1
1
UNIVERSITÉ DU QUÉBEC À MONTRÉAL
COMPOSffiON DE LA BANQUE DE SEMENCES LE LONG D'UN GRADIENT DE SUCCESSION AU SUD-OUEST DE LA FORÊT BORÉALE QUÉBECOISE
:MÉMOIRE PRÉSENTÉ
COMME EXIGENCE PARTIELLE DE LA MAÎTRISE EN BIOLOGIE
PAR
CAROLE-LYNE RA TEL
Mise en garde
La bibliothèque du Cégep de l’Témiscamingue et de l’Université du Québec en Abitibi-Témiscamingue a obtenu l’autorisation de l’auteur de ce document afin de diffuser, dans un but non lucratif, une copie de son œuvre dans Depositum, site d’archives numériques, gratuit et accessible à tous.
L’auteur conserve néanmoins ses droits de propriété intellectuelle, dont son droit d’auteur, sur cette œuvre. Il est donc interdit de reproduire ou de publier en totalité ou en partie ce document sans l’autorisation de l’auteur.
1
1
REMERCIEMENTS
Au terme d'un long et fastidieux voyage dans le dédale des études supérieures, je tiens à remercier particulièrement tous ceux de mon entourage qui sont apparus et disparus au cours de ces années de maîtrise. Naturellement, je remercie premièrement ma mère, "celle qui est pas du monde", qui m'a encouragée et empêchée de sombrer dans la "folie" à maintes reprises ainsi que mon frère qui est toujours là pour moi. Ensuite, vient mon directeur Yves Bergeron qui m'a laissé beaucoup de liberté lors de cette expérience. Malgré mes hauts et mes bas, il n'a jamais perdu confiance en moi. Je ne peux oublier ma co-directrice, Francine Tremblay, qui a toujours su alléger l'atmosphère lors de nos diverses réunions.
Je remercie également les gens suivants pour leur support technique en commençant par mes aides de terrain, Stéphane Rhéault et Caroline Chouinard avec qui j'ai passé de bons moments dans le bois de l'Abitibi; Sadek Ahras qui s'est montré très patient à compter avec moi la multitude de petites graines durant les quatre mois d'un été splendide, passés à l'intérieur d'un laboratoire frigorifié et sans fenêtres; Alain l..arouche du département de Géographie de l'Université de Montréal sans qui je n'aurais jamais pu identifier mes graines inconnues; Alain Leduc, le grand manitou des statistiques, qui a été très patient avec moi lors de ses explications.
Je ne saurais terminer ces remerciements sans accorder une attention toute particulière à mes ami(e)s du lOe avec qui j'ai partagé beaucoup et j'ai nommé France, Jacques, Louis et Marty et plusieurs autres que je ne peux énumérer car la liste serait longue. Et également, tous mes amis extérieurs à ce domaine qui, de temps à autre, se sont payé ma tête avec le titre de mon projet
1
TABLE DES .MATIÈRES
REMERCIEMENTS ... ·... .. .. . ... .. .. . ... .. .. u
TABLE DES MA TI ÈRES ... ... ... Ill
LISTE DES FIGURES ... v
LISTE DES TABLEAUX ... VIl RÉSUMÉ . . . .. . . vili CHAPITRE! INTRODUCTION GÉNÉRALE ... 2
1.1 La banque de semences ... 3
1.2 Effets des perturbations sur la banque de semences ... 5
1.3 Objectifs et hypothèses ... 8
CHAPITRE II EFFET DU TEMPS ÉCOULÉ DEPUIS LA DERNIÈRE PERTURBATION SUR LA CO.MpoSmO~ DE LA BANQUE DE SEMENCES AU SUD-OUEST DE LA FORET BOREALE QUEBECOISE ... 10
2.1 Introduction ... 11
2.2 Matériel et méthodes ... 14
2.2. 1 Description du territoire étudié ... 14
2.2.2 Échantillonnage sur le terrain ... 16
2.2.3 Traitements en serre . .. . . . .. . . .. . . . .. . . .. . . . .. . . . .. . . . .. . . 17
2.2.4 Tamisage des sols ... ... ... .... ... .. . . ... 17
2.2.5 Analyses statistiques ... 18
2.3 Résultats ... 19
2.3.2 Effet de la lumière ... 21
2.3.3 Effet du temps écoulé depuis le dernier feu... 21
2.3.4 Dynamique des espèces de la banque de semences persistante... 27
2.3.5 Relation entre les espèces en forêt et dans la banque ... 35
2.4 Discussion ... :... ... 37
2.4.1 Caractéristiques de la banque de semences ... 37
2.4.2 Stratégies des espèces persistantes dans la banque de semences... 40
2.5 Conclusion ... 46
RÉFÉRENCES DE L'ARTICLE ... 47
CONCLUSION GÉNÉRALE ...
55
RÉFÉRENCES .. ... .... . ... ... . ... ... ... . . .. ... .. ... ... ... 57
1
LISTE DES FIGURES
Figure Page
2.1 Carte du Lac Duparquet montrant les transects (en ligne
épaisse) et les différentes aires de feu ... 15
2.2 Nombre moyen de graines germées par échantillon de sol dans les horizons organique et minéral en fonction de deux
intensités lumineuses (100% et 25% de lumière) ... 22
2.3 Nombre total d'espèces herbacées et arbustives avec banques de semences persistante ou transitoire, échantillonnées en serre
en fonction du temps écoulé depuis le dernier feu ... 23
2.4 Nombre total de semis d'espèces herbacées et arbustives provenant de graines et de rhizomes, dénombrés en serre
en fonction du temps écoulé depuis le dernier feu ... 25
2.5 Nombre total d'espèces herbacées et arbustives intolérantes et tolérantes à l'ombre échantillonnées en serre en fonction
du temps écoulé depuis le dernier feu ... 26
2.6 Nombre total de semis d'espèces ·herbacées et arbustives intolérantes et tolérantes à l'ombre provenant de graines,
1
le dernier feu ainsi que les droites de régression dénombrés en serre en fonction du temps écoulé depuisFigure 2.7
2.8
2.9
2.10 2.11 2.121
Fréquences moyennes d'apparition sur le terrain et en serre ainsi que le nombre total de semis et de graines pour
Corydalis sempervirens et Fragaria americana en fonction du temps écoulé depuis le dernier feu ... .
Fréquences moyennes d'apparition sur le terrain et en serre ainsi que le nombre total de semis et de graines pour
Geranium bicknellii et Diervilla lonicera en fonction du temps écoulé depuis le dernier feu ... .
Fréquences moyennes d'apparition sur le terrain et en serre ainsi que le nombre total de semis et de graines pour
Ranunculus abonivus en fonction du temps écoulé depuis le dernier feu ... .
Fréquences moyennes d'apparition sur le terrain e~ en serre ainsi que le nombre total de semis, de graines et de graines viables pour Polygonum cilinode et Sambucus pubens en fonction du temps écoulé depuis le dernier feu ... .
Fréquences moyennes d'apparition sur le terrain et en serre ainsi que le nombre total de semis, de graines et de graines viables pour Prunus pensylvanica en fonction du temps écoulé depuis le dernier feu ... .
Fréquences moyennes d'apparition sur le terrain et en serre ainsi que le nombre total de semis et de graines pour Carex spp. et Rubus idaeus en fonction du temps écoulé depuis le dernier feu ... . V1 Page
29
30 3132
33 341
1
LISTE DES TABLEAUX
Tableau
2.1 Différence des fréquences d'espèces herbacées et arbustives pour l'ensemble des peuplements, calculées à partir de la fréquence moyenne d'apparition sur le terrain (quadrats 1-m2) moins la fréquence moyenne d'apparition en serre (sur 552 échantillons
Page
de sol). On y retrouve également le nombre de graines non-germées retrouvées dans les 376 sols tamisés et la fréquence moyenne ~o~espon~te. I.a. tolérance à l'ombre des espèces
est aussi Indiquee... ... 20
2.2 Coefficients de similarité de Steinhaus entre les fréquences d'apparition sur le terrain et celles dans la banque de semences pour l'ensemble des espèces en fonction du temps écoulé
1
RÉSUl\fÉ
La dynamique de la forêt boréale est contrôlée par des perturbations naturelles. Suite à la destruction de la végétation par un feu ou une épidémie de tordeuse des bourgeons de l'épinette
(Choristoneurafumiferana),
la banque de semences est un des facteurs clés assurant la régénération naturelle. Le but de ce projet est d'évaluer l'effet du type de perturbations (feu ou épidémie d'insectes) et du temps écoulé depuis la dernière perturbation sur la composition de la banque de semences, le long d'un gradient de succession. L'importance des espèces intolérantes à l'ombre dans la banque de semences devrait diminuer plus le temps écoulé depuis le dernier feu est grand. La formation de trouées dans les sites de plus de 123 ans après feu, suite aux épidémies de tordeuse, permettrait le recrutement périodique et l'augmentation successive des espèces tolérantes à l'ombre dans la banque de semences. En milieu contrôlé, nous avons testé l'hypothèse que la germination des espèces mieux adaptées aux petites trouées (tolérantes à l'ombre) serait favorisée sous un traitement de 25% de lumière comparativement à 100% de lumière. Des échantillons de sols ont été prélevés dans huit sites où sont survenus des feux de grande importance entre 1760 et 1964. Ils ont ensuite été mis en serre et soumis à des traitements afin de reproduire l'effet lumineux associé à deux types de perturbations, un feu (100% lumière) et une trouée (25% lumière) sur la germination des graines. Les résultats indiquent que le nombre d'espèces ainsi que l'abondance de celles-ci dans la banque ne subissent pas de variations importantes dans le temps bien que de légères fluctuations entre les feux soient présentes. Aucune espèce, même dans celles qualifiées de tolérantes à l'ombre, ne semble mieux adaptée pour germer sous un couvert où la quantité de lumière est plus faible. Certains patrons attendus dans l'énoncé des hypothèses ne s'observent pas au niveau de la communauté mais plutôt lors de l'étude individuelle de quelques espèces.Geranium bicknellii
est la seule espèce qui, par sa banque de semences, possède une adaptation réelle au feu. L'établissement dans les sites de .Corydalis sempervirens,
Ranunculus abortivus, Diervi/la lonicera
etCarex
spp. est tributaire d'un intervalle très court entre les feux.Polygonum cilinode, Prunus pensy/vanica
etSambucus pubens
colonisent à nouveau les sites après feu grâce à leur maintien prolongé dans la banque ou par la dispersion des graines par les oiseaux et petits mammifères.Rubus idaeus,
une espèce polyvalente, produira une imposante banque de semences, peu importe que le type de perturbation soit un feu, une coupe ou une trouée.1
3:ïV'H~N~~ NOI~::>IlGO'H~NI
1
CHAPITRE I
INTRODUCTION GÉNÉRALE
En Amérique du Nord, les perturbations à grande échelle jouent un rôle majeur dans la détermination de la structure de la forêt et de la composition des espèces (Pickett et White, 1985). Le feu est le plus important facteur contrôlant la dynamique de la forêt boréale (Heinselman, 1981; Johnson, 1979; Wein et MacLean, 1983) et doit être considéré comme un processus écologique normal (Rowe et Scotter, 1973; Wright et Heinselman, 1973). Outre le feu, les épidémies d'insectes (Blais, 1983; Morin, . Laprise et Bergeron, 1993) et autres types d'ouvertures de la canopée provoquées par la chute d'arbres (Oliver, 1981; Poulson et Platt, 1989) sont également des éléments perturbateurs ayant un impact important sur la mosaïque végétale locale.
Les espèces herbacées et arbustives retrouvées en majorité sur le sol en forêt boréale possèdent des caractéristiques morphologiques et reproductrices influençant leur réponse aux perturbations (lngersoll et Wilson, 1990). Généralement, les premières espèces qui s'établiront après une perturbation sont les espèces dites opportunistes. Elles produisent un très grand nombre de graines facilement transportées par le vent et sont intolérantes à l'ombre. D'autres espèces se développent principalement en formant des rhizomes, des rejets de racines ou de souches (régénération végétative). Les espèces arbustives en font généralement partie. Enfin, seulement quelques espèces dépendent d'une réserve de graines dans le sol s'accumulant entre les perturbations (Archibold, 1989). Contrairement aux autres types de forêts, la banque de semences en forêt boréale n'est pas très importante. En effet, plusieurs études effectuées à différentes latitudes montrent une diminution du nombre de graines retrouvées dans le sol plus la latitude s'approche du cercle polaire (Johnson, 1975; Livingston et Allessio, 1968; Olmstead et Curtis, 1947; Oosting et
1
1
1
1
3
Humphreys, 1940). Cette diminution dans la banque touche surtout les espèces herbacées car elles exploitent davantage cette stratégie de reproduction (banque de semences) que les arbres. Une période de croissance plus courte dans les régions plus au nord occasionne cette baisse du nombre de graines. La courte période de croissance sélectionnera les graines qui vont germer rapidement lorsque les conditions sont favorables afin que les semis puissent s'établir avant la fin de l'été. Le temps que la graine résidera dans le sol sera donc influencé par cette nécessité de profiter pleinement de ce court laps de temps (Johnson, 1975).
1. 1 La banque de semences
Une banque de semences est constituée par toute graine viable présente sous ou
à la surface du sol, ou associée à la litière du sol (Roberts, 1981). Elle représente une source continue de graines potentiellement capables de remplacer une plante adulte empêchant ainsi l'extinction de populations végétales (Kalisz, 1991; Roberts, 1991). Généralement, la banque est continuellement enrichie par l'apport aérien de graines (Thompson et Grime, 1979). C'est pourquoi la banque est perçue comme un facteur clé assurant la régénération des communautés après une perturbation (Archibald, 1979; Egler, 1954; Livingston et Allessio, 1968; Marks, 1974; Moore et Wein, 1977) et elle joue un rôle crucial dans la dynamique des populations végétales.
La banque de semences possède une dimension spatiale et temporelle ainsi qu'une distribution verticale et horizontale reflétant les mécanismes de dissémination et mouvements subséquents des graines dans le sol (Hills et Morris, 1992). Thompson et Grime (1979) ont montré que les espèces d'herbacées et d'arbres ont une gamme variée de types de banque de semences, basée principalement sur la stratégie de survie des espèces. La germination, la dormance ainsi que le temps et la durée de production des graines sont les facteurs jouant un rôle dans la classification des banques de semences. Elles sont généralement divisées en deux catégories, transitoire et persistante. Le qualificatif transitoire implique que la production de graines d'une année sera morte ou aura germé avant l'entrée de nouvelles graines dans la banque. On parle donc de graines viables moins d'un an. La catégorie appelée persistante comprend les graines viables plus d'un an et généralement de plus petite taille que celles du type transitoire (Thompson et Grime, 1979). Le cerisier de Pennsylvanie
(Prunus pensylvanica
L.),1
4
une espèce à banque persistante, fait toutefois exception à cette règle, avec ses graines pouvant atteindre 9 mm de diamètre. Les espèces habituellement retrouvées dans la banque de semences sont les espèces pionnières et de début de succession plutôt que les espèces pérennes, caractéristiques des stades plus matures (Harper, 1977; Oosting et Humphreys, 1940). En effet, les espèces pionnières, étant intolérantes à l'ombre, nécessitent pour la germination et la survie, la pleine lumière retrouvée dans les sites récemment perturbés. Elles produisent de grosses quantités de petites graines à longévité élevée contrairement aux espèces de forêt mature qui produisent de très petites quantités de grosses graines à longévité courte (Harper, 1977; Houle et Phillips, 1988; Huston et Smith, 1987). Les graines d'espèces tolérantes à l'ombre sont habituellement plus grosses (Salisbury, 1942). En effet, pour assurer un bon taux de germination en couvert fermé, l'embryon de ces graines nécessite beaucoup plus de "nourriture" (Bierzychudek, 1982). Certaines espèces pionnières, comme Rubus
idaeus
(Granstrôm, 1982) etPrunus pensy/vanica
(Marks, 1974), présentes habituellement dans le sous-bois des jeunes stades de succession, seront cependant retrouvées, à cause de la grande longévité de leurs graines, dans le sol des forêts matures.Thompson et Grime ( 1979) ont divisé les catégories transitoire et persistante en quatre types distincts. Les types I et II sont transitoires tandis que les types III et IV sont persistants. Le type I est composé de graminées annuelles et pérennes retrouvées dans les régions sèches ou perturbées et capables de germination immédiate. Dans le type II, on retrouve les herbacées annuelles et pérennes colonisant les trouées au printemps. Les herbacées annuelles et pérennes germant surtout en automne et qui maintiennent une petite banque de semences persistante font partie du type III. Enfin, le type IV comprend les espèces herbacées et arbustives annuelles et pérennes possédant une importante banque de semences persistante. Cavers (1983) a démontré que la banque de semences d'une espèce peut être du type "transitoire" dans un habitat et du type "persistante" dans un autre.
La dynamique de la banque de semences est influencée par plusieurs facteurs, comprenant l'espèce et l'abondance de la source de graines, la dispersion, la prédation par les petits mammifères, oiseaux, insectes (Harper, 1977; Louda, 1989) et la mortalité des graines par les maladies causées par les champignons et enfin le type et les
1
1
5
conditions du sol (Simpson, Leck et Parker, 1989). Les espèces pionnières spécialisées dans la colonisation de sites récemment perturbés peuvent adopter une stratégie de dispersion soit spatiale ou temporelle. Dans le second cas, on retrouvera les espèces avec une banque de semences contenant des graines caractérisées par une longue période de dormance. Ces graines attendent des conditions favorables à leur germination qui surviendront lors de la perturbation (Fenner, 1985).
Une particularité importante de la stratégie des banques de semences est la période de temps pendant laquelle la graine demeurera viable dans le sol (Fenner, 1985). Des études sur la germination des graines ont montré que la quantité de graines viables tend à diminuer exponentiellement avec le temps des jeunes stades successionnels aux plus vieux (Freud-Williams, Chancellor et Drennan, 1983; Johnson, 1975; Roberts et Feast, 1973; Thompson, 1978). Ce taux de perte de viabilité sera encore plus grand si le sol est sujet à des perturbations (Frank et Safford, 1970; Whipple, 1978).
1 . 2 Effets des perturbations sur la banque de semences
Le feu ainsi que les épidémies de tordeuse des bourgeons de l'épinette sont deux perturbations d'ordre naturel qui donnent à la forêt boréale l'allure d'une mosaïque de stades successionnels différents. Allumés naturellement par la foudre, les feux en forêt boréale sont généralement des feux de cotironne, de très forte intensité, ou bien des feux de surface sévères qui détruisent de vastes étendues de forêt (Heinselman, 1981).
Les épidémies de la tordeuse des bourgeons de l'épinette (
Choristoneura
fwniferana)
sont les plus importantes perturbations causées par des insectes dans l'est de l'Amérique du Nord en raison de leur étendue et leur sévérité. Trois épidémies ont eu lieu au 20e siècle, autour de 1910, 1940 et 1970, et ont ravagé respectivement 10, 25 et 57 millions d'hectares (Blais, 1983). Ces épidémies durent plusieurs années et sont très destructrices. Elles sont, plus particulièrement, associées au stade mature (peuplement de sapins-épinettes) (Blais, 1959).Le type de perturbation agira donc différemment sur le recrutement des espèces de la banque de semences. Un feu détruit la litière accumulée sur le sol forestier, réduit l'épaisseur des horizons organiques de surface (Van Wagner, 1983) et élimine les
1
1
1
6 espèces compétitrices déjà présentes avant la perturbation (Archibold, 1989; Keddy et
al., 1989). Il permet donc le recrutement de toutes les espèces pionnières. Les
ouvertures du couvert forestier provoquées par les épidémies d'insectes permettent à plus de lumière d'atteindre le sol et stimulent la germination des graines (MacLean, 1988). Cependant, seulement le recrutement de certaines espèces sera favorisé sous ces conditions. Souvent, celles-ci seront plutôt des espèces de forêt mature qui se doivent d'être tolérantes à l'ombre (Venable et Brown, 1988). En effet, la quantité de lumière filtrée sous le couvert forestier d'une trouée moyenne est d'environ 25% (Smith, Knapp et Reiners, 1989).
L'intensité de la perturbation influencera la contribution de la banque de semences pour la régénération de la végétation future (Morgan et Neuenschwander, 1988; Pratt, Black et Zamora, 1984). Un feu de faible intensité détruira une grande proportion de graines mais plusieurs graines plus en profondeur dans l'horizon organique pourront recoloniser le site. Et même si tout l'horizon organique est brûlé par un feu de forte intensité, un nombre appréciable de graines présentes dans l'horizon minéral sera capable de germer (Moore et Wein, 1977). Au Québec, la sévérité des épidémies d'insectes créera des trouées plus ou moins grandes dans la forêt. Dans les petites trouées isolées, le faible niveau de lumière atteignant le sol permettra le recrutement des espèces tolérantes à l'ombre présentes dans la banque de semences. Seules les herbacées et arbustes tolérants à l'ombre pourront s'installer sous la canopée (Poulson et Platt, 1989), tandis qu'une grande trouée, avec un niveau de lumière plus élevé, permettra la germination d'un plus grand nombre d'espèces.
L'intervalle de temps entre les perturbations joue aussi un rôle important sur la
régénération par graines dans la plupart des communautés végétales. Dans des environnements sujets à de fréquentes perturbations, la banque de semences permet aux espèces de courte durée de vie et intolérantes à l'ombre de survivre entre les perturbations (Morgan et Neuenschwander, 1988). Tandis que les communautés où les perturbations sont peu fréquentes possèdent une faible quantité de graines viables dans le sol (Frank et Safford, 1970; Whipple, 1978).
Le potentiel de germination des graines sera influencé par plusieurs facteurs physiques. L'influence de la perturbation sur le succès de germination des espèces de la banque de semences semble dépendre surtout de la profondeur des graines dans le
.1
7
sol (Granstrôm, 1986). Les différents horizons (litière, organique, minéral) du sol ne possèdent pas la même dynamique. La majorité des graines de la banque se retrouve dans les premiers centimètres de l'horizon organique (Archibald, 1989; Houle et Payette, 1990; Moore et Wein, 1977). Elles sont exposées aux conditions favorables pour la germination telle que la lumière, la température plus chaude et la présence
d'oxygène e t germent donc plus rapidement Les graines enfouies dans le sol minéral
demeurent viables plus longtemps car elles sont moins sujettes aux prédateurs présents à la surface du sol et aux nombreux pathogènes de l'horizon organique (Houle et Payette, 1990). Pour que la germination s'effectue, les graines doivent être exposées à des conditions environnementales spécifiques qui interagissent souvent entre elles.
Une concentration élevée de
CD2
(Wesson et Wareing, 1967), un manque de lumière(Strickler et Edgerton, 1976) et d'humidité (Grime et Jeffrey, 1965), une basse température (Tubbs, 1965) inhiberont la germination des graines en général.
Pour une composition végétale donnée, les variations dans les conditions du sol tel que la texture, le pH et autres conditions physiques influencent la persistance des
graines de la banque (Archibald, 1989; Schafer et Chilcote, 1970). Le type
géomorphologique (couplet dépôt-drainage) influence la diversité et l'abondance des
espèces herbacées, arbustives et arborescentes de la banque. Les sols argileux
contiennent habituellement plus de graines que les tills car une aération insuffisante et une baisse de la température du sol semblent préserver les graines plus longtemps (Milton, 1944).
Plusieurs études ont déjà démontré que dans les forêts matures et non-perturbées, la composition des espèces présentes dans la banque ne correspond habituellement pas à celle de la végétation au sol (Johnson, 1975; Kellman, 1970; Livingston et Allessio, 1968; Marquis, 1975; Olmsted et Curtis, 1947; Oosting et
Humphreys, 1940; Strickler et Edgerton, 1976). Toutefois, dans des habitats
fréquemment perturbés, elle est généralement similaire (Fenner, 1985). Puis, au fur et à mesure que le peuplement prend de l'âge, elle diverge (Pratt, Black et Zamora, 1984; Whipple, 1978).
1
1
1
1
1
1
8 1. 3 Objectifs et hypothèsesLa banque de semences persistante des espèces herbacées et arbustives sera principalement considérée lors de cette étude car elle représente un mécanisme de survie pour les espèces retrouvées dans des régions sujettes à des perturbations périodiques, comme dans la forêt boréale.
Tel que décrit ci-haut, le type de perturbation, en raison de son influence sur la dimension des trouées, aura vraisemblablement des conséquences sur la composition de la régénération. En effet, la quantité de lumière atteignant le sol de la forêt est différente, que l'on soit dans un site où a eu lieu un feu ou bien dans une trouée. De ce fait, le succès de germination des diverses espèces de graines de la banque le sera également
Plusieurs auteurs ont également démontré que l'âge d'un peuplement forestier influence la densité et la diversité des graines retrouvées dans la banque. Or, il serait intéressant de voir comment une perturbation causée par les épidémies de tordeuse des bourgeons de l'épinette dans les peuplements âgés modifiera la composition de la banque.
Les objectifs de ce projet sont premièrement d'examiner l'effet du temps depuis la dernière perturbation (feu ou épidémie d'insectes) sur la composition de la banque de semences en comparant cette dernière avec la végétation actuelle. De plus, nous comparerons expérimentalement l'effet de l'intensité lumineuse associée à deux types de perturbations, un feu (100% lumière) et une trouée (25% de lumière) sur la germination des graines. Dans un troisième temps, la dynamique de quelques espèces herbacées et arbustives et d'une espèce arborescente possédant une banque de semences persistante sera caractérisée selon le gradient de succession.
Les hypothèses sont les suivantes:
1) Un intervalle de temps plus grand écoulé depuis le dernier feu entraînera une diminution du nombre et de l'abondance des espèces herbacées et arbustives intolérantes à l'ombre dans la banque.
1
1
1
1
1
9 2) La formation de trouées suite à une épidémie de la tordeuse des bourgeons de l'épinette dans les forêts plus susceptibles (plus de 123 ans après feu) permettra le recrutement périodique d'espèces tolérantes à l'ombre qui viendront enrichir la banque de semences.3) La lumière retrouvée dans les trouées (traitement 25% de lumière) favorisera davantage la germination des espèces tolérantes à l'ombre de la banque alors que les espèces intolérantes à l'ombre germeront en plus grand nombre sous le traitement 100% lumière.
4) Plus le temps écoulé depuis le dernier feu est grand, plus la similarité entre la composition végétale dans le sous-bois et celle dans la banque sera faible.
Ces hypothèses permettront de caractériser l'évolution de la banque de semences suivant un gradient de peuplements d'âges différents. De plus, une étude plus détaillée de la dynamique de quelques espèces retrouvées dans la banque de semences permettra de mieux comprendre les réponses spécifiques de chaque espèce le long d'un gradient de succession.
Une bonne compréhension de la dynamique de la banque de semences dans différents peuplements de composition variée peut aider à prédire la tendance successionnelle de ces peuplements lorsqu'une perturbation survient. Elle permet l'identification de peuplements dans lesquels l'approvisionnement en graines pourrait limiter ou même empêcher l'apparition de certaines espèces tout en identifiant les espèces pouvant causer des problèmes de compétition pour les semis d'arbres suite à
une coupe forestière.
Le chapitre IT du mémoire est rédigé sous forme d'article en vue de sa publication dans une revue scientifique. Il sera suivi d'une conclusion générale reprenant les conclusions énoncées dans l'article.
1
1
CHAPITRE II
EFFET DU TEMPS ÉCOULÉ DEPUIS LA DERNIÈRE PERTURBATION SUR LA COI\1POSITION DE LA BANQUE DE SEI\1ENCES AU SUD-OUEST DE LA FORÊT BORÉALE QUÉBÉCOISE
1
,,
,.
1
1
CHAPITRE II 2 .1 IntroductionLorsque la végétation d'un site est modifiée par une perturbation tel qu'un feu, une coupe ou une épidémie d'insectes, une ouverture ou une trouée plus ou moins
grande est créée. À l'intérieur de cette éclaircie, la composition végétale peut être
renouvelée de trois façons: principalement par la germination de graines viables déjà
présentes dans le site perturbé (Egler, 1954), par le développement de rejets végétatifs, et finalement par la germination de graines provenant de l'extérieur du site perturbé (Granstrôm, 1986b; Kellman, 1974; Moore et Wein, 1977).
La régénération de la forêt par la banque de semences dépend considérablement
de la fréquence des perturbations (Grubb, 1976; Miles, 1974; Milthorpe, 1961). Au fur et à mesure que le temps s'écoule depuis la dernière perturbation, la banque de semences varie en quantité et en composition. Les communautés où les perturbations sont peu fréquentes possèdent une faible quantité de graines viables dans le sol
appartenant surtout aux espèces semi-tolérantes et tolérantes à l'ombre (Frank et
Safford, 1970; Rowe, 1983; Whipple, 1978). De Grandpré, Gagnon et Bergeron ( 1993) ont démontré que chez les peuplements plus vieux caractérisés par une
dominance d'Abies ba/samea dans la canopée et une diminution des espèces feuillues,
on observe également une baisse de la richesse, la diversité et l'équitabilité des strates
herbacée et arbustive. Dans les sites fréquemment perturbés, la banque est plus
importante et est composée principalement d'espèces pionnières (Hall et Swaine, 1980;
Hopkins et Graham, 1983; Kellman, 1974; Liew, 1973).
Plusieurs études ont déjà démontré que dans les forêts matures et non-perturbées, la composition des espèces présentes dans la banque ne correspond habituellement pas à celle de la végétation au sol (Johnson, 1975; Kellman, 1970; Livingston et Allessio, 1968; Marquis, 1975; Olmsted et Curtis, 1947; Oosting et
1
1
,f
1
1
1
12fréquemment perturbés, elle est généralement similaire (Fenner, 1985). Puis, au fur et à mesure que le peuplement prend de l'âge, elle diverge (Pratt, Black et Zamora, 1984; Whipple, 1978).
Au sud-est de la forêt boréale canadienne, la structure et la composition des forêts sont régies par le feu et par les épidémies de la tordeuse des bourgeons de l'épinette (Heinselman, 1981; Johnson, 1979; Pickett et White, 1985; Rowe et Scotter, 1973; Wein etMacLean, 1983). Au sud de la forêt boréale, le cycle de feu plus long permet souvent, lors d'une succession naturelle, une transition dans la composition des espèces (Bergeron et Dansereau, 1993). Les jeunes peuplements sont dominés par des feuillus. Les peuplements intermédiaires sont constitués d'un mélange d'espèces feuillues et de conifères suivis par des peuplements âgés dominés par des conifères tels
Abies balsamea et Thuja occidentalis (Bergeron et Dubuc, 1989). Au cours du dernier siècle, trois épidémies de tordeuse des bourgeons de l'épinette se sont produites dans la région étudiée, affectant surtout les peuplements mixtes et de conifères issus de feux anciens. Bergeron et al. (1995) ont démontré que l'impact des épidémies de tordeuse des bourgeons de l'épinette est d'autant plus grand que les peuplements sont établis depuis une longue période après le feu car ces derniers sont constitués de sapins baumiers matures. De plus, ces derniers sont l'hôte préféré de la tordeuse des bourgeons de l'épinette.
Les feux et les épidémies de la tordeuse des bourgeons de l'épinette, ainsi que leurs intensités (Morgan et Neuenschwander, 1988; Pratt, Black et Zamora, 1984), agiront différemment sur le recrutement des espèces de la banque de semences. Un feu détruit la litière accumulée sur le sol forestier, réduit l'épaisseur des horizons organiques de surface (Van Wagner, 1983) et élimine les espèces compétitrices déjà présentes avant la perturbation (Archibold, 1989; Keddy et al., 1989). ll facilite donc la germination des graines du sol et également l'envahissement par des graines de l'extérieur (Hill et Stevens, 1981). Si un feu est de faible intensité, il détruira une grande proportion de graines mais plusieurs graines plus en profondeur dans l'horizon organique pourront recoloniser le site. Même si tout l'horizon organique est brûlé par un feu de forte intensité, un nombre appréciable de graines présentes dans le sol minéral seront en mesure de germer (Moore et Wein, 1977). Quant aux épidémies d'insectes, elles provoquent des ouvertures dans le couvert forestier et augmentent ainsi
1
1
1
,,
1
1
13la quantité de lumière au sol ce qui stimule la germination des graines (MacLean, 1988). Au Québec, la sévérité des épidémies de la tordeuse crée des trouées plus ou moins grandes dans la forêt. Il s'en suit une différence dans la quantité de lumière parvenant au sol. Dans les petites trouées isolées, le faible niveau de lumière atteignant le sol permettra surtout le recrutement des espèces tolérantes à l'ombre présentes dans la banque de semences et ces espèces domineront ensuite la strate herbacée (Poulson et Platt, 1989). Tandis qu'une très grosse trouée, offrant un niveau de lumière plus élevé, favorisera la germination d'une plus grande diversité d'espèces.
Les objectifs de ce projet sont d'examiner l'effet du temps écoulé depuis la dernière perturbation (feu ou épidémie d'insectes) sur la composition de la banque de semences en forêt boréale, le long d'un gradient de succession. Comme la richesse des strates herbacée et arbustive diminue plus la forêt vieillit, nous suggérons premièrement que les espèces intolérantes à l'ombre soient moins nombreuses dans la banque plus le temps écoulé depuis le dernier feu est considérable. Il en sera de même pour la quantité de graines dans la banque. La formation de trouées dans les peuplements âgés de 123 . ans et plus, suite aux épidémies de tordeuse, permettrait le recrutement périodique d'espèces tolérantes à l'ombre et de ce fait, entraînerait une augmentation de l'abondance des espèces dans la banque de semences. En conditions expérimentales, nous avons testé l'hypothèse voulant que certaines espèces soient mieux adaptées à des trouées de petites dimensions. Ainsi, leur germination serait favorisée sous un traitement avec 25% de lumière (Smith, Knapp et Reiners, 1989). Alors que la germination des espèces adaptées au feu et intolérantes à l'ombre serait favorisée sous un traitement pleine lumière. Enfin, la similarité entre la composition végétale du sous-bois et celle de la banque de semences divergera plus le temps augmente depuis le dernier feu. Car bien que les espèces intolérantes à l'ombre se maintiennent dans la banque, elles disparaissent néanmoins du sous-bois à cause du couvert forestier qui se referme.
1
1
1
1
t
1
l '
14 2. 2 l\1atériel et méthodes2. 2. 1 Description du territoire étudié
Les sites d'échantillonnage sont situés dans la région du Lac Duparquet, en Abitibi, dans le nord-ouest du Québec (figure 2.1). Le Lac Duparquet fait partie de la ceinture d'argile du nord de l'Ontario et du Québec, une grande région physiographique qui est née de l'extension maximale des lacs proglaciaires Barlow et Ojibway (Vincent et Hardy, 1977).
La station météorologique la plus proche est située à La Sarre, à 35 km au nord
du Lac Duparquet La température annuelle moyenne est de 0,8"C et les précipitations
totales annuelles s'élèvent à 856,8 mm (calculées pour la période allant de 1961 à
1990). Même si des périodes de gel peuvent survenir durant toute l'année, la période
moyenne annuelle sans gel est de 64 jours (Environnement Canada, 1993). Le Lac
Duparquet, situé à l'extrémité sud de la forêt boréale, est inclus dans l'aire du domaine climacique de la sapinière à bouleau blanc (Richard, 1980), et l'on y observe la
dominance de
Abies balsamea
(L.) Mill.,Picea mariana
(Mill.) B.S.P. etBetu/a
papyrifera
Marsh. avec la présence dePopu/us tremuloides
Michx. etPicea g/auca
(Moench) Voss (Rowe, 1972). Le territoire d'étude est caractérisé par une topographie alternant entre des plaines argileuses et des petites collines rocheuses recouvertes de
tills remaniés par les eaux du Lac Ojibway (Bergeron
et
al., 1983).Le type de végétation varie selon les dépôts de surface et les stades de
succession (Bergeron et Dubuc, 1989). Le type géomorphologique échantillonné dans
le cadre de cette étude est l'argile à drainage modéré. Sur l'argile, les jeunes stades
successionnels (<100 ans) sont dominés par
Populus tremuloides
et les stadesintermédiaires (100-200 ans) par
Abies ba/samea, Betu/a papyrifera
etPicea glauca
tandis qu'en fin de succession (>200 ans), on retrouve
Abies balsamea
et7huja
occidentalis
L (Bergeron, 1991).Le feu (Bergeron, 1991; Dansereau et Bergeron, 1993) et les épidémies de
tordeuse de bourgeons de l'épinette (Bergeron
et
al., 1995; Blais, 1983; Morin, Laprise,.,
.i
1
1
1
1
1
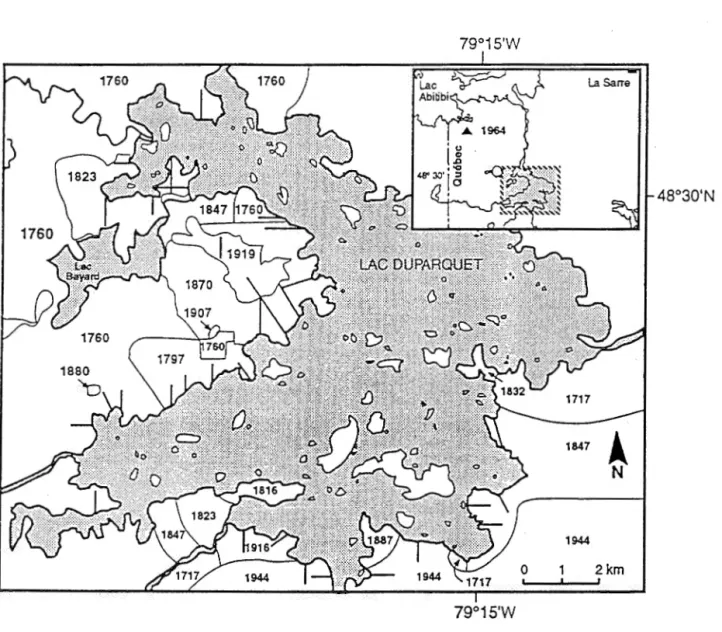
79°15'W 79°15'WFigure 2.1 Carte du Lac Duparquet montrant les transects (en ligne épaisse) et les
différentes aires de feu. Le feu ayant eu lieu en 1964 est localisé à environ dix
kilomètres au nord du lac (voir dans l'encadrement) (Tiré de Bergeron et al., 1995).
15
1
l'
r
.a·
i
1:
16 important sur la mosaïque végétale locale. À l'aide d'un décompte de cicatrices de feux et de l'estimation de l'âge des forêts par une étude dendroécologique, Dansereau et Bergeron (1993) et Bergeron (1991) ont effectué la reconstitution historique et la cartographie des feux dans la région du Lac Duparquet depuis les 300 dernières années. Ils ont établi que le cycle de feu est estimé à 63 ans avant 1870 et plus grand que 99 ans après 1870. Depuis les cent dernières années, trois épidémies de tordeuses des bourgeons de l'épinette ont eu lieu entre 1970 et 1987, entre 1930 et 1950 et entre 1919 et 1929 (Morin, Laprise et Bergeron, 1993). Les forêts utilisées pour fins d'études ont été épargnées par les coupes forestières ou toute autre perturbation anthropique.2. 2. 2 Échantillonnage sur le terrain
De l'ensemble des feux importants survenus dans cette région, huit ont été retenus pour l'étude. Sept ont eu lieu autour du lac (en 1760, 1797, 1823, 184 7, 1870, 1916 et 1944 ). Le plus récent en 1964 se trouve à une dizaine de kilomètres au nord du lac (figure 2.1). Lors d'une étude précédente de Bergeron et al. (1995), la composition et les caractéristiques abiotiques avaient fait l'objet d'un échantillonnage dans 624 quadrats répartis le long de 3 à 4 transects d'environ 1 km, par année de feu. Quatre quadrats de 16m x 16m sur argile à drainage modéré ont été choisis aléatoirement dans chaque année de feu, si possible sur des transects différents. Dix échantillons de sol de 10 cm x 10 cm et 10 cm de profondeur ont été récoltés aléatoirement dans chaque quadrat, pour un total de 320 échantillons. Le prélèvement des sols ayant eu lieu à la fonte des neiges, soit du 18 mai au 3 juin 1993, la banque de semences devait donc exclure toute nouvelle production de graines de l'année (Schiffman et Johnson 1992). Les graines à l'intérieur des sols avaient donc traversé une période de stratification durant l'hiver et étaient prêtes à germer (Whipple 1978). Sur le terrain, lorsque cela était possible, les sols ont été séparés en horizons organique et minéral et déposés à l'intérieur de contenants de 10 cm x 10 cm x 8 cm, pour un total de 640 échantillons (276-organique, 276-minéral, 88-non séparés). Au mois de juillet, des relevés de végétation des strates herbacée et arbustive ont été faits dans dix micro-quadrats de 1m2, à l'endroit même où les échantillons de sol avaient été prélevés. Seulement une espèce arborescente,
Prunus pensylvanica
L., a été incluse dans l'étude1
li
l
1
1
1
_l~
1
1
1
17 et sa fréquence d'apparition provient des relevés de la strate arborescente effectués en1992 (Bergeron
et al.,
1995) pour l'ensemble des quadrats.2. 2. 3 Traitements en serre
Les échantillons de sols récoltés sur le terrain ont été transférés en serre, dans un environnement uniforme et nous avons suivi l'émergence des semis (Kellman, 1970; Oosting et Humphrey, 1940; Strickler et Edgerton, 1976; Whipple, 1978). Les échantillons ont été maintenus en serre du début juin à la fin août à une température moyenne de 25°C, une humidité relative moyenne de 80% et une photopériode moyenne de 15 heures de lumière. Les sols étaient arrosés matin et soir à l'aide d'un brumisateur et ce, pendant une demi-heure. La surface des échantillons de sols a été tournée régulièrement de façon à favoriser la germination des graines plus en profondeur.
Les échantillons ont été répartis aléatoirement dans quatre blocs à l'intérieur de la serre. Ensuite, chaque bloc a été séparé en deux afin de les soumettre aux traitements simulant l'effet lumineux de deux types de perturbation. Un filtre ne laissant passer que 25% de la lumière (env. 496 J.1mole1n21S) a été placé sur la moitié des échantillons de chaque bloc afin de simuler l'effet lumineux d'une trouée petite à moyenne de forêts de conifères (Smith, Knapp et Reiners, 1989). L'autre moitié des échantillons a été exposée à la pleine lumière (env. 2 056 J.lmolefu2fs) afin de simuler l'effet lumineux suite à un feu. Les semis émergeant des sols ont été identifiés à l'espèce lorsque la croissance était assez avancée pour permettre une bonne identification. Une fois identifiés, les semis étaient dénombrés, arrachés hebdomadairement durant les trois mois qu'a duré l'expérience et classés dans les catégories provenant soit de graines ou soit de rhizomes.
2. 2. 4 Tamisage des sols
Du mois de septembre 1993 au mois d'avril 1994, les sols étaient conservés à la noirceur, dans une chambre froide. lls ont ensuite été mis à sécher afm de pouvoir déterminer le poids sec de chacun. Durant l'été 94, 376 échantillons ont été choisis aléatoirement afin de répartir équitablement 12 échantillons/feux pour chaque type
'1
t
1
.,,
t·
;1
1
1
18d'horizon (organique et minéral) et pour chaque traitement lumière ( 100% et 25% ). Les sols ont été tamisés afin de récolter et compter les graines qui n'avaient pas germé. Pour les horizons organiques, seulement le tiers de chaque échantillon a été tamisé et le nombre de graines a ensuite été estimé pour l'échantillon entier. Pour les horizons minéraux, la totalité de l'échantillon a été tamisée. Des tamis en laiton de 20,3 cm de diamètre, avec des mailles de 2 mm, 1 mm, 600 !Jm et 125 J,tm, ont permis de séparer l'échantillon de sol en quatre fractions remplies de graines. Les échantillons étaient préalablement rincés à l'eau. Ensuite, chaque tamis était examiné sous un binoculaire et les graines récoltées à l'aide d'un pinceau humide. Des tests de viabilité au chlorure de tétrazolium (Grabe, 1970) ont été effectués sur Polygonum cilinode, Prunus
pensylvanica
etSambucus pubens
car chez ces espèces, un nombre important de graines était demeuré dans le sol comparativement à la quantité ayant germé.2. 2. 5 Analyses statistiques
Un test de khi-carré a permis de comparer les distributions du nombre d'espèces végétales (persistantes, transitoires, intolérantes et tolérantes à l'ombre) et du nombre de semis provenant de graines à des distributions théoriques et ce, en fonction du temps écoulé depuis le dernier feu. L'abondance dans le temps pour les semis d'espèces intolérantes et tolérantes à l'ombre a été vérifiée par une analyse de régression. Une analyse de variance de rangs (test de Kruskall-Wallis), effectuée avec le logiciel SAS (SAS Institute, 1985), a été utilisée pour comparer le nombre moyen de graines germées en serre pour les deux horizons entre les deux traitements. La fréquence des valeurs de chaque échantillon égales à zéro étant très élevée, les données ont été analysées à l'aide de tests de comparaison non-paramétriques. Ce test a été appliqué sur l'ensemble des espèces ainsi que sur chaque espèce individuellement Les espèces identifiées en forêt mais dont aucune graine n'a germé en serre ont été omises lors des analyses sur la germination. Les fréquences sur le terrain pour chaque espèce herbacée et arbustive, à l'exception de
Prunus pensylvanica,
sont basées sur la présence-absence de l'espèce dans dix micro-quadrats. Elles étaient transformées en pourcentage et ensuite une moyenne des transects par feu était calculée. La fréquence en serre était calculée pareillement mais à partir des échantillons récoltés dans ces mêmes micro-quadrats. Étant une espèce arborescente, la fréquence sur le terrain dePrunus
'1
' ··l,
-a
1
,.
1
t
1
19pensylvanica était indiquée par 100% si présent dans le quadrat et 0% si absent. Une moyenne des transects par feu était ensuite calculée. Les fréquences d'apparition dans la banque et celles sur le terrain pour l'ensemble des espèces dans les huit différents sites ont été comparées à l'aide du coefficient de similarité de Steinhaus (S17) (Legendre et Legendre, 1984).
2. 3 Résultats
Soixante-deux espèces ont été identifiées sur le terrain pour l'ensemble des sites échantillonnés (tableau 2.1). Des semis de seulement 39 espèces ont été dénombrés dans les échantillons de sols mis en serre. Quatre espèces, soit Corydalis sempervirens
(L.) Pers., Fragaria americana (Porter) Britton, Geranium bic/aze/Iii Britton et Sa/ix disco/or Mühl., absentes dans le sous-bois, ont germé en serre.
2. 3.1 Identification des espèces à banque de semences persistante
Le tableau 2.1 dresse la liste des espèces en ordre décroissant suivant la différence entre la fréquence d'apparition sur le terrain et celle en serre. Les espèces retrouvées en bas de liste avec de faibles fréquences et des fréquences négatives indiquent que la fréquence dans la banque est plus importante que celle sur le terrain. La quantité de graines non-germées retrquvées dans le sol pour les mêmes espèces
(Prunus pensylvanica - 448, Sambucus pubens Michx. - 111, Polygonum cilinode
Michx. - 465, Carex spp. - 1737, Diervilla lonicera Mill. - 495 et Rubus idaeus L. - 6 753) confirme leur importance dans la banque de semences. Ranunculus abortivus L. avec seulement quatre graines non-germées fait également partie de ce groupe. Elles sont reconnues pour produire une banque de semences persistante de même que
Geranium biclazellü, Corydalis sempervirens et Fragaria americana (Archibold, 1989; Granstrôm, 1986a; Harrington, 1972; Marks, 1974; Mladenoff, 1990; Whitney, 1986). L'absence de graine dans le sol pour ces trois espèces s'explique par le fait que la totalité des graines a germé en serre. Les espèces suivantes, Sa/ix disco/or, Epilobium glandulosum Lehm., Lactuca serriola L. et Petasites palmatus (Ait.) Gray, même si elles sont situées en bas de liste, sont exclues du groupe car ce sont des espèces à banque de semences transitoire (Archibold, 1989; Brinkman, 1974;
t
.,
1
'1
'1
1
Tableau 2.1. Différence des fr~quences d'esp«es berhac.Xs et arbustives pour l'ensemble des peuplements, calculées à partir de la fréquence moyenne d'apparition sur le terrain (quadrats 1-m~) moins la fréquence moyenne d'apparition en serre (sur 552 échantillons de sol). On retrouve également le nombre de graines non-gcrmées
retrouv~c:s dans les 376 sols tamisés et la fréquence moyenne correspondante. La tolérance à l'ombre des espèces est
aussi indiquée (lorsque connue).
ESPÈCES Ac~r sficatwn Aralia nudicazdis Climonia bor<alis Tri~n.talis borealis Mit<Jia nuda Rubus pubucen.r l.irrtuMa borealis !t.faillTilhonum c.·UfUJlknu Streptopu.r ros~ Astu macrophyllw C ory us corrwta Galium triflorum Viola "PP· lonicera canadmsis Corrws canadmsi.r T ams canatknsis Pyrola asarifolia Galium asprdum Riburrisu Pyro/.a <iliptica Ribes lacwtre DryoprerU spinulosn Athyriumfili:r-J<mint> Dryopt<ris disjuncta Ribes glunduluswn Vibum..,. «lui< Coptis gromlandica Ac~~Ma rubra Rosa acicularis M~nmsia panù:ulata lmpalims capoa.sis Sorbus amt'ricllflll Vacciniwn an gustifolium O.xtlÜS ln()PiltUJ.tl Vibll17lUm trilob< Solidngo canadmsis Dryo,.<ris p/t<gopt<ris A.ln.us rugosa Goody<ra r<pnu Vncrinium myrriUoilhs Tri Ilium cemMm
Acta<a alba Epilobium angustifolium Prunus ~n.ryivanica • Sambucus pub<ns * lurtLlanchier arborra Thalictrwn pW>acens Cil'ro<aai{Ma Borrychium virginianum Cinna latifolia Comus swlorrifrro Poly11onum cilinod< * Pdwùes JXdmUhi.S ÛJctucn serriola Ranuncu.Jus arbonivus * Fragan·o lMJLriclftQ * Coryialis S<mf'"'irrns * Epilobiwn glundulosum Cart!~IPP · * Sali:r disco/or Ckrani~~m bic~llii * DiuviUa lonicera * Rubus idnn<S * Différ'ence de fréquence terrain-banque 53 .9 40.~ 35.6 32.4 32.0 30. 1 29.0 25.1 18.9 17.9 165 16.0 15.8 15.6 14.8 13.8 t3.1 1~.0 11.3 9.1 8.8 7 .8 6.9 6 .4 6 .0 6.0 j.J 5.1 5.1 4.8 4.2 3.t 3.1 :!A 2.4 2.0 1.9 1.7 1.7 1.6 1.1 0.8 0 .7 0.6 0.5 0.4 0.4 0.4 0.3 0.3 0.3 0.3 0.3 0 .0 -0.6 - 1.3 - 1.6 -7.8 -7.9 -7.9 -10.1 -11.3 -11.:\
* Ea.pècm avec banque de graines penis tante
Total -de graines
non-gemu~es dans la banque 75 0 0 0 0 0 0 0 0 0 0 2009 0 0 12 0 0 0 0 0 0 0 0 0 0 0 0 0 0 Il 0 0 0 0 0 0 18 0 0 0 0 0 448 Ill 0 0 0 0 0 46.~ 0 0 4 0 0 0 1737 0 0 495 6753 Fréquence de graines non-germées dans la banaue 10.7 0 0.4 0 0 0 0 0 0 0 0 0 60.2 0 0 1.9 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 .9 0 0 0 0 0 0 1.6 0 0 0 0 0 36.1 10.4 0 0 0 0 0 0 .8 20.3 0 0 0.5 0 0 0 36.5 0 0 17.6 61.5 Tolérance à l'ombre lntol<r:mt Tol<r.mt Tol<r.mt Tol<r.mt Tolér.mt Tol<r.mt Tol<rant lntol(r.mt Tol<r.mt lntol<r:mt Tol<r.mt Tolérant Tolér:mt Tol<r.mt Tol<r.mt Tol<r.mt Tolérant Tolér:mt Tolérant Tolénnt Tolérant Scm.i-toJér.mt à tolér.mt Semi-tolér.mt à tolér.mt Semi·tolér.mt ln toi. à ~<:mi-tolérant lntolér:mt lntolér.mt Tol<r.wt Tol<r.wt Tolérant lntol<r.mt lntolér.mt lntolér:mt Tolér:mt Tolér:mt Scm.i-tolâant à tolénmt intol<r.mt Tol<r.mt lntolér.mt !ntoiU...:.t lntol<r.mt lntolér:mt lntol<r.mt Tol<r.mt lntol<r.mt lntolér.mt Tolérant lntol. à aemi-tolérnnt 20
1
1
1
1
1
&
1
1
21 Harrington, 1972). Pour Salix disco/or, sa présence dans le sol s'explique par une dispersion hâtive des graines car, chez cette espèce, les processus de floraison et de maturation des fruits s'effectuent avant l'apparition des feuilles, dispositions favorables à la dispersion très tôt de graines aigrettées (Marie-Victorin, 1964 ). Les graines d 'Epilobium glandulosum, présentes dans le site, dataient de la production de l'année précédente car il était encore trop tôt au mois de mai pour l'apparition de cette herbacée dans le sous-bois. L'absence de Lactuca serriola et la faible fréquence de Petasitespalma/us (0,3) s'expliquent par des fréquences sur le terrain et dans le sol quasi égales.
2.3.2 Effet de la lumière
Pour l'ensemble des espèces, l'analyse de variance de rang sur le nombre moyen de graines germées par échantillon de sol en fonction de deux intensités lumineuses, ne montre aucune différence significative entre le traitement 100% lumière et celui à 25% de lumière (p=0,1938), que les horizons minéraux et organiques soient pris séparément ou combinés (figure 2.2). De plus, même si le résultat s'avère non-significatif, on peut toutefois remarquer que les échantillons d'horizon organique contiennent un nombre moyen de graines germées légèrement plus élevé que les échantillons d'horizon minéral (figure 2.2). Lorsque les espèces sont analysées individuellement, le pourcentage de germination est significativement plus élevé sous le traitement 100% lumière pour
Geranium bicknellii (p=0,0358), Ranunculus abortivus (p=0,0473) et Sa/ix disco/or (p=0,0001), alors qu'aucune espèce ne germe mieux sous le traitement 25% lumière (P>0,05).
2. 3. 3 Effet du temps depuis le dernier feu
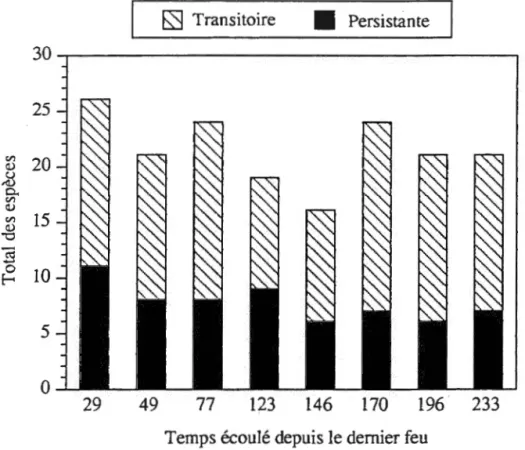
Espèces transitoires vs persistantes
La richesse en espèces est plus élevée dans le site de 29 ans après feu et décroît légèrement jusqu'à celui de 146 ans après feu, passant de 26 à 16 espèces (figure 2.3). La présence en plus grand nombre des espèces transitoires dans les trois plus vieux sites hausse le nombre total d'espèces à 24 dans le site de 170 ans et 21 pour ceux de 196 et 233 ans après feu. Quant aux espèces à banque de semences persistante, la richesse pour chacun des sites varie peu, passant de 11 espèces pour le plus jeune site à
1;
1
1
'
• Organique0
Minéral 12.---~---, 10 8 # mo:yen de grrunes germées 6 par échantillon 4 2 0 100% 25% Intensité lumineuseFigure 2.2. Nombre moyen de graines germées par échantillon de sol dans les horizons organique et minéral en fonction de deux intensités lumineuses (100% et 25% de lumière). Aucune différence significative n'est observée entre les traitements (p=O, 1938).
1
"'
Cl) ~ 0.."'
Cl) "' Cl) -o "'@ ... 0 f-<1
[:;3
Transitoire • Persistante 30~---~25
20
15
10
5
029
49
77
123
146
170
196
233
Temps écoulé depuis le dernier feu
Figure 2.3. Nombre total d'espèces herbacées et arbustives avec banques de semences transitoire et persistante échantillonnées en serre en fonction du temps écoulé depuis le dernier feu.
•
•
'-
.-1 1-•
•
•
•
•
.-•
•
•
•
·--:-::o.•
•
•
•
-24 7, pour le plus vieux. À l'exception du site de 123 ans, les espèces à banque de semences transitoire sont toujours plus nombreuses que celles à banque de semences persistante. Les tests de khi-carré, effectués sur les espèces transitoires, persistantes et sur le total des deux, montrent une répartition non significativement différente à travers les classes pour chacun des cas (p>0,05).Graines vs rhizomes
La quantité la plus élevée de semis provenant de graines est observée en début de succession (figure 2.4). Le test de khi-carré révèle que la quantité de graines dans le site de 29 ans après feu est plus élevée que la valeur prévue par la distribution théorique. Quant à eux, les sites de 77, 146 et 196 ans après feu obtiennent des valeurs au-dessous de celles de la distribution théorique. Le site de 29 ans contient le plus grand nombre de semis provenant de graines, soit 437. Ce nombre décline jusqu'au site intermédiaire de 146 ans où il est le plus bas, avec 34 semis. Dans les sites de 170, 196 et 233 ans après feu, les quantités de semis augmentent respectivement à 200, 101 et 140. À l'exception du site de 146 ans, les semis provenant de graines surpassent toujours en quantité ceux provenant de rhizomes. On peut supposer que les rhizomes ont été endommagés lors de la collecte des échantillons de sol, réduisant ainsi le nombre de semis provenant de rhizomes .
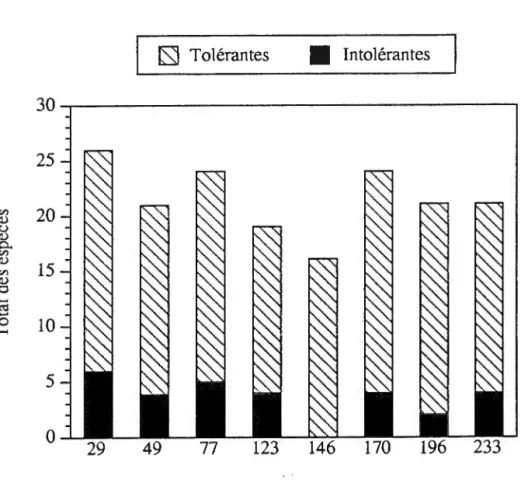
Espèces intolérantes vs tolérantes à l'ombre
À la suite de la répartition des espèces selon leur tolérance à l'ombre (Archibold, 1989; Dansereau, 1957; Harrington, 1972; Rowe et Scotter, 1973; Whitney, 86), on observe que la quantité d'espèces tolérantes à l'ombre surpasse dans les huit sites celle des espèces intolérantes à l'ombre (figure 2.5). Les tests de khi-carré effectués sur les espèces intolérantes et tolérantes à l'ombre ne montrent, dans les deux cas, aucune différence significative (p>0,05) selon une distribution théorique pour chaque classe. En effet, la quantité d'espèces intolérantes dans sept de ces huit classes ne varie qu'entre six et deux tandis que celle des espèces tolérantes varie entre 15 et 20 . Contrairement aux autres sites, la banque de semences du site de 146 ans après feu ne contient aucune espèce intolérante. Les résultats montrant le nombre de semis d'espèces intolérantes et tolérantes à l'ombre indiquent que, peu importe le temps écoulé depuis le dernier feu, les espèces tolérantes à l'ombre se retrouvent en plus
600
500400
Cil"ê
Cl) Cil300
Cl) "'0 "@ 0 E-<200
100
01
1lZl
Rhizomes • Graines1·
29
49
77
123
146
170
196
233
Temps écoulé depuis le dernier feu
Figure 2.4. Nombre total de semis d'espèces herbacées et arbustives provenant de graines et de rhizomes dénombrés en serre en fonction du temps écoulé depuis le dernier feu.
30 25 <Il 20 cu u , Cl) o.. <Il cu <Il 15 cu "'0 "@ ... 0 10 t-< 5 0
1
1
rs'J
Tolérantes • Intolérantes 77 123 146 170 196 233Temps écoulé depuis le dernier feu
Figure 2.5. Nombre total d'espèces herbacées et arbustives tolérantes et intolérantes à l'ombre échantillonnées en serre en fonction du temps écoulé depuis le dernier feu.
1
27 grand nombre dans la banque de semences que les espèces intolérantes (figure 2.6).
Également pour les espèces intolérantes et tolérantes à l'ombre, les droites de régression ne montrent pas une diminution significative du nombre de semis plus le temps écoulé depuis le dernier feu augmente (figure 2.6). Dans le cas des espèces intolérantes, le vieux site de 170 ans après feu contient le nombre le plus élevé de semis, soit 32, se rapprochant ainsi du plus jeune site qui compte 27 semis. Une légère augmentation des semis d'espèces tolérantes est également observée 170 ans après feu mais c'est le plus jeune site qui contient le plus grand nombre de semis, soit 410.
2. 3. 4 Dynamique des espèces de la banque de semences persistante
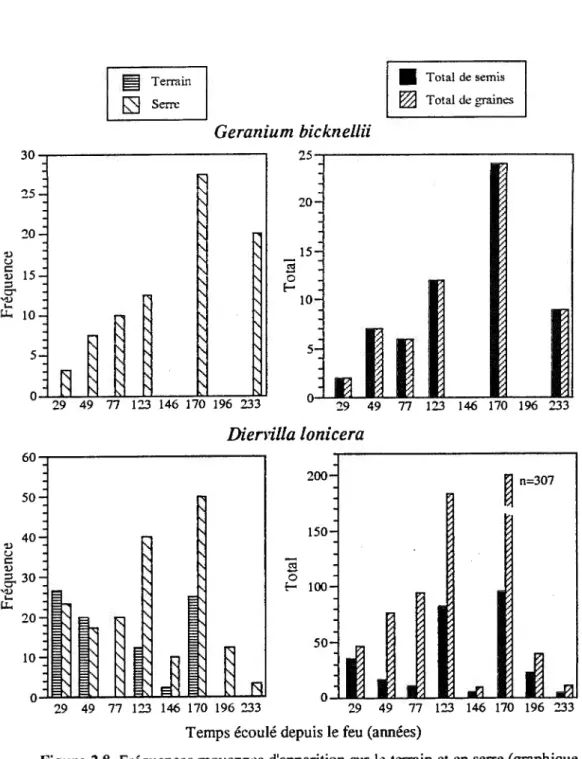
La dynamique en forêt des espèces à banque de semences persistante varie considérablement selon l'espèce. Pour
Coryda/is sempervirens
etFragaria americana
bien qu'elles soient absentes des quadrats échantillonnés, on retrouve néanmoins des graines viables dans la banque de semences (figure 2.7). Dans le cas deGeranium
bicknel/ii,
également absente des quadrats échantillonnés, l'abondance de graines augmente avec le temps écoulé depuis le dernier feu, à l'exception des sites de 146, 196 et 233 ans après feu. Le même phénomène se produit pourDiervil/a lonicera.
(figure 2.8). Les sites d'âge intermédiaire de 123 et 170 ans sont ceux oùDiervil/a lonicera
compte le plus de graines au total ainsi que de semis (graines viables).N'apparaissant qu'en fréquence très faible,
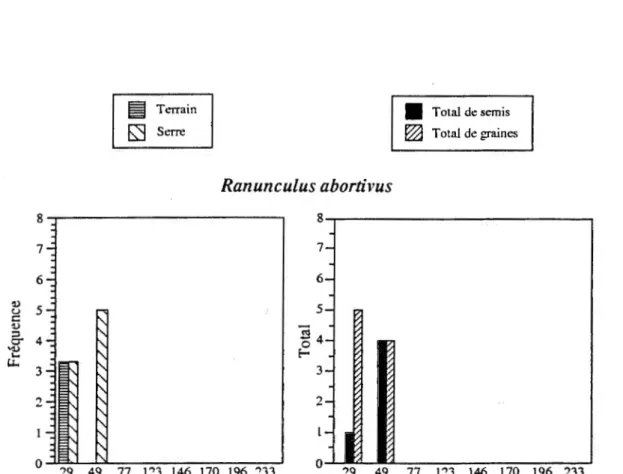
Ranuncu/us abortivus
n'est retrouvée, tant sur le terrain que dans la banque, que dans les deux plus jeunes sites (figure 2.9). La présence dans le sol dePolygonum cilinode
etSambucus pubens
est beaucoup plus importante que leur présence dans le sous-bois (figure 2.10). PourPrunus pensylvanica.,
sa fréquence d'apparition sur le terrain surpasse généralement celle dans le sol (figure 2.11). Cependant, pour les trois espèces, la fréquence dans la banque tend à diminuer dans les sites plus âgés. À l'exception des graines deSambucus pubens
toutes non-viables, une très faible quantité de graines était viable pourPolygonum cilinode
etPrunus pensylvanica.
(figures 2.10, 2.11). Suite une diminution graduelle de la fréquence dans la banque pourCarex
spp. etRubus idaeus
du plus jeune site aux sites intermédiaires, ces mêmes fréquences ont tendance àaugmenter dans les sites de 170 et 233 ans après feu (figure 2.12). Il en est de même pour la fréquence d'apparition sur le terrain, sauf dans le cas de
Carex
spp ..<Il "§ Q) <Il Q) "0 "§ 0 E-< <Il "§ Q) <Il Q) "0 "§ 0 E-<
1
• Intolérantes35
y= -.053757x + 24.874117, r2 = .1328330
p=0,374725
20
15
105
0
~ Tolérantes450
y= -.799299x + 248.835341, r2 = .256794400
p=O,:!OOO350
300
250
200
150
100
50
0
29 49 77 123 146 170 196 233
Temps écoulé depuis le dernier feu
Figure 2.6. Nombre total de semis d'espèces herbacées et arbustives intolérantes et tolérantes à l'ombre provenant de graines, dénombrés en serre en fonction du temps écoulé depuis le dernier feu ainsi que les droites de régression correspondantes.
1
~ Terrain ~ Serre • Total de semis ~ Total de graines Corydalis sempervirens 14-r---, 12- 10-v ~ 8-v ::l C" 'V6-t.t
4- 2-o~~---~~---~ 29 49 77 123 146 170 196 233 20~---, 18- 16- 14- 12-.::1 lO-o f- 8- 6- 4- 2-0iiJ
113 1 29 49 77 123 146 170 · 196 233Fragaria ame ricana
14~---, 14~---, 12- 12-10- 10-tl 8-c: v ::l 8-<13 ë
,B"
...
6- f- 6-tl. 4-4-"-
2-0 29 49 77 123 146 170 196 233 " 29 49 77 123 146 170 196 233Temps écoulé depuis le feu (années)
Figure 2.7. Fréquences moyennes d'apparition ~ur le terrain et en serre (graphique de gauche) ainsi que le nombre total de semis et de graines (graphique de droite) pour Corydalis sempervirens et Fragaria americana en fonction du temps écoulé depuis le feu.
30 25 20 ~ u
5
15 ;:::1 0" '~ 1-o ~ 10 ~ u r::: ~ 5 0 60 50 40g.
30 '~ 1-o [.1., 20 10 0-~
~
~ ~ ~
Geranium bicknellii
-
5 0 r--2 2 1 5 0- 5- 0-5-~
• Total de semislZ'à
Total de graines 29 49 77 123 146 170 196 233 29 49 77 123 146 170 196 233 ~ ~ ~ ~ ~ ~ ~ f' ~'
["'
r-- ["'
r--:,n
'
r--'
r--"
["Dienilla lonicera
~s
0 r--~ n=307 29 49 77 123 146 170 196 233 29 49 77 123 146 170 196 233Temps écoulé depuis le feu (années)
Figure 2.8. Fréquences moyennes d'apparition sur le terrain et en serre (graphique de gauche) ainsi que le nombre total de semis et de graines (graphique de droite) pour Geranium biclazellii et Diervilla lonicera en fonction du temps écoulé depuis le dernier feu.