LES CHANGEMENTS GENOMIQUES ET EVOLUTIFS
DIRIGÉS PAR L'EXPOSITION CHRONIQUE AUX MÉTAUX
CHEZ LA PERCHAUDE {Perça flavescens) SAUVAGE
Mémoire présenté
à la Faculté des études supérieures et postdoctorales de l'Université Laval dans le cadre du programme de maîtrise en biologie
pour l'obtention du grade de Maître es Sciences (M.Sc.)
DEPARTEMENT DE BIOLOGIE FACULTÉ DES SCIENCES ET DE GÉNIE
UNIVERSITÉ LAVAL QUÉBEC
2012
Résumé
La pollution d'origine anthropique peut diriger la microévolution d'adaptations chez les populations d'organismes vivants. Cette étude vise la découverte et la caractérisation de polymorphismes fonctionnels candidats chez des perchaudes (Perça flavescens) chroniquement exposées à la contamination métallique. Au cours d'une première étape, notre méthodologie fait appel à un balayage transcriptomique contrastant les séquences de novo de groupes d'individus provenant de lacs sains (n = 16) et contaminés (n = 16). À partir de ce balayage, 48 marqueurs SNP potentiellement sous sélection par les métaux ont été génotypes et des mesures de bioaccumulation métallique ont été prises chez des perchaudes provenant de 10 populations (n = 1052 individus). L'analyse de ces données a permis de distinguer trois SNPs soumis à la sélection directionnelle et dont les fréquences alléliques sont associées aux niveaux populationnels moyens de contamination selon au moins deux méthodes de régression différentes. De vastes régions génétiques (environ 17kb d'ADN génomique et le mitogénome entier) flanquant ces candidats ont été visées par une expérience de séquençage 454. L'analyse de ces séquences a révélé des substitutions d'acides aminés non-synonymes qui pourraient être adaptatives dans le contexte particulier de la contamination au cadmium. En se basant sur les fonctions des gènes identifiés, nous argumentons que la divergence adaptative entre les perchaudes saines et contaminées suggère que les populations contaminées sont sélectionnées pour la completion rapide de leur cycle vital (voie métabolique de p53, gène de la cycline Gl) et pour la mitigation d'effets délétères au niveau de la mémoire spatio-temporelle (voie métabolique de la potentialisation post-synaptique à long terme; gène de la sous-unité 2 de la NADH déshydrogénase). En accord avec l'attendu évolutif chez les populations stressées et énergiquement limitées, les perchaudes adaptées auraient peu d'aptitude à réparer les dommages causés par les métaux et alloueraient plutôt leur énergie à la croissance. Pour la même raison, l'évolution favoriserait dans ce système des adaptations à faible coût pour la mitigation des impacts néfastes plutôt qu'une stratégie coûteuse de détoxication. Les résultats indiquent que 85 ans de sélection directionnelle imposée par la contamination des métaux a dirigé l'évolution rapide, potentiellement adaptative en sélectionnant des alleles sur des gènes conférant un potentiel adaptatif accru dans les environnements contaminés.
Abstract
Anthropogenic pollution is known to drive rapid evolutionary change in wild animal and plant populations. This study seeks to identify candidate functional polymorphisms of wild yellow perch (Perca flavescens) chronically exposed to metal contamination. We first performed a transcriptome scan from de novo sequencing to contrast a subset of individuals from clean (n = 16) and contaminated (n = 16) lakes. Based on this scan, 48 candidate SNPs located on annotated genes were then genotyped and measurements of metallic bioaccumulation were taken in yellow perch from 10 populations (n = 1052 individuals). Thirdly, a genome scan distinguished three outlier SNPs, one nuclear (located on the cyclin Gl gene) and two mitochondrial (located on the cytochrome b and NADH dehydrogenase subunit 2 genes), with allelic frequencies that correlated to mean population levels of cadmium contamination according to two different regression methods. Whole mtDNA and a 17 kb fragment surrounding the cyclin Gl gene were then sequenced by means of 454 sequencing. This revealed two non-synonymous amino acid (AA) substitutions involving very distinct amino acids. Based on biological functions associated with those genes and the pattern of differentiation between clean and chronically exposed populations, our results suggest that yellow perch from contaminated sites may have been selected for fast life cycle completion (p53 pathway; cyclin Gl gene) and for mitigation of deleterious impacts on temporal and spatial memorization (long-term potentiation pathway; NADH dehydrogenase 2 gene). In accordance with the evolutionary trajectory expectations for stressed and energy deprived organisms, yellow perch adapted to cadmium pollution would not compensate for the shut down of their repair mechanisms to instead reallocate energy towards growth. Similarly, selection may also have favoured inexpensive impairment mitigation adaptations over costly detoxification strategies. Overall, these results indicate that 85 years of directional selection imposed by metal contamination drove the rapid, potentially adaptive evolution of perch populations by selecting alleles at genes conferring increased fitness in polluted environments.
Je dépose aujourd'hui, avec une immense fierté, ce mémoire dont les pages ne peuvent exprimer l'expérience humaine sous-jacente. La nature stérile de la littérature scientifique nous fait souvent oublier les êtres humains qui se cachent derrière celle-ci. Je souhaite que tout lecteur soit conscient que chacune des pages de ce manuscrit est autant teintée de la sueur des efforts soutenus que des éclats de rire les ayant ponctués. Je considère que je sors grandi de cette expérience et je me sens plus apte que jamais à combattre avec optimisme les injustices du monde.
Mes remerciements les plus sincères reviennent à Louis Bernatchez, Serge Higgins et Nicolas Derome qui m'ont successivement donné ma chance dans le milieu de la recherche alors que, fraîchement diplômé du baccalauréat, j'avais bien peu d'expérience à offrir. La confiance et la latitude qu'ils m'ont accordé m'ont été particulièrement précieuses et je ne conserve que des souvenirs agréables de nos collaborations professionnelles.
Les différents postes que j'ai successivement occupé à l'IBIS ont éventuellement attisé une passion pour la génétique qui m'a mené à entreprendre une maîtrise avec Louis Bernatchez pour lequel j'ai le plus grand respect intellectuel. Cette aventure n'aurait pas non plus été possible sans la codirection de Patrice Couture dont l'optimisme et la bonne humeur sont véritablement contagieux. Mes directeurs ont tous deux su me conseiller, m'encourager et m'inspirer dans les moments les plus critiques de mon cheminement. Au-delà des méandres de la recherche, ils démontrèrent des qualités humaines remarquables et je les remercie pour tous les moments de qualité que nous avons partagés.
Je me dois de m'incliner bien humblement devant mes collègues et amis des laboratoires Bernatchez, Couture, Derome et du LARSA pour leur aide, soutien, amitié et sens de l'humour. Pour leur aide académique et soutien, je recermie spécifiquement Éric N., Julie G., Bérénice B., Vincent B., Jean-Christophe T., Guillaume C , Lucie P., Pierre-Alexandre G., Christopher S., Fabien-Claude L., Fabien P., Éliane V., Sébastien B., Nathalie B., Caroline C, François-Olivier G, Geneviève O., Charles P., Scott P., Magnus J., Jérôme S.,
Gaétan L., Larose S., Laroche J., Johan P., Mélanie B., Olivier P., Charlotte T., Ferguson D., Serge H., Gaétan L., Mike D. et Brian B.
Je termine en remerciant ma famille qui m'a toujours encouragé à persévérer pour atteindre le sommet. Leur soutien et leur amour inconditionnel constituent ma plus grande richesse.
Merci!
Au moment de faire mon dépôt initial, le chapitre 1 n'est pas encore soumis à un périodique scientifique. Les coauteurs du chapitre 1 ont contribué à ma supervision; le travail de laboratoire, les analyses et la rédaction furent réalisés par moi-même sous la supervision de mon directeur (L.B.), co-directeur (P.C.) et collègues énumérés dans les remerciements.
Au moment de faire mon dépôt final, le chapitre 1 est soumis au périodique Ecotoxicology dans le but d'être publié en tant qu'article original dans un numéro spécial portant sur les effets à long termes des contaminants. Les coauteurs sont Patrice Couture, Peter G.C. Campbell et Louis Bernatchez.
Ce travail n'aurait pu être possible sans le support financier du Conseil de recherches en sciences naturelles et en génie du Canada (CRSNG), du groupe interinstitutionnel de recherches océanographiques du Québec (Québec-Océan) et de la compagnie minière Vale.
Table des matières
RESUME iii
ABSTRACT iv AVANT-PROPOS v
TABLE DES MATIÈRES viii
LISTE DES TABLEAUX x LISTE DES FIGURES xi
INTRODUCTION 1 ÉMERGENCE DE L'ÉCOTOXICOLOGIE ÉVOLUTIVE 1
APPROCHE CONCEPTUELLE 3
La mutagénèse 4 La sélection naturelle 5
La dérive génétique et le goulet d'étranglement 1
Le flux génique 8 ÉTUDE DE L'EVOLUTION ADAPTATIVE FACE À LA POLLUTION 10
Origine et fondements théoriques 10
Approche moderne 12 Applications et potentiel futur 14
BIOLOGIE DE LA PERCHAUDE 16
PROBLÉMATIQUE 17
OBJECTIFS 18 CHAPITRE 1 EVOLUTIONARY CHANGE DRIVEN BY METAL EXPOSURE
AS REVEALED BY CODING SNP GENOME SCAN PN WILD YELLOW PERCH
(Perca flavescens) 20 RÉSUMÉ FRANÇAIS 21
INTRODUCTION 22 MATERIAL AND METHODS 24
Sampling 24 Metal quantification 24
Coding gene SNP development and genotyping 24 Candidate gene library preparation for large amplicon 454 sequencing 25
Descriptive statistics 27 Temporal stability 27 Outlier detection of selection 28
Genetic association with metal contamination 38 Sequence characterization of candidate genes 29
RESULTS 32 Metal quantification 32
Marker development 33 Temporal stability 33 Descriptive statistics 34
Outlier detection and association tests 34 Mapping of candidate genes 3 5
Sequence analysis of cyclin Gl 35 Sequence analysis of the mitogenome 36
DISCUSSION 38 Adaptive importance of cyclin GI for metal tolerance 39
Adaptive importance of NADH dehydrogenase subunit 2 for metal tolerance 42
Implications for conservation biology and impact assessment 44
CONCLUSION AND PERSPECTIVES 46
CONCLUSION 59 BIBLIOGRAPHIE 64 ANNEXE 1 73 ANNEXE 2 75 ANNEXE 3 77 ANNEXE 4 78 ANNEXE 5 79 ANNEXE 6 81 ANNEXE 7 83 ANNEXE 8 85 ANNEXE 9 86
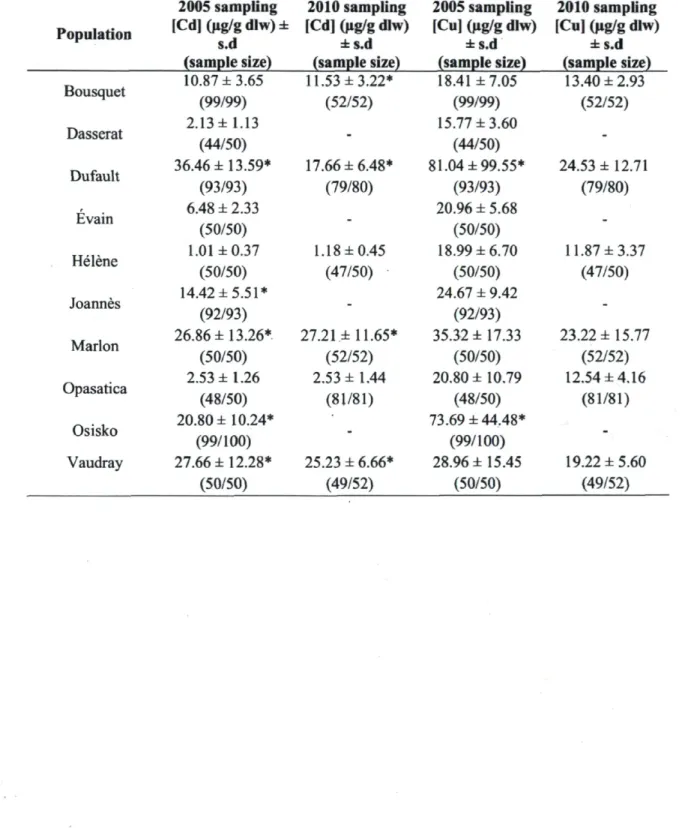
Table 1. Population means of hepatic Cd and Cu concentrations. All bioaccumulation values are population means calculated from individual values measured as an average of three ICP-MS measurements per sample, s.d. = standard deviation, dlw = dry liver weight. Sample sizes show the number of values used for average calculation (taking into account missing data) versus the number of individuals sampled and used for this study. * Indicates average values above significant contamination thresholds (11 ug/g dlw for Cd and 38.8 ug/g dlw for Cu) set by
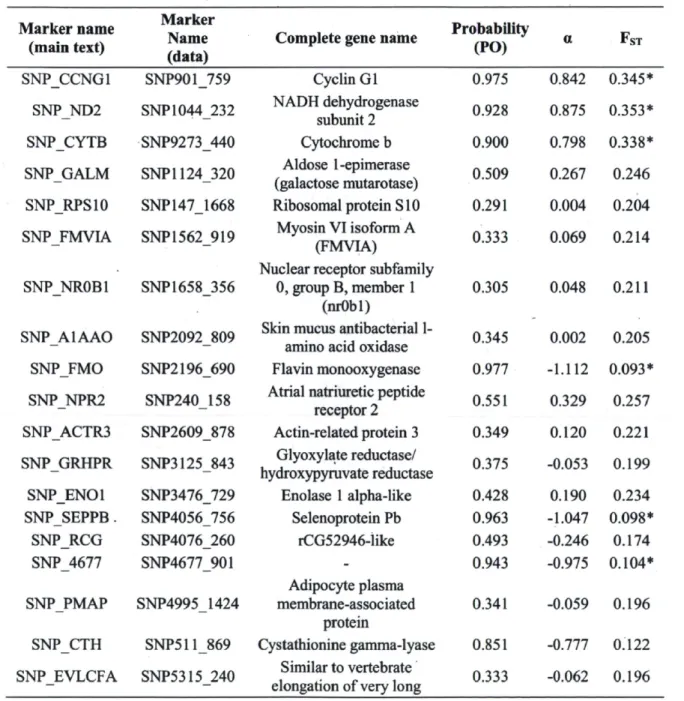
Couture and Pyle (2008) 47 Table 2. Outlier detection and gene annotation. Output of outlier test performed with
BayeScan, PO = probability of being under selection compared to neutrality, a = strength and directionality of selection, FST = bayesian F-statistic. Marker names in the main text are abbreviations of the genes on which the markers are located. They represent usual abbreviations for each gene when possible. Marker names in the data have numbers referring to the contig number and position on which they are located in Pierron et al. (2009) assemblies. Complete gene name was found by blasting (Blastn and Blastx; NCBI) a large sequence surrounding the SNP and always correspond to results with e-values > 1E-30. * Indicates outlier FST values; positive a values indicate directional selection while negative values indicate
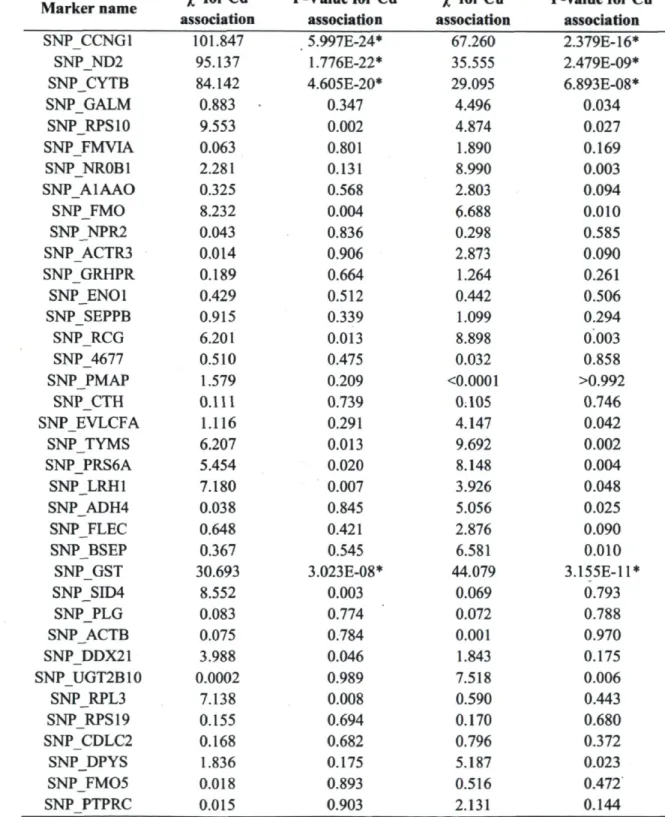
balancing selection 48 Table 3. Stratification-corrected x2 associations with population mean hepatic metal
contamination. ""Indicates significant values after Bonferroni correction (a = 0.05,
critical value = 0.001) 51 Table 4. Characteristics of non-synonymous substitutions on candidate genes. All
references are made in relation to a consensus contig (see Annex 7 for haplotype details and accession numbers). Reference SNPs are underlined (position 4876 =
Liste des figures
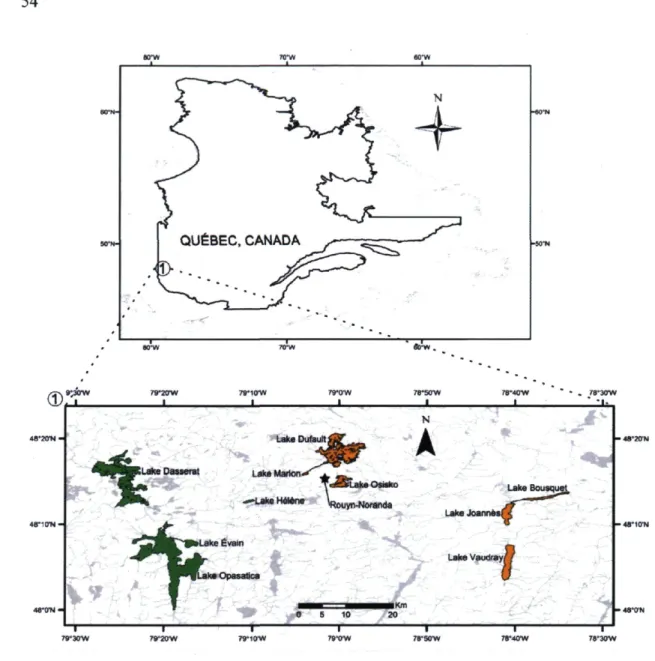
Figure 1. Map of study area and sampled lakes. In green, control lakes; in orange,
contaminated lakes. The star indicates Rouyn-Noranda from where metal emissions
take place 54 Figure 2. Scaled representation of annotated exons for each gene (partial or complete)
located in cyclin Gl genomic region. Numbers under grey boxes represent annotated exons under the figure. Exons number 1 are initiation exons with start codons (except for septin 8, which is partial). Polyadenylation signals (A) of cyclin Gl and NudC2 are side by side; none was found for septin 8 and GABRA6's signal is outside of the contig. Amplicon names/lengths can be used to retrieve
corresponding primer sequences (Annex 1). Annotation : Gamma-aminocutyric acid A receptor, alpha 6-like (GABRA 6; - strand; exon 1: 772-818; exon 2:
589-695), NudC domain containing protein 2-like (NUDC2; + strand; exon 1 (init): 1461-1649; exon 2:2861-2909; exon 3: 3005-3156; exon 4 (term): 3968-4051; polyA signal: 4373-4378), Cyclin Gl (CCNG1; strand -; exon 1 (init): 7508-7771; exon 2: 7155-7408; exon 3: 6686-6764; exon 4: 5457-5864; exon 5 (term): 5353-5544; PolyA signal: 4381-4386), Septin 8 (SEPT8; - strand; exon 1: 16627-16823; exon 2: 15908-16094; exon 3: 15653-15814; exon 4: 15121-15217; exon 5: 14816-14984; exon 6: 13489-13621; exon 7: 13171-13361; exon 8: 11466-11543). Init = Contains initiation (START) codon. Term = Contains terminal (STOP) codon. PolyA signal = Polyadenylation signal. GABRA6 and septin 8 do not have initiation or terminal codons because they are partial on this contig or because they could not
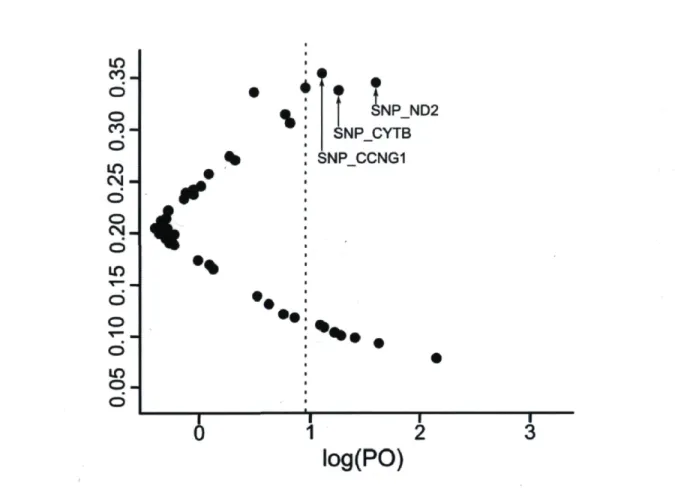
be inferred 55 Figure 3. Results of the BayeScan 2.0 outlier test. Posterior probability significance
threshold (vertical bar) of 0.90 after Bonferroni correction (a = 0.05). Outliers most likely under directional selection are labeled (see Table 4 for complete gene
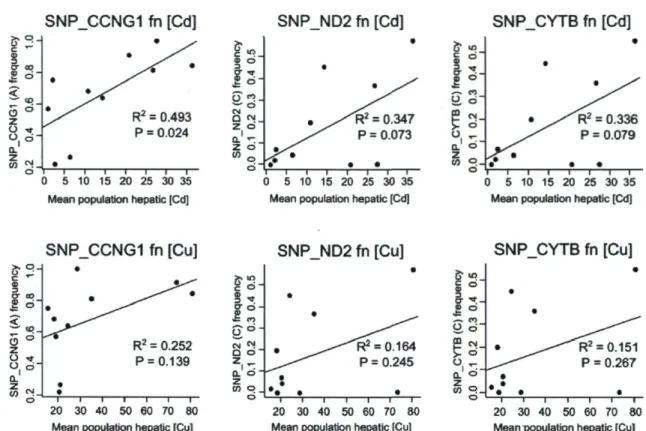
names) 56 Figure 4. Linear regressions between candidate SNP allelic frequencies in each population
and mean population hepatic contamination. P-value significance threshold of 0.001 after Bonferroni correction (a ■ 0.05). Computed in the R 2.13.0 environment (R
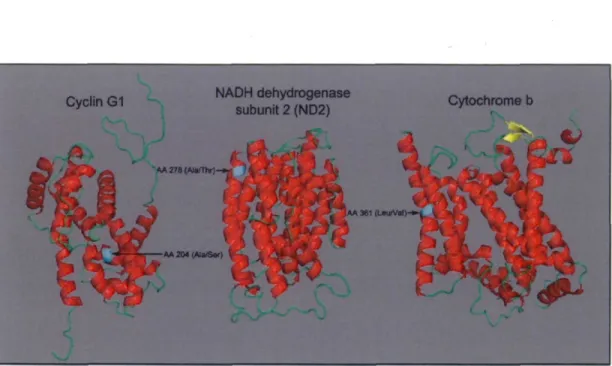
core development team) and edited with InkScape 0.48 57 Figure 5. Candidate proteins 3D models. Red, alpha helixes; yellow, beta sheets; green,
loops; cyan, candidate SNP for each characterized protein. Models were created in conjunction between GeneSilico prediction metaserver (Kurowski and Bujnicki 2003) and FRankenstein3D (Kosinski et al. 2003), visual output produced with
L'être humain exerce désormais une domination absolue sur les écosystèmes de notre planète tant par leur exploitation directe que par leur dégradation indirecte (Vitousek et al,
1997; Akimoto, 2003). Par surcroît, il apparaît évident que la pérennité de nos civilisations dépend directement de l'intégrité de l'écosystème global. Parmi les plus grands destructeurs à l'échelle planétaire, la pollution atmosphérique pose un problème particulier puisqu'elle ne connaît pas de frontières. Ainsi, la coopération tant interdisciplinaire qu'internationale est toute indiquée pour s'attaquer à ce problème qui ravage nos étendues naturelles et nourricières. Or, la communauté scientifique s'accorde pour dire que les connaissances véritables sur les ramifications des impacts environnementaux des polluants (xénobiotiques) demeurent parcellaires (Hooper et al., 2005). Il s'avère aussi flagrant que tout geste de protection mal guidé n'est généralement qu'un coup d'épée dans l'eau doublé d'un gaspillage de ressources intellectuelles et financières (Ludwig et al, 1993). Un survol historique de l'émergence de l'écotoxicologie évolutive, une discipline scientifique visant à combler un pan de cette ignorance, est essentiel pour introduire les fondements et motifs de ce mémoire. Dans le même but, nous ferons ensuite un survol des concepts importants du domaine en question et détaillerons particulièrement l'étude de l'évolution adaptative dirigée par la pollution anthropique.
Émergence de l'écotoxicologie évolutive
Aux racines de la science moderne, la toxicologie s'est initialement développée dans le domaine médico-légal et visait surtout la mise au point de méthodes permettant la détection des poisons utilisés pour camoufler des meurtres en décès naturels. Plus tard, il fut découvert qu'un agent chimique donné peut à la fois présenter des propriétés toxiques et médicinales, dépendamment de la dose administrée. Ce constat important fit prendre un virage pharmaceutique à la toxicologie. Il faut donc attendre l'ère industrielle du 19e siècle,
aussi appelée ère chimique, pour que la toxicologie s'intéresse véritablement aux effets néfastes qu'ont les polluants sur les êtres vivants. On s'intéresse alors particulièrement aux effets sur l'être humain que peuvent avoir les agents chimiques composant les effluents
L'approche toxicologique dite «classique» consiste alors à exposer des animaux à diverses doses de contaminants en laboratoire afin d'en documenter les effets ainsi que les doses délétères ou mortelles (Barnes & Stoner, 1959). Éventuellement, cette approche qui est toujours d'usage pour certaines applications, commença à être critiquée pour son manque de pertinence par rapport à la complexité des environnements chimiques des contextes naturels (Mendrick, 2008; Hartung, 2009). Plusieurs travaux vinrent démontrer l'existence d'interactions complexes entre certains polluants (Unger et al, 2006), leur bioaccumulation dans les tissus (Hare, 1992; Morel et al, 1998), leur biomagnification à travers la chaîne alimentaire (Houde et al, 2006) ainsi que les perturbations qu'ils causent à l'échelle de l'écosystème (Sherwood et al, 2002). Il apparut alors clair que la toxicologie se devait de migrer vers une approche écosystémique intégrée permettant d'obtenir une véritable compréhension des ramifications des impacts des polluants (Truhaut, 1977; Chapman, 2002).
René Truhaut proposa en 1969 le terme écotoxicologie pour désigner cette nouvelle science qui porte sur l'étude des effets néfastes de la pollution chimique sur les constituants des écosystèmes naturels (Truhaut, 1977). Les bases de l'écotoxicologie moderne furent alors posées, pavant la voie à l'étude des effets des contaminants sur un vaste éventail de niveaux d'organisation allant du métabolisme à l'écosystème, en passant par la cellule, l'organe, l'individu et la communauté (Chapman, 2002). En 1992, le Sommet de la Terre de Rio de Janeiro attira enfin l'attention internationale sur l'importance du concept de diversité génétique, une composante de la biodiversité, en l'identifiant comme un élément crucial à toute stratégie de conservation (Panjabi, 1997). Partant de cet impératif et sachant que plusieurs polluants peuvent altérer la dynamique génétique (i.e. taux mutationnel, dérive génétique, sélection directionnelle et migration) des populations sauvages (Belfïore & Anderson, 2001), l'écotoxicologie se devait naturellement d'évoluer vers l'inclusion des considérations génétiques. Il s'avère également que lorsqu'un processus adaptatif survient dans une population et qu'il n'est pas pris en compte, l'évaluation de l'impact de la
Ainsi, une population peut paraître saine tout en subissant d'importants dommages écologiques relatifs à la perte de diversité génétique ainsi qu'aux effets de la pléiotropie négative (i.e. effets néfastes découlant de mutations sur un gène qui détermine plusieurs phénotypes) sur le potentiel adaptif (Medina et al, 2007). Certains chercheurs soutiennent même que l'identification d'adaptations à la pollution pourrait servir à modifier génétiquement des animaux menacés afin de les réimplanter au sein d'environnements perturbés par l'homme (Rice & Emery, 2003).
Vers 1993, suite à la prise de conscience internationale face à l'importance de la diversité génétique, une nouvelle discipline née de la combinaison de l'écotoxicologie et de la génétique des populations (Bickham & Smolen, 1994) émerge : l'écotoxicologie évolutive, aussi appelée génotoxicologie ou toxicologie génétique. Cette science pluridisciplinaire se concentre alors sur la compréhension des effets des polluants sur la dynamique génétique des populations animales et végétales (Matson et al, 2006; DeWoody, 2010) dans le but ultime de guider les stratégies de conservation de la biodiversité (Bickham et al, 2000) et de restauration (Rice & Emery, 2003). Bien sûr, les progrès en écotoxicologie évolutive visent également l'étoffement fondamental de la théorie de l'évolution qui revêt elle-même un caractère crucial pour différentes sphères d'activité humaine.
Approche conceptuelle
Les polluants peuvent affecter spécifiquement plusieurs processus qui déterminent eux-mêmes la dynamique génétique des populations sauvages. Les principaux mécanismes en question sont la mutagénèse, la sélection naturelle, la dérive génétique, le goulet d'étranglement et même le flux génique (Belfïore & Anderson, 2001). Ces concepts sont inter-connectés et leur étude conjointe est la seule façon de véritablement comprendre la dynamique génétique d'une population chroniquement (i.e. de façon continue, à long terme) exposée à la pollution. La présente section vise à présenter l'état actuel de la compréhension des effets génétiques qu'ont les polluants et ce, dans l'optique d'une étude d'écotoxicologie évolutive.
mutation génétique, généralement en provoquant des dommages directs à l'ADN ou en inhibant la machinerie cellulaire de réparation de l'ADN, comme le cadmium chez les mammifères (Bertin & Averbeck, 2006). Parmi la liste d'agents génotoxiques, les radiations, les métaux, les HAP (hydrocarbones aromatiques polycycliques), les amines hétérocycliques, les pesticides et les déchets pétrochimiques sont notoires (Bickham & Smolen, 1994; Ohe et al, 2004). Les mutations découlant de la génotoxicité peuvent affecter les gamètes d'un organisme (mutation germinale) ou toute autre cellule (mutation somatique). Les mutations somatiques ne se transmettent pas à la progéniture puisqu'elles n'affectent pas les cellules participant à la reproduction. Cependant, leur accumulation entraîne une diminution de la viabilité (e.g. en déclenchant des cancers) et du potentiel adaptatif des individus ce qui nuit à la dynamique globale des populations touchées (Bickham & Smolen, 1994). D'un autre côté, les mutations germinales se transmettent aux générations suivantes et peuvent même parfois présenter un caractère adaptatif pour les individus contaminés (Bickham & Smolen, 1994). Dans une telle éventualité, le polymorphisme adaptatif est généralement sélectionné, ce qui contribue à la viabilité des individus dans l'environnement pollué.
De façon plus générale, lorsque le taux mutationnel est élevé, l'accumulation de mutations délétères (i.e. compromettant la fonction d'un gène) au fil des générations peut entraîner une «fonte mutationnelle» (mutational meltdown) (Lynch et al, 1995). Les individus accumulent trop de mutations réduisant leur capacité de survivre et de se reproduire ce qui provoque la disparition de la population en question. Le phénomène est particulièrement sévère chez les populations de petite taille qui ont une capacité réduite à purger (i.e. éliminer par contre-sélection) les alleles délétères (Lynch et al, 1995). En définitive, les mutations découlant de la génotoxicité peuvent soit fournir une nouvelle variabilité génétique sélectionnable permettant la microévolution de la tolérance à la pollution ou encore entraîner des effets délétères qui altèrent la dynamique génétique d'une population, menant parfois à son effondrement.
La sélection naturelle est un mécanisme par lequel les traits permettant aux individus de survivre et de se reproduire avec succès dans un certain contexte environnemental sont préférentiellement transmis au fil des générations (Darwin, 1859). Les polluants environnementaux, dans certaines conditions, peuvent créer des pressions sélectives. Celles-ci surviennent généralement lorsqu'un contaminant a un impact néfaste sur le cycle vital, la survie ou le métabolisme, donc sur le potentiel adaptatif (fitness) des individus d'une population (Reznick & Ghalambor, 2001; Mineau, 2005; Meyer et al, 2010). Dans de tels cas, la sélection favorisera des phénotypes visant à compenser pour les impacts néfastes, mais seulement si ces traits ont des bases génétiques héritables (Belfïore & Anderson, 2001). Ce succès accru fait augmenter la fréquence du phénotype avantageux dans les générations suivantes et la population entière finit généralement par acquérir le trait en question (issue dépendante de l'environnement évolutif, du gradient de sélection et l'héritabilité du trait (Posthuma & Vanstraalen, 1993)). Selon une approche mathématique, la réponse à la sélection (R) peut être décrite quantitativement grâce à l'équation R= h2*S
où h représente le degré d'héritabilité du trait et S le différentiel de sélection (Falconer, 1981). Cette équation illustre comment la réponse évolutive est le produit de l'héritabilité d'un trait et de la force de la sélection. Dans le contexte de la contamination environnementale, les pressions sélectives sont généralement fortes ce qui peut rapidement produire de fortes réponses évolutives (i.e. la modification rapide d'un trait phénotypique) lorsque les phénotypes sont héritables (van Straalen & Timmermans, 2002).
On considère habituellement la diversité génétique de départ (standing genetic variation) comme un élément déterminant pour la survie d'une espèce lorsqu'une pression sélective intense survient brusquement (Barrett & Schluter, 2008). En effet, il n'est pas rare d'observer des taux de mortalités importants lorsque la contamination est élevée ce qui peut résulter en de fortes pressions sélectives (Posthuma & Van straalen, 1993). Cette réalité peut mener à l'extinction de la population contaminée si elle ne dispose pas déjà de polymorphismes génétiques permettant une tolérance accrue au dit polluant. À plus long terme, si la population parvient à survivre, les mutations aléatoires du génome produiront
vue comme un processus opportuniste agissant sur la variabilité existante ou sur celle qui survient au hasard. Ainsi, l'évolution tend à optimiser des mécanismes existants pour leur donner des fonctions nouvelles plutôt que de produire des solutions radicalement nouvelles.
Il ne faut toutefois pas voir la sélection naturelle comme un processus omnipotent; elle est soumise à certaines contraintes qui concourent à déterminer sa trajectoire. La contrainte énergétique en est un exemple particulièrement pertinent dans le contexte d'un écosystème chamboulé par une substance toxique. Chaque animal dispose d'une quantité finie d'énergie et de ressources (e.g. acides aminés, lipides, glucides et autres nutriments) que son organisme investit dans chacune des fonctions vitales: survie, croissance et reproduction (Sibly & Calow, 1989). L'énergie et les ressources sont généralement limitantes, surtout lorsqu'un polluant vient anéantir une partie de l'écosystème d'où les individus puisent leur ressources (Sibly & Calow, 1989). Cette situation entraîne obligatoirement un compromis entre survie, croissance et reproduction ce qui va contraindre la nature des solutions évolutives viables (Sibly & Calow, 1989; Posthuma & Van straalen, 1993). La théorie de réponse du cycle vital au stress de Sibly et Calow (1989) stipule qu'un stress réduisant la qualité d'un environnement poussera l'évolution à favoriser des adaptations à faible coût énergétique qui permettront la completion rapide du cycle vital. Cela est dû au fait que les adaptations pour la réparation des dommages et la défense contre les polluants (e.g. détoxication) sont trop coûteux énergétiquement pour représenter un avantage évolutif dans une telle situation. Un animal énergétiquement limité qui investirait presque toute son énergie pour détoxiquer son organisme et réparer les dommages subis pourrait difficilement croître et ne parviendrait pas à compléter son cycle vital. Au contraire, un animal qui investit toute son énergie pour croître et se reproduire le plus rapidement possible, aura plus de chances de compléter son cycle vital puisqu'il minimise la durée de l'intervalle naissance-reproduction au cours duquel il risque de mourir des suites de la contamination (Sibly & Calow, 1989). Cet exemple illustre bien comment certaines contraintes produisent différentes solutions évolutives, selon le contexte.
organismiques qu'écosystémiques. La compréhension de la sélection naturelle ainsi que de la trajectoire évolutive provoquée par diverses circonstances sont d'ailleurs des sujets centraux à l'écotoxicologie évolutive. Le savoir relatif à ces questions est toujours incomplet et la variabilité des contextes évolutifs fait que l'issue de la sélection est souvent imprévisible.
La dérive et le goulet d'étranglement
Le terme goulet d'étranglement (bottleneck) désigne un événement prononcé de dérive génétique produisant à terme une population à la diversité génétique extrêmement réduite (Nei et al, 1975). Il faut savoir que la dérive génétique est un processus naturel qui survient dans toute population animale dont le nombre effectif (nombre de reproducteurs génétiquement distincts) n'est pas infini (Lande, 1976). Par exemple, les lacs d'eau douce isolent les poissons en populations distinctes de nombre fini dont la diversité génétique sera affectée par la dérive génétique. La dérive est un processus neutre (i.e. non-directionnel, contrairement à la sélection) dirigé par le hasard et régi par la loi des grands nombres. Plus spécifiquement, la loi des grands nombres exprime la réalité selon laquelle plus un échantillon est grand, plus ses caractéristiques statistiques se rapprocheront de la population dans laquelle il a été prélevé (Lande, 1976). À l'inverse, un petit échantillon a plus de probabilités d'être biaisé par rapport à la population d'où il est originaire. Les générations se succédant au sein des populations animales, il se produit un tirage matérialisé par la reproduction de certains individus et la transmission de leurs gènes à la génération suivante (i.e. l'échantillon). Or, la taille de ce tirage n'équivaut jamais à la taille totale de la population puisque certains individus ne se reproduisent pas et survient ainsi un biais aléatoire à chaque génération qui fera dériver la fréquence de polymorphismes génétiques soit vers la fixation (fréquence populationnelle de 1) ou vers la disparition (fréquence populationnelle de 0) (Lande, 1976). À titre d'exemple, on peut s'imaginer un polymorphisme génétique bi-allélique de fréquence 0,5 dans une population parentale. L'un des deux alleles pourrait voir sa fréquence passer à 0,4 dans la génération suivante par le seul effet de la loi des grands nombres. Sa fréquence variera ainsi stochastiquement lors des
générations suivantes jusqu'au point où sa fréquence sera tellement faible que si aucune force évolutive ne le maintient, le polymorphisme disparaîtra.
À cet effet, il est bien connu que plusieurs polluants peuvent entraîner des taux de mortalité bien au delà des valeurs naturelles chez plusieurs populations animales. Cette mortalité élimine des variants génétiques rares et diminue la taille de l'échantillon sélectionné à chaque génération (Posthuma & Van straalen, 1993). Cela peut provoquer une diminution dangereuse de la diversité génétique populationnelle, ce qui entraine généralement un goulet d'étranglement (Nei et al, 1975). À moins que la migration ou les mutations régénèrent la diversité perdue, la population victime de ce goulet d'étranglement risque de dériver tellement qu'elle en devient consanguine (Posthuma & Van straalen, 1993). On assiste alors à la reproduction entre individus apparentés ce qui diminue ultimement le potentiel adaptatif des rejetons. Ce phénomène est notamment causé par l'augmentation de l'homozygotie (i.e. la détention du même allele sur les deux copies d'un chromosome) d'allèles délétères récessifs qui rendent les individus mal adaptés. À cela s'ajoute la réduction de l'hétérozygotie (i.e. la détention de deux alleles différents sur chaque copie d'un chromosome) de certains gènes pour lesquels la diversité est essentielle à la fonction. Par exemple, l'hétérozygotie des gènes du complexe majeur d'histocompatibilité détermine en partie l'efficacité du système immunitaire (Allendorf, 1986).
Globalement, la dérive et son cas extrême, le goulet d'étranglement, sont des phénomènes particulièrement importants à documenter lorsque l'on s'intéresse aux effets génétiques qu'ont les polluants sur une population. L'étude de la dérive permet, entre autres, de déterminer si une population affectée par la pollution dispose de suffisamment de diversité génétique pour continuer à prospérer (Medina et al, 2007).
Le flux génique
Les effets potentiels de la contamination sur le flux génique (i.e. le transfert de variation génétique découlant de la migration d'individus d'une population à l'autre) demeurent peu étudiés. Certains auteurs reconnaissent toutefois l'importance potentielle de ce phénomène en nature (Bickham & Smolen, 1994). À cet effet, le mécanisme potentiel le plus évoqué
qui dispose d'un certain degré d'adaptation génétique pour tolérer le contaminant local (Beaty et al, 1998). Cette mortalité spécifique aux immigrants non-adaptés à la contamination réduit les possibilités d'accroissement de la diversité génétique des populations contaminées. Le résultat peut être un degré d'isolement accru favorisant les premières étapes de la spéciation ou bien une accélération de la dérive génétique entraînant un degré de consanguinité handicapant et fatal pour la population (Bickham & Smolen, 1994; Beaty et al, 1998). De futurs travaux viendront sans doute préciser cette hypothèse car il pourrait s'agir là d'un élément crucial contribuant établir des stratégies de conservation efficaces chez les populations adaptées à la contamination.
En résumé, les précédentes sections démontrent comment la dynamique génétique d'une population affectée par la contamination peut être chamboulée par divers procédés reliés entre eux. D'abord, la génotoxicité de certains polluants provoque des mutations pouvant produire de la diversité alimentant le moteur évolutif ou bien, au contraire, entraîner une fonte mutationnelle fatale. Quant à elle, la sélection naturelle dirigée par la contamination répare et ajuste des mécanismes préexistants grâce à la diversité disponible et aux mutations tout en étant restreinte par des facteurs externes, comme la disponibilité en énergie. Ensuite, la dérive génétique peut être augmentée par la mortalité due à la toxicité, résultant parfois en un goulet d'étranglement et en une hausse handicapante de la consanguinité. Finalement, le flux génique introduit peut se voir interrompu chez les populations contaminées, contribuant à la spéciation ou à une dérive génétique accélérée. Chacun de ces aspects est important à prendre en compte dans une étude d'écotoxicologie évolutive. Un portrait complet des effets de la contamination sur une population animale offre la possibilité de bien cerner la nature et la directionalité des phénomènes génétiques à l'œuvre. Cela permet ultimement d'élaborer une stratégie de conservation ou de mitigation efficace, de déterminer un seuil néfaste de contamination ou simplement de contribuer à étoffer le savoir fondamental en écotoxicologie évolutive.
Étude de l'évolution adaptative face à la pollution
L'étude des processus génétiques affectés par les polluants (i.e. mutation, dérive et flux génique) est indispensable pour guider les programmes de conservation de la biodiversité et pour comprendre la dynamique génétique des populations contaminées. Toutefois, la mise en évidence de l'évolution adaptative est le thème principal de l'écotoxicologie évolutive et de ce mémoire; il convient ainsi d'en dresser un portrait plus détaillé. Dans ce but, les fondements théoriques et méthodes de mise en évidence de l'évolution adaptative doivent être présentés. La première partie portant sur l'origine des fondements théoriques de l'étude de l'évolution explique l'émergence des méthodes et concepts devenus centraux à l'approche moderne, faisant elle-même l'objet de la seconde partie. La troisième partie conclut en démontrant la puissance des applications actuelles de l'écotoxicologie évolutive et propose une ouverture quant au potentiel futur de la discipline en regard des progrès prévus au niveau des technologies de séquençage.
Origine et fondements théoriques
A ses origines, la perception scientifique dominante de la théorie de l'évolution (Darwin, 1859) en fait un processus extrêmement lent que l'on croit alors difficile, voire impossible, à observer à l'échelle de la vie humaine (Gingerich, 2009). De nos jours, il n'existe toujours pas de consensus et beaucoup de chercheurs croient encore que l'évolution est obligatoirement lente. Toutefois un groupe grandissant de scientifiques, dont John N. Thompson (1998), David Reznick (2001) et Sephen Palumbi (2001), défendent l'idée selon laquelle l'évolution peut être extrêmement rapide (Gingerich, 2009). Bien que le débat sur le rythme de l'évolution entoure surtout la question du rythme de spéciation (i.e. l'apparition d'une nouvelle espèce), l'étude de la microévolution s'avère cruciale pour décrire les processus qui sont justement sous-jacents à la spéciation.
Il s'avère que seulement quelques années peuvent être suffisantes pour qu'un processus évolutif adaptatif quantifiable survienne (Gingerich, 2009). À ce sujet, Nacci et al. (1999) ont d'ailleurs démontré que 15 générations avaient été suffisantes pour que des populations de Fundulus heteroclitus évoluent vers une tolérance aux contaminants de la famille des
dioxines. En fait, la documentation de processus adaptatifs rapides face à la pollution n'est pas une nouveauté. L'un des premiers cas bien documentés concerne un lépidoptère, la phalène du bouleau. Kettlewell (1961) décrivit comment l'abondance d'un morphe foncé de ce papillon augmenta drastiquement suite à la révolution industrielle de Grande-Bretagne. En effet, le charbon, alors abondamment utilisé comme source d'énergie, produisait de grandes quantités de suies noires qui se déposaient sur toutes les surfaces, dont les arbres. Le camouflage clair des papillons devint alors flagrant aux yeux perçants des prédateurs, ce qui contre-sélectionna le morphe clair et favorisa le morphe foncé, jusqu'alors excessivement rare. Les populations de papillons exposées à la suie passèrent alors rapidement d'un phénotype clair à un phénotype beaucoup plus foncé. Plus tard, l'utilisation du charbon décrut, certaines lois interdirent l'émission excessive de suies et ainsi, les surfaces redevinrent propres. On nota alors la quasi-disparition du morphe foncé et le retour du morphe clair. Ce cas d'école attira l'attention sur la rapidité avec laquelle l'évolution adaptative peut survenir en nature et inspira plusieurs travaux qui jetèrent ensuite les bases de l'écotoxicologie évolutive. (Kettlewell, 1961)
Toutefois, il faut dire que le cas des papillons porte sur une perturbation environnementale évidente provoquant l'évolution d'un phénotype facilement observable. Le caractère flagrant de cet exemple en fait une situation d'exception. Effectivement, les principaux effets néfastes de la pollution chez les organismes sont habituellement internes et physiologiques (Posthuma & Van straalen, 1993). Conséquemment, la réponse évolutive associée concerne généralement des mécanismes intrinsèques qui sont parfois uniquement observables à l'échelle moléculaire. L'augmentation récente du pouvoir analytique des outils de biologie moléculaire a véritablement permis à l'écotoxicologie évolutive de s'attaquer aux cas plus subtils et fréquents d'évolution dirigée par la pollution.
Posthuma et van Straalen (1993) furent parmi les premiers à publier une revue de littérature complète portant sur des travaux faisant fortement appel à la biologie moléculaire pour mettre en évidence l'évolution adaptative rapide en milieu contaminé. Dans ce qui peut être considéré comme l'un des articles fondateurs de l'écotoxicologie évolutive, les auteurs discutent de questions devenues fondamentales pour la discipline (Posthuma & Van
straalen, 1993). Plusieurs des considérations qu'ils soulèvent sont d'ailleurs toujours importantes près de 20 ans plus tard, notamment concernant la nature des facteurs poussant une population animale à s'engager dans une stratégie évolutive coûteuse (e.g. détoxication et réparation) versus une stratégie peu coûteuse (e.g. tolérance et cycle vital accéléré). En plus de discuter des questions centrales à l'écotoxicologie évolutive, Posthuma et van Straalen regroupent certains des meilleurs exemples de méthodes ayant servi à démontrer l'évolution face à la pollution. À l'époque de la publication, les marqueurs moléculaires servent déjà à révéler plusieurs gènes qui sont sélectionnés par les contaminants, dont la métallothionéine, une enzyme de détoxication (Posthuma & Van straalen, 1993). En plus de détecter statistiquement le signal de la sélection, on se sert alors des caractéristiques moléculaires des gènes associés à des marqueurs candidats pour inférer des hypothèses adaptatives. C'est donc par la transposition des méthodes servant à l'étude de l'évolution adaptative dans son sens le plus large qu'apparaît la méthodologie de caractérisation de l'adaptation à la pollution.
Approche moderne
En 2012, l'approche de mise en évidence de l'évolution adaptative face à la pollution est toujours fidèle aux origines de l'écotoxicologie évolutive. Il s'agit de mettre en évidence les patrons moléculaires résultant de la sélection naturelle et d'établir des corrélations (relations linéaires ou logistiques selon le modèle préconisé combiné à différents scénarios exprimant la base génétique d'un trait: additivité, dominance, codominance, récessivité, épistasie (Balding, 2006)) entre des polymorphismes candidats et des traits potentiellement adaptatifs, souvent grâce à la comparaison de populations saines et contaminées. Cependant, la puissance grandissante des technologies moléculaires permet désormais de ratisser le génome plus largement afin de découvrir un vaste éventail de nouveaux polymorphismes adaptatifs. Le séquençage à haut débit de génomes et de transcriptomes entiers fournit les données de base qui permettent de découvrir un nombre croissant d'adaptations génétiques potentielles (Gilad et al, 2009). Ces approches que l'on surnomme généralement genome-wide ou transcriptome-wide portent souvent sur le séquençage du patrimoine génétique de multiples individus provenant de différentes populations. Les séquences ainsi produites peuvent être comparées pour découvrir des marqueurs génétiques
dont la caractérisation permet de chercher le signal de la sélection naturelle (Vignal et al, 2002). Une fois identifiés, ces marqueurs peuvent être génotypes à un prix relativement bas sur des centaines ou des milliers d'individus ce qui permet ultimement d'obtenir suffisamment de données pour étudier l'évolution adaptative dans un contexte particulier.
A ce sujet, la mise en évidence de la sélection naturelle repose beaucoup sur les F-statistiques (FST) développées, entre autres, par Lewontin et Krakauer (1973) en génétique des populations. De façon sommaire, le calcul d'une valeur FST (FST = (n^er - n^tm V n^ter où n est le nombre moyen de différences génétiques entre deux individus d'une même sous-population (n^tra) ou de deux populations différentes (n^ter)) exprime le contraste entre les fréquences génotypiques de deux sous-populations (Lewontin & Krakauer, 1973). Un contraste suffisamment grand entre deux sous-populations produit une valeur de FST qui se situe à l'extérieur d'une distribution de valeurs à laquelle on s'attend lorsqu'un polymorphisme se comporte de façon neutre (i.e. n'est pas soumis à la sélection). C'est alors qu'il est possible de déterminer si la valeur FST indique un patron qui est spécifique à la sélection directionnelle (i.e. diversifiante) ou balancée (i.e. stabilisante).
En complément aux F-statistiques, les statistiques associatives servent à vérifier si la fréquence d'un polymorphisme est corrélée au degré d'un facteur tel la contamination ou encore la capacité à détoxiquer. On s'attend généralement à ce que la fréquence d'un polymorphisme adaptatif augmente (atteignant parfois la fixation) dans les milieux les plus contaminés et diminue dans les moins contaminés (disparaissant des milieux propres) (Balding, 2006). Cette tendance s'explique en partie par le fait que toute adaptation comporte un coût énergétique qui devient superflu et contre-sélectionné lorsque cette dernière est inutile (Sibly & Calow, 1989). Aussi, certaines adaptations contre la pollution deviennent mal adaptées en milieu sain (Sibly & Calow, 1989). Ce phénomène a notamment été observé chez des plantes tolérantes aux métaux dont le potentiel adaptatif se trouve réduit lorsqu'elles poussent dans un sol salubre (Wilson, 1988). Chez les Vertébrés, il fut démontré que des rats adaptés pour tolérer la warfarine, un puissant anti-coagulant et antagoniste de la vitamine K, sont fortement contre-sélectionnés en son absence puisque leurs adaptations deviennent néfastes pour leur survie (Rost et al, 2009). En somme,
l'établissement d'une association entre un polymorphisme et un trait adaptatif ou une pression sélective particulière constitue un outil puissant pour inférer l'implication du dit polymorphisme au sein d'un processus adaptatif. Il existe plusieurs modèles différents pour faire des statistiques associatives (e.g. régression linéaire, régression logistique, test de Chi-Carré) et chaque méthode dispose de ses forces et faiblesses. Une approche répandue, utilisée dans le cadre des travaux présentés, consiste à faire appel à plusieurs modèles afin de croiser leurs résultats pour déterminer quels marqueurs renvoient les signaux les plus cohérents (e.g. Narum et al, 2010). Cela permet d'éliminer les faux-positifs des différentes méthodes statistiques et de se concentrer sur les cibles candidates les plus prometteuses pour expliquer l'évolution face à la contamination.
En somme, l'approche moderne de mise en évidence de l'évolution adaptative en écotoxicologie évolutive repose surtout sur la détection de la sélection et les associations statistiques. Il ne faut toutefois pas perdre de vue l'importance des autres éléments essentiels à la caractérisation d'un processus évolutif qui ont été présentés précédemment : mutation, dérive et flux génique. De plus, d'autres indications, tel l'attendu évolutif (i.e. réponse évolutive prédite par un modèle de réponse à la sélection) dans un environnement aux caractéristiques particulières, sont cruciaux à l'interprétation des cas de microévolution adaptative. En définitive, les statistiques portant sur la dimension génétique des populations ne suffisent pas à conclure, hors de tout doute, qu'un polymorphisme donné est véritablement adaptatif. À cet effet, diverses méthodes de modélisation computationnelle permettent d'obtenir des indications supplémentaires quant au potentiel d'un polymorphisme de provoquer, par exemple, une modification de l'activité ou de la fonction d'une protéine. Par dessus tout, il est indispensable d'utiliser des approches in vivo pour établir clairement les ramifications de l'impact fonctionnel d'un polymorphisme candidat pour en valider le caractère adaptatif.
Applications et potentiel futur
Les chercheurs en écotoxicologie évolutive exploitent encore peu les approches mariant la découverte massive de marqueurs à l'échelle du génome aux statistiques associatives. Rappelons d'abord que le coût de séquençage d'un génome entier est passé de 100 millions
US$ en 2001 à 1 million US$ en 2008 pour enfin atteindre la barre des 10 000 US$ en 2011 (Dewitt et al, 2012). En fait, la période de 2007 à 2012 a vu la diminution du coût de séquençage par génome dépasser largement les prédictions de la loi de Moore, diminuant d'un facteur de 10 à chaque année (Dewitt et al, 2012). Cette loi décrit comment le ratio coût/puissance des technologies de l'information suit habituellement un rythme de diminution logarithmique. Par exemple, pour les puces électroniques à transistor, le ratio coût/puissance est divisé par deux tous les deux ans (Lundstrom, 2003).
Jusqu'en 2009, le coût élevé du séquençage à haut débit a surtout confiné l'approche genome wide aux axes de recherches les plus subventionnés, essentiellement au niveau de
la génétique humaine. Il existe néanmoins déjà quelques exemples de travaux utilisant l'approche transcriptome-wide (i.e. visant seulement l'ensemble des gènes exprimés plutôt que le génome entier) en écotoxicologie évolutive. Larissa Willams et al. (2010) ont notamment publié un article dans lequel ils décrivent leur méthode de détection et de sélection de centaines de SNPs (single nucleotide polymorphisms; polymorphisme génétique impliquant un seul nucleotide) chez des populations de fondules (Fundulus heteroclitus) exposées à plusieurs polluants et ce, grâce au séquençage à haut débit. Plus tard, les chercheurs ont utilisé ces marqueurs pour calculer des F-statistiques ainsi que des tests d'association entre la fréquence de polymorphismes sous sélection et la capacité des poissons à détoxiquer. Ils conclurent que plusieurs loci étaient probablement impliqués dans un processus d'adaptation rapide face aux polluants locaux (Williams & Oleksiak, 2011a). Par la suite, les auteurs démontrèrent qu'un polymorphisme trouvé dans le promoteur du gène codant pour le cytochrome P4501A était responsable de la surexpression de ce gène dans les populations contaminées de fondules (Williams & Oleksiak, 2011b). Cette série de publications est parmi les plus solides de celles qui exploitent le séquençage de dernière génération en écotoxicologie évolutive. Toutefois, les prochains mois promettent d'être riches en publications et découvertes faisant appel aux approches de séquençage à haut débit. Le chapitre constituant le noyau de ce mémoire en fera certainement partie.
Alors que s'amorçait cette maîtrise en 2010, il aurait été inimaginable de baser notre méthodologie sur le séquençage de génomes entiers (un peu moins de 100 000 US$ par génome; Dewitt et al, 2012). Or, Life Sciences annonçait en janvier 2012 la commercialisation imminente de sa technologie Ion Torrent qui aura la capacité de sequencer un génome humain entier en deux heures pour seulement 1 000 US$ (DeFrancesco, 2012). À ce rythme, le séquençage d'un génome humain entier pourrait coûter moins de 100 US$ aussi tôt qu'en 2014. Un budget relativement modeste pourra par conséquent bientôt servir à sequencer des centaines de génomes entiers. Cette réalité décuplera par plusieurs ordres de grandeur la puissance statistique des jeux de données utilisés en écotoxicologie évolutive et rendra les méthodes actuelles de découverte de marqueurs et de génotypage obsolètes. Les multiples génomes séquences serviront à la fois à la découverte des millions de marqueurs génétiques et de matériel de génotypage en raison des couvertures de séquençage immenses qui seront possibles à très bas prix.
Il semble que la puissance grandissante du séquençage à haut débit assure un avenir prometteur à l'écotoxicologie évolutive. Plusieurs concepts centraux de la discipline qui sont toujours mal compris pourront être étudiés plus efficacement au cours des prochaines années. D'ailleurs, les espèces non-modèles, comme la perchaude (Perca flavescens), deviennent des candidates de plus en plus attrayantes pour tenter d'élucider la complexité de la dynamique évolutive découlant de l'exposition à la contamination en milieu naturel.
Biologie de la perchaude
La perchaude (Mitchill, 1874) est parmi les Poissons considérés ubiquitaires en Amérique du Nord et son exploitation génère des revenus importants par le biais des pêches sportives et commerciales (Todd & Hatcher, 1993). Perca flavescens habite généralement les zones littorales et limnétiques des lacs tempérés, on la qualifie ainsi de démersale d'eau douce. Elle se retrouve en plus forte abondance dans les lacs présentant une densité moyennement dense de macrophytes, mais n'est pas obligatoirement associée à un type d'habitat spécifique (Engel & Magnuson, 1976). Au niveau de l'alimentation, ce prédateur opportuniste et généraliste peut considérablement modifier sa diète selon l'habitat, l'année,
la saison et le stade du cycle vital (Sandheinrich & Hubert, 1984). En définitive, la perchaude se caractérise par une grande plasticité qui lui confère l'adaptabilité nécessaire pour prospérer dans un vaste éventail de conditions environnementales (Brown et al, 2009). Elle est d'ailleurs souvent parmi les seules espèces parvenant à survivre dans les milieux lacustres fortement altérés par l'acidité ou la pollution métallique (Tremblay et al, 2008). De plus, la perchaude est sédentaire (Aalto & Newsome, 1990) ce qui signifie que sa charge de bioaccumulation reflète la contamination de son milieu. Ces deux dernières caractéristiques en font un organisme de choix pour une étude d'écotoxicologie évolutive.
Problématique
Il existe une littérature abondante concernant les effets du cuivre et du cadmium sur les perchaudes contaminées des écosystèmes lacustres de Rouyn-Noranda (Québec, Canada). La multitude d'effets physiologiques néfastes que ces métaux ont sur les poissons locaux ne font désormais plus aucun doute (Couture & Pyle, 2008). Puisqu'ils peuvent théoriquement avoir un impact sur le potentiel adaptatif des perchaudes, les métaux sont susceptibles d'engendrer un régime sélectif directionnel entraînant un processus adaptatif (van Straalen & Timmermans, 2002). Justement, les travaux de Bourret et al. (2008) sont venus caractériser la dimension génétique des populations contaminées de Rouyn-Noranda et ont démontré la corrélation négative existant entre la bioaccumulation de cadmium et la diversité génétique populationnelle. Plus récemment, Pierron et al. (2009; 2011) ont illustré comment la contamination pouvait influencer le niveau d'expression de plusieurs gènes chez Perca flavescens. Sachant que l'expression génétique comporte une composante déterministe codée par le génome et qu'une altération de la diversité génétique par la contamination peut, entre autres, être le résultat d'un régime sélectif intense (van Straalen & Timmermans, 2002), la possibilité que la microévolution adaptative survienne dans le système s'impose naturellement.
Objectifs
L'objectif principal de cette maîtrise est de déterminer si la sélection naturelle a pu provoquer l'apparition d'adaptations permettant une meilleure tolérance aux métaux chez les populations de perchaudes contaminées de Rouyn-Noranda. Si tel est le cas, un objectif secondaire est de découvrir et caractériser d'hypothétiques polymorphismes permettant cette adaptation. Pour atteindre ces objectifs, il faudra d'abord identifier des marqueurs de type SNP particulièrement différenciés entre les populations de perchaudes saines et contaminées. Dans un deuxième temps, la détection de la sélection et les statistiques associatives devraient permettre de cerner des marqueurs génétiques se situant dans des régions génomiques sous sélection et potentiellement porteuses de polymorphismes adaptatifs. Troisièmement, le séquençage des ces régions candidates devrait rendre possible la caractérisation in silico de polymorphismes potentiellement adaptatifs et ce, au sein des séquences codantes de multiples gènes.
Plus précisément, au niveau expérimental, les séquences transcriptomiques provenant de 32 individus (16 sains et 16 contaminés) seront utilisées pour découvrir des SNPs dont la différenciation entre les milieux sains et contaminés présente les caractéristiques de la sélection naturelle. Ensuite, une sélection de marqueurs sera génotypée chez 1052 individus provenant de 10 populations différentes (six contaminées et quatre saines) et les métaux seront dosés dans le foie de chaque animal. Ensuite, le croisement des résultats de la détection de la sélection avec ceux de trois méthodes d'association entre les fréquences alléliques et le degré de bioaccumulation hépatique fournira une liste de marqueurs candidats potentiellement situés dans des régions génomiques sous sélection par les métaux. Au cours d'une dernière étape, la technologie de séquençage à haut débit 454 sera utilisée pour amplifier de vastes régions génomiques entourant ces candidats chez 40 individus. Un éventail complet de méthodes computationnelles servira finalement à inférer des hypothèses adaptatives impliquant les polymorphismes non-synonymes découverts dans les séquences codantes de ces régions génomiques.
L'intérêt général de notre démarche est de déterminer si la microévolution d'adaptations peut survenir dans les conditions caractéristiques aux environnements contaminés de Rouyn-Noranda ce qui permet de détailler les impacts concrets des activités minières sur les populations sauvages de perchaudes. En plus de bonifier notre compréhension des effets des métaux sur la dynamique génétique des populations, l'identification de nouveaux gènes adaptatifs présente un intérêt fondamental pour l'étude de l'évolution, en particulier pour la branche de l'écotoxicologie évolutive. Ces informations pourraient, dans un sens plus large, contribuer à étoffer plusieurs modèles évolutifs et alimenter le débat portant sur la vitesse avec laquelle .l'évolution peut agir. Finalement, l'intérêt appliqué de l'étude de l'évolution face à la contamination est de contribuer à définir les paramètres des stratégies de conservation de la biodiversité des populations animales affectées par la pollution.
Chapitre 1 Evolutionary change driven by metal exposure
as revealed by coding SNP genome scan in wild yellow
Résumé français
La pollution provenant des activités humaines peut potentiellement provoquer la microévolution d'adaptations chez les populations animales et végétales sauvages. Cette étude concerne la découverte et la caractérisation de polymorphismes fonctionnels candidats chez des populations de perchaudes (Perca flavescens) chroniquement exposées à la contamination par les métaux. Tout d'abord, notre méthodologie emploie un balayage transcriptomique qui contraste les séquences de novo de groupes de poissons provenant de lacs sains (n = 16) et contaminés (n = 16). Grâce à ce balayage, 48 SNPs candidats ont été génotypes et des mesures de bioaccumulation métallique ont été saisies chez 1052 perchaudes provenant de 10 populations différentes. L'analyse de ces données a permis de discriminer trois SNPs potentiellement sous sélection directionnelle et dont les fréquences alléliques sont associées aux niveaux populationnels moyens de contamination d'après deux méthodes de régression complémentaires. Les régions génétiques étendues flanquant ces candidats ont été l'objet d'une expérience de séquençage à haut débit. L'analyse des séquences produites a permis l'identification de substitutions d'acides aminés non-synonymes qui peuvent théoriquement être adaptatives, surtout dans des conditions de contamination au cadmium. En se basant sur les fonctions des gènes affectés par les polymorphismes, nous proposons que la divergence adaptative entre les perchaudes saines et contaminées indique que les populations contaminées ont pu être sélectionnées pour l'accélération de leur cycle vital (voie métabolique de p53, gène de la cycline Gl) et pour la mitigation d'effets délétères au niveau de la mémorisation spatio-temporelle (voie métabolique de la potentialisation post-synaptique à long terme; gène de la sous-unité 2 de la NADH déshydrogénase). Conformément à l'attendu évolutif chez les populations stressées et énergiquement limitées, les perchaudes adaptées auraient peu d'aptitude à réparer les dommages causés par les métaux pour plutôt allouer leur énergie vers la croissance. Dans le même sens, l'évolution semblerait favoriser des adaptations à faible coût énergétique pour la mitigation des impacts néfastes des contaminants plutôt qu'une stratégie de détoxication coûteuse.
Introduction
The impact of human activities is global and affects all ecosystems to some extent (Vitousek et al, 1997). Airborne pollution is of particular concern as it spreads across the planet, forcing the need to address the associated negative impacts through world-wide cooperation (Akimoto, 2003). To this end, thorough documentation of the dynamics and consequences of pollution in an ecological framework is crucial (Chapman, 2002).
Moreover, it is well documented that certain classes of anthropogenic pollutants pose various fitness issues to wild populations, potentially resulting in strong directional selection pressure (Posthuma & Van straalen, 1993). In such cases, microevolution can lead to local adaptation to new environmental conditions (Hendry & Kinnison, 2001; Reznick & Ghalambor, 2001). Instances of human induced rapid evolution are indeed reported in a growing number of studies (reviewed in Smith & Bernatchez, 2008; Tobler et al, 2010; Williams & Oleksiak, 201 lb). However, although understanding the underlying evolutionary mechanisms is key to ecological risk assessment (ERA) as well as restoration strategies , they are still poorly understood (van Straalen & Timmermans, 2002).
The nascent field of evolutionary ecotoxicology precisely seeks this knowledge and is increasingly empowered by the fast paced "-omics" technological improvements (Rokas & Abbot, 2009). The discipline is thus in the midst of a paradigm shift that tends to focus studies on functional targets of pollution-driven selection in wild populations. Genome and transcriptome wide analyses by means of next generation sequencing (NGS) are the main drivers of new approaches allowing large-scale marker mining without the need for prior target candidates.Yet, few studies to date have bridged the gap between outlier detection and functional characterization of putative targets.
The region of Rouyn-Noranda (Québec, Canada) presents a highly relevant context to study local environmental impacts of the mining industry. For the last 85 years (since 1927), local dominant winds have blown metal emissions from a copper smelter towards the north-east, creating a polymetallic gradient of contamination in nearby lakes (Couillard et al, 2004). Moreover, the cadmium (Cd) and copper (Cu) that deposit in lacustrine environments are responsible for profound, well documented ecosystemic perturbations of the bacterial
communities (Laplante & Derome, 2011), benthic niche (Kovecses et al, 2005), planktonic communities (Cattaneo et al, 2008) and freshwater molluscs (Perceval et al, 2006).
Numerous studies have provided evidences that indigenous yellow perch populations suffer from these alterations (Sherwood et al, 2002; Kovecses et al, 2005). Notably, perch bioaccumulate metals in various tissues, making them prone to several physiological and metabolic impairments that may affect their fitness in natural environments (Couture et al., 2008a; Couture & Pyle, 2008; Couture et al., 2008b; Pyle et al, 2008). Bourret et al. (2008) reported a negative correlation between Cd contamination and genetic diversity, thus revealing evolutionary consequences of metal contamination in these populations. Previous studies did not show that yellow perch from contaminated environments have superior detoxification capacities (Campbell et al, 2005). Indeed, when resources are scarce in contaminated environments, detoxification may not be the first trait to evolve due to its high energetic cost as a general defense mechanism (Sibly & Calow, 1989; Posthuma & Van straalen, 1993). Instead, especially if mortality is high, traits allowing for fast life-cycle completion may be more important targets of selection (Sibly & Calow, 1989). Metal-contaminated yellow perch from the Rouyn-Noranda area may have taken this evolutionary trajectory since they display various negative physiological effects while tending to grow fast, reproduce and die at early age (Couture & Pyle, 2008).
This study aims to identify putative adaptive polymorphisms in nucleotide sequences and to set the basis for in silico functional characterization of metal contamination-induced selection in wild yellow perch populations. This was achieved following a four-step approach: i) perform a transcriptome scan from de novo sequencing to identify putative candidate SNPs located in annotated genes; ii) develop SNP assays for a subset of these markers to genotype 1052 individuals from 10 populations spanning a wide range of Cd and Cu contamination; iii) assess the association between allele frequency of outliers and mean liver metal concentrations using different regression methods; iv) resequence large amplicons to investigate the nature and functional basis of polymorphisms surrounding these outliers.
Material and methods
Sampling
Six lakes were selected for their varying degree of Cd and Cu contamination and previous knowledge on the system (Bourret et al, 2008; Pierron et al, 2009) (Fig. 1). Sampling was conducted in June and July 2010 using beach seine nets in shallow waters. A minimum of 50 yellow perch were sampled in each lake (Table 1). Each fish was measured, weighed and sexed. Fin clips were collected and stored in 95% EtOH for subsequent DNA extraction. Since it is the most suitable organ to evaluate bioaccumulation of Cd and Cu in these populations (Giguere et al, 2004), liver was sampled and snap-frozen in liquid nitrogen within 15 min following sacrifice. In addition, samples from Bourret et al. (2008) collected in 2005 were also used for genotyping. These provided samples for four additional lakes (total number of lakes = 10) and provided two temporal replicates for six lakes.
Metal quantification
Fragments of liver samples were freeze-dried in 1.5 mL tubes that had previously been washed in 15% HNO3 baths to remove any trace metals. Cold digestion was conducted according to Pierron et al. (2009). Certified reference material provided by the National Research Council of Canada (TORT2) and blanks were also included for certifying analytical accuracy and monitoring metal recovery. Concentrations of Cd and Cu were measured using an inductively coupled plasma mass spectrometer (ICP-MS, Thermo Elemental, Model X-7). TORT-2 and blank samples were analyzed every 12 samples to control for contamination and assess reproducibility.
Coding gene SNP development and genotyping
We used whole transcriptome sequences from Pierron et al. (2011) for SNP identification. Briefly, total RNA was extracted from 16 individuals equally sampled in clean lakes Opasatica and Adeline as well as 16 individuals from contaminated lakes Dufault and
Marlon. The retrotranscribed cDNA was individually marked using MIT tags and the 32 individuals were sequenced with a Roche GS-FLX 454 DNA Sequencer. CLC Genomics Workbench 3.7 was used to identify SNPs using the Neighborhood Quality Standard method (NQS; Brockman 2008). The Beaumont and Nichols (1996) selection detection method implemented in Arlequin 3.5 (Excoffier & Lischer, 2010) was used to identify putative targets of selection between clean and contaminated populations. All outliers potentially under directional selection and a subset of putative neutral markers were both retained for subsequent genotyping in large numbers of individuals from all studied populations.
A total of 1052 individuals were genotyped by KBioscience (Hoddeson, UK, http://www.kbioscience.co.uk) who also developed the genotyping assays (48 validated polymorphic markers out of 87 genotyped and 130 submitted; see Results) using the KASP chemistry (Annex 1 for genotyping primers sequences) which is based on the specific hybridization of fluorescence-marked primers on SNPs. Severeal factors can explain the fact that only 48 out of 130 markers identified from cDNA sequence data were validated as true SNPs in genomic DNA, including alignment of paralogs, occurence of exon-exon boudaries and PCR efficiency(Sauvage et al, 2012).
Candidate gene library preparation for large amplicon 454 sequencing
Reference genomes from the phylogenetically closest relatives of Perca flavescens were retrieved from NCBI (www.ncbi.nlm.nih.gov) and Ensembl Genome Browser (Flicek et al, 2012; http://useast.ensembl.org) in order to build scaffolds on which the transcriptomic sequences of Pierron et al. (2011) were aligned with CLC. These scaffolds were used for primer design (Primer3 vO.4.0; Rozen & Skaletsky, 2000; Suppl. mat 1 for primer sequences ) in order to amplify and analyse further three genes that stood out as being the most relevant targets based on outlier and association tests (see below). Using an adapted version of the approach described by Jacobsen et al. (2012), the long-range PCR chemistry (Invitrogen Platinum® Taq DNA Polymerase High Fidelity kit) was used to amplify large