleur) les temps de réponse étaient plus rapides lors des essais où l‟image de la main était compatible avec la réponse (p.ex., ouvrir la main en voyant une main ouverte) que lorsque qu‟elle ne l‟était pas (p.ex., ouvrir la main en voyant une main fermée) (Sturmer, Aschersleben, & Prinz, 2000). D‟autres études ont plutôt utilisé une procédure modifiée de « go / no-go » où le stimulus qui indique aux participants de produire la réponse (signal go) est un stimulus moteur (p.ex., un doigt qui bouge) semblable à celui que les participants doivent produire pour répondre (p.ex., (Catmur, Walsh, & Heyes, 2009)). Aussi, plusieurs travaux ont utilisé une tâche de temps de réaction (TR) où les participants doivent produire un mouvement de l‟index (ou de l‟auriculaire) lorsqu‟ils voient une cible. Cette cible est superposée à une image/vidéo d‟un mouvement de l‟index ou de l‟auriculaire. Cette procédure engendre des essais compatibles (répondre avec l‟index après l‟observation d‟un mouvement de l‟index) et incompatibles (répondre avec l‟index après l‟observation d‟un mouvement de l‟auriculaire). Les résultats de nombreuses études ont mis en évidence que les essais compatibles étaient associés à des TR plus rapides (Bertenthal, Longo, & Kosobud, 2006; Brass, Bekkering, Wohlschlager, & Prinz, 2000; Catmur & Heyes, 2010; Catmur et al., 2009; Press, Bird, Walsh, & Heyes, 2008; Vogt, Taylor, & Hopkins, 2003) (voir Figure 1.4) et à une exécution plus précise des mouvements (p.ex., mouvements plus rectilignes) (Gowen, Stanley, & Miall, 2008; Kilner,
Paulignan, & Blakemore, 2003; Marshall, Bouquet, Thomas, & Shipley, 2010), ce qui supporte l‟hypothèse que l‟observation d‟actions induit une facilitation motrice. Ainsi, regarder une action recruterait la représentation motrice de cette action, facilitant l‟activation de son patron moteur et conférant un avantage pour la réponse compatible. Il est intéressant de noter que ces résultats sur la compatibilité entre le mouvement observé et la réponse à produire vont de pair avec les travaux en IRMf et en TMS qui suggèrent que l‟observation d‟action produit un effet spécifique à l‟effecteur (p.ex., (Catmur et al., 2008; Strafella & Paus, 2000)).
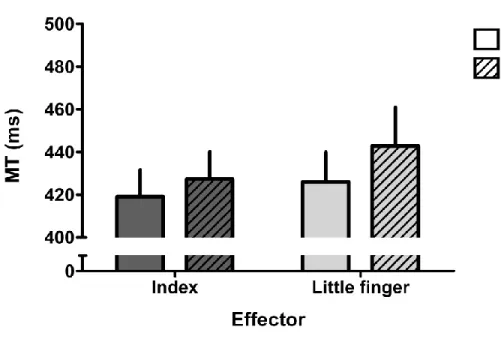
19 Figure 1.4. Facilitation motrice dans une tâche d’imitation automatique.
Temps de réaction (TR; RT dans la figure) lors de conditions où le mouvement observé est congruent (en blanc) ou incongruent (en gris) avec la réponse motrice à produire. On peut remarquer que la condition congruente est associée à des TR significativement plus courts.
Modifié de Catmur et Heyes, 2010 (Catmur & Heyes, 2010).
La concordance entre les résultats sur l‟effet comportemental et neurophysiologique de l‟observation d‟actions appuie la proposition d‟utiliser des tâches comportementales afin d‟évaluer le fonctionnement du système perception-action. En effet, bien que ceci n‟ait jamais été empiriquement confirmé, plusieurs auteurs sont d‟avis que la magnitude des effets comportementaux pourrait être une mesure directe de l‟effet de l‟observation d‟actions sur l‟activation du système perception-action ainsi que de l‟augmentation d‟excitabilité dans M1 (Heyes, 2011; Kilner et al., 2003; Longo, Kosobud, & Bertenthal, 2008; Press, Bird, Flach, & Heyes, 2005; van Schie, van Waterschoot, & Bekkering, 2008). Si tel était le cas, l‟utilisation de ces tâches rapides et peu coûteuses pourrait permettre entre autres aux cliniciens d‟évaluer la « qualité » de la réponse à l‟observation d‟actions de leurs patients : de gros effets comportementaux suggérant une bonne activation du système moteur lors de l‟observation d‟actions. Ceci est important puisque l‟activation du système moteur par l‟observation d‟actions est une technique thérapeutique de plus en plus
employée en réadaptation (Rothgangel, Braun, Beurskens, Seitz, & Wade, 2011; Small, Buccino, & Solodkin, 2012).
1.3 Le système perception-action : utilisation en clinique
Les effets « facilitateurs » de l‟observation d‟actions sur le cerveau et les comportements moteurs ont amené de nombreux chercheurs à vérifier si l‟observation d‟actions pourrait être un outil en réadaptation. Cependant, à ce jour les études cliniques semblent ne s‟appuyer qu‟en partie sur les abondantes données fondamentales déjà obtenues sur le sujet. Ceci s‟explique potentiellement par les différences dans les tâches utilisées en clinique vs. en recherche, qui complique l‟établissement de lien direct entre les évidences fondamentales et les résultats cliniques.
Les problèmes moteurs associés à des conditions cliniques comme les accidents vasculaires cérébraux (AVC) sont habituellement traités lors de séances de réadaptation impliquant de la pratique physique. En effet, l‟activation des réseaux impliqués dans la production de mouvements favoriserait la réorganisation fonctionnelle et le maintien ou la récupération des facultés motrices (Calautti & Baron, 2003; Nudo, 2006). Cependant, cette approche peut parfois être difficile à implémenter, particulièrement en phase aigüe lorsque la fatigue est très présente et que les déficits moteurs limitent de façon importante la production de mouvements. Puisque l‟observation d‟actions active les représentations motrices et induit une facilitation motrice, cette technique pourrait permettre d‟entraîner le système moteur sans que des mouvements aient à être produits. Il n‟est donc pas étonnant que de nombreux chercheurs et cliniciens aient proposé l‟utilisation de l‟observation d‟actions (avec ou sans entraînement moteur concomitant) dans les programmes de réadaptation (Buccino, Solodkin, & Small, 2006; de Vries & Mulder, 2007; Garrison, Winstein, & Aziz-Zadeh, 2010; Iacoboni & Mazziotta, 2007; Pomeroy et al., 2005; Small et al., 2012). De récentes revues de la littérature ont confirmé que l‟utilisation de l‟observation d‟actions pouvait produire des améliorations cliniques (Ezendam, Bongers, & Jannink, 2009; Ramachandran & Altschuler, 2009; Rothgangel et al., 2011; Thieme, Mehrholz, Pohl, Behrens, & Dohle,
21 penchées sur l‟efficacité clinique des approches se basant sur l‟observation d‟actions. Aussi, il est apparent que les caractéristiques des interventions utilisées varient grandement entre les études (voir (Rothgangel et al., 2011)). Il ressort de ceci qu‟il n‟existe pas de directives claires sur les paramètres qui devraient être utilisés lors des interventions (durée, type de mouvements, perspective visuelle, instructions, etc.) (Thieme et al., 2012). Pourtant, une optimisation de ces paramètres devrait permettre d‟obtenir de meilleurs résultats cliniques.
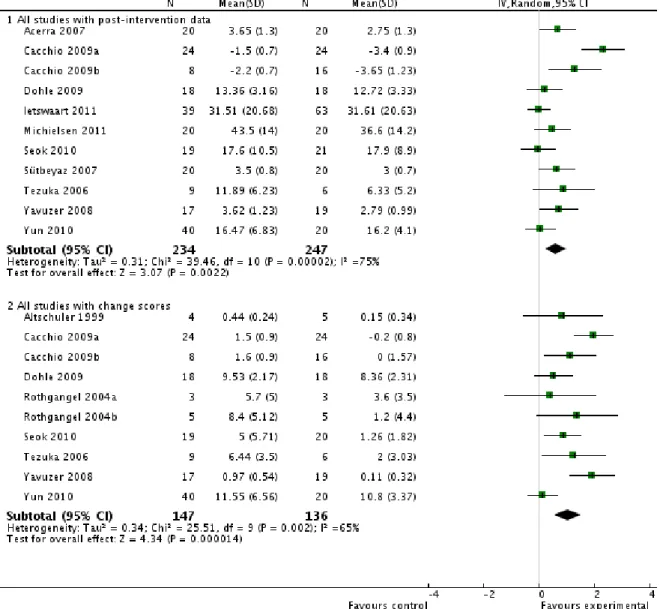
Figure 1.5. Méta-analyse sur les effets de la thérapie miroir vs. tous les autres types d’interventions sur les fonctions motrices.
Taille d‟effet sur les fonctions motrices de la thérapie miroir (un type d‟intervention basée sur l‟observation d‟actions) vs. thérapie contrôle. Partie du haut : études avec des données post-intervention. Partie du bas : études avec données sur score de changement. Modifié de
23
1.4 Le fossé entre la recherche fondamentale et la clinique
Comme présenté dans les sections précédentes, les travaux en recherche fondamentale sur l‟observation d‟actions ont déjà généré une grande quantité de données sur l‟influence de plusieurs variables sur l‟activation du système perception-action et sur la facilitation motrice que ce couplage génère. Le fait que ces résultats semblent ne pas avoir été pleinement pris en compte dans le choix des paramètres à utiliser lors des interventions par les chercheurs cliniques et les professionnels de la santé est un autre exemple de la difficulté à traduire les données fondamentales en pratiques cliniques (translational
research) (Butler, 2008). Les différentes étapes du translational pipeline qui amène les savoirs scientifiques aux pratiques cliniques sont : 1) la recherche fondamentale où les découvertes sont effectuées; 2) les études de démonstration de principes (proof of
principle) où sont testées dans le cadre d‟études à petite échelle des interventions ou
stratégies dérivées des données fondamentales; 3) des études cliniques à grande échelle, et enfin; 4) l‟implémentation d‟interventions basées sur les évidences scientifiques (p.ex., protocoles cliniques, guides de pratique, etc.). Le Cumberland Consensus Working Group a récemment proposé un modèle de pipeline réitératif et bidirectionnel pour le domaine des AVC (Cheeran et al., 2009) (voir Figure 1.6). Ce modèle est dit bidirectionnel puisqu‟il est basé sur la mise en commun des objectifs et des réalités des deux pôles du pipeline. Une des clés de ce modèle est la prise en compte par les chercheurs du pôle fondamental de la nature et des contraintes liées aux activités quotidiennes des représentants du pôle clinique
Figure 1.6. Le pipeline réitératif et bidirectionnel de recherche biomédicale.
Les différentes étapes du passage des données issues de la recherche fondamentale à l‟implémentation de ces connaissances dans la pratique clinique. Les flèches indiquent que les intérêts et objectifs des deux pôles devraient être pris en compte de façon réitérative tout au long du processus. Modifié de Cheeran et al., 2009 (Cheeran et al., 2009).
Cette prise en compte du monde clinique semble n‟avoir que très peu orienté les travaux fondamentaux sur l‟observation d‟actions. Dans le cadre de programmes de réadaptation, les tâches écologiques (impliquant des actions relativement complexes de la vie courante) incluant souvent la manipulation d‟objets sont habituellement utilisées puisqu‟elles ont plus de signification pour les patients et qu‟elles favorisent la performance et l‟apprentissage (French et al., 2007; French et al., 2010; Hétu & Mercier, 2012; Van Peppen et al., 2004). De plus, afin de favoriser la participation des patients, les exercices proposés sont souvent associés à un but (pratiquer cet exercice afin de faire…) (pour l‟observation d‟actions voir (Garrison et al., 2010)). Enfin, les cliniciens sont évidemment plus intéressés aux résultats individuels de leurs patients qu‟aux résultats moyens d‟une population donnée.
Parallèlement à ceci, la recherche fondamentale sur l‟observation d‟actions a principalement utilisé des stimuli très simples, particulièrement des mouvements intransitifs n‟impliquant qu‟une seule articulation. Ces actions ont donc habituellement peu de signification pour les participants (p.ex., la rotation du poignet sans objet n‟a pas de signification propre alors que tourner une poignée de porte en utilisant le même mouvement de poignet est une action familière ayant une signification). Si l‟influence de la transitivité (présence d‟un objet) et de la signification commencent à être étudiées de plus près, les résultats demeurent contradictoires. En effet, certains travaux ont montré que