Dissolution biologique des phosphates : Interaction
bactéries - mycorhizes
Thèse
Salma Taktek
Doctorat en microbiologie agroalimentaire
Philosophiae doctor (Ph.D.)
Québec, Canada
Résumé
Une énorme partie du phosphore (P) soluble ajouté sous forme d’engrais chimiques et de fumiers précipite dans le sol et devient non disponible aux plantes. Par ailleurs, l’utilisation excessive des engrais chimiques n’est pas compatible avec l’agriculture moderne qui se veut durable, ni avec l’agriculture biologique. De plus, ces pratiques ont été effectuées sans tenir compte de la microflore présente au niveau de la mycorhizosphère ce qui conduit à des applications pouvant être onéreuses et néfastes. En effet, les microorganismes bénéfiques du sol, notamment les bactéries solubilisant les phosphates (BSP) et les champignons mycorhiziens arbusculaires (CMA), ont une influence importante sur la fertilité des sols et la productivité végétale.
Une nouvelle approche basée sur le piégeage des BSP au niveau de l’hyphosphère du CMA
Rhizophagus irregularis (Ri) DAOM 197198, préalablement inoculé avec des suspensions
de microorganismes telluriques, a permis d’isoler des BSP compétentes capables de dissoudre efficacement le phosphate de roche (PR) d’origine ignée en milieu liquide. Les travaux ont ensuite permis de prouver l’importance du synergisme entre les hyphobactéries (Burkholderia anthina Ba8 et Rhizobium miluonense Rm3) et le mycélium extraracinaire des CMA Ri dans l’amélioration de la solubilisation des phosphates in vitro. L’étude approfondie des mécanismes qui pourraient être impliquées dans cette interaction montre que les hyphobactéries, principalement la souche B. anthina Ba8, adhèrent fortement à la surface des hyphes et aussi à celle du PR. Il est fortement probable que les interactions décrites ainsi que les caractéristiques bénéfiques aux plantes exprimées par les BSP sont responsables de l’amélioration de la croissance, de la nutrition phosphatée et du rendement en matière fraîche et sèche chez le maïs cultivé en serre, coinoculé avec les BSP et les CMA Ri et fertilisé avec le superphosphate ou le PR du Québec.
Abstract
Soluble phosphorus (P) fertilizers added to soil rapidly precipitate, forming sparingly soluble phosphates, not available to plants. Furthermore, the excessive use of chemical fertilizers to compensate soil P deficiency is not considered sustainable and it leads to costly and potentially harmful applications. Many reports confirmed that beneficial soil microorganisms, including phosphate-solubilizing bacteria (PSB), have a significant influence on soil fertility and crop productivity. Indeed, PSB can also improve phosphate rock (PR) efficiency when directly applied to soil. However, most published works on PSB overlooked the possible interaction between PSB and arbuscular mycorrhizal fungi (AMF), which are ubiquitous in cultivated plants.
A new approach based on the trapping of PSB strongly attached to the hyphosphere of AMF Rhizophagus irregularis (Ri) DAOM 197198, previously inoculated with microbial soil suspensions was developed to isolate relevant PSB able to mobilize P from a low reactive igneous PR more efficiently than those directly isolated from the same rhizosphere soil samples. An in vitro study demonstrated that the synergism between hyphobacteria (Burkholderia anthina Ba8 and Rhizobium miluonense Rm3) and Ri hyphae highly improved the solubilisation of PR. Our results go beyond the existing studies and showed specific mechanisms involved on PSB-AMF interactions. Indeed, hyphobacteria, mainly B.
anthina Ba8, strongly adhere to Ri hyphal surfaces and PR particles forming a structured
biofilm. Under greenhouse conditions, the direct application of PSB and AMF Ri as biostimulants for sustainable corn production showed that these beneficial microorganisms improve growth and P uptake of corn fertilized with superphosphate or Quebec PR.
Table des matières
Résumé ... iii
Abstract ... v
Table des matières ... vii
Liste des tableaux ... xi
Liste des figures ... xiii
Liste des abréviations ... xv
Remerciements ... xix
Avant-propos ... xxi
Chapitre 1 : Introduction et revue de littérature ... 1
1.1. La vie en symbiose des plantes ... 2
1.2. Les mycorhizes ... 2
1.2.1. Généralités ... 2
1.2.2. Les endomycorhizes ou mycorhizes arbusculaires ... 4
1.2.3. Rôles des symbioses mycorhiziennes ... 5
1.3. Le phosphore et ses différentes formes dans le sol ... 6
1.3.1. Importance mondiale du phosphore ... 6
1.3.2. Cycle du phosphore et ses différentes formes ... 7
1.4. Mobilisation du phosphore dans le sol ... 9
1.4.1. La désagrégation ... 10
1.4.2. La solubilisation ... 10
1.4.3. La minéralisation ... 11
1.4.4. L’immobilisation ... 11
1.5. Les biofilms associés à la mycorhizosphère ... 12
1.5.1. Les biofilms : définition, mécanisme de formation, structure et habitats ... 12
1.5.2. Les biofilms associés aux plantes ... 14
1.5.2.1. Implication des mycorhizes dans la croissance des bactéries ... 15
1.5.2.2. Avantages écologiques des biofilms ... 19
1.6. Hypothèses et objectifs ... 20
1.6.1. Hypothèses ... 20
1.6.2. Objectifs ... 21
Chapitre 2 : Trapping of phosphate solubilizing bacteria on hyphae of
the arbuscular mycorrhizal fungus Rhizophagus irregularis DAOM
197198 ... 23
Résumé ... 24
Abstract ... 25
2.1. Introduction ... 26
2.2.3. Isolation of PSB strongly attached to hyphae or directly from the soil ... 29
2.2.5. Identification of PSB ... 31
2.2.6. Identification of organic acids ... 31
2.2.7. Siderophores production and organic phosphate mineralization ... 32
2.2.8. Phytase activity ... 33
2.2.9. Interaction between PSB and AMF: phosphate solubilization and pH changes . 33 2.2.10. Statistical analysis ... 34
2.3. Results ... 35
2.3.1. Isolation and identification of bacteria ... 35
2.3.2. Mineral phosphate solubilization ability of PSB and organic acids production . 38 2.3.3. Siderophores production and organic phosphate mineralization ... 43
2.3.4. PSB and AMF interaction ... 45
2.4. Discussion ... 47
Acknowledgements ... 51
References ... 52
Supplementary material ... 56
Chapitre 3 : Solubilization of igneous phosphate rock by biofilm forming
mycorrhizospheric bacteria ... 61
Résumé ... 62
Abstract ... 63
3.1. Introduction ... 64
3.2. Materials and methods ... 65
3.2.1. Strains and mycorrhizal fungus ... 65
3.2.2. Evaluation of bacterial biofilm formed on sparingly soluble phosphates ... 66
3.2.2.1. Crystal violet assay ... 66
3.2.2.2. Exopolysaccharides assay ... 67
3.2.2.3. Fluorescein diacetate assay ... 67
3.2.2.4. Quebec phosphate rock solubilization under biofilm formation ... 68
3.2.3. Fluorescent labeling and confocal laser scanning microscopy ... 68
3.2.4. Scanning electron microscopy ... 69
3.2.4.1. Bacterial attachment to abiotic surface ... 69
3.2.4.2. Bacterial attachment to biotic surface ... 69
3.2.5. Statistical analysis ... 70
3.3. Results ... 70
3.3.1. Biofilm quantification ... 70
3.3.2. Phosphate solubilization under biofilm formation ... 72
3.3.3. CLSM and SEM analysis ... 75
3.4. Discussion ... 78
Chapitre 4 : Phosphate de roche et bioinoculants pour une agriculture
durable ... 87
Résumé ... 88
4.1. Introduction ... 89
4.2. Matériel et méthodes ... 90
4.2.1. Caractérisation des phénotypes PGPR des BSP ... 90
4.2.1.1. Production de l’acide indole acétique ... 90
4.2.1.2. Détermination de l’élongation racinaire du maïs ... 90
4.2.1.3. Tests de motilité ... 91
4.2.1.4. Tests d’antagonisme ... 91
4.2.1.5. Test de production de l’acide cyanhydrique (HCN) ... 91
4.2.1.6. Test de production des chitinases ... 92
4.2.1.7. Analyse statistique ... 92 4.2.2. Essai en serre ... 92 4.2.2.1. Substrat utilisé ... 92 4.2.2.2. Dispositif expérimental ... 93 4.2.2.3. Paramètres mesurés ... 94 4.2.2.4. Analyse statistique ... 94 4.3. Résultats ... 95 4.3.1. Caractérisation PGPR des BSP ... 95 4.3.2. Essai en serre ... 96 4.3. Discussion ... 104 Références ... 110 Matériel supplémentaire ... 115
Chapitre 5 : Conclusion générale ... 119
Liste des tableaux
Tableau 1.1. Exemples de quelques bactéries bénéfiques dans la mycorhizosphère……...15 Table 2.1. Chemical composition and reactivity of Quebec phosphate rock………...28 Table 2.2. Genera to which belong the 51 selected PSB trapped to AMF hyphae or isolated
from the rhizosphere from 13 sampling sites.………...…....36
Table 2.3. Identification of the selected PSB isolates by sequencing of the 16S rDNA…..37 Table 2.4. Organic acid production by selected PSB strongly attached to AMF hyphae
(Hyphobacteria) or isolated from the mycorrhizosphere (Rhizobacteria) on NBRIP medium containing Quebec PR or hydroxyapatite after 7 days of incubation……..42
Table 2.5. Activity of enzymes involved in organic phosphate mineralization…………...44 Table 2.S1. Chemical characteristics of mycorrhizospheric soil samples collected from 13
sites in Quebec and used for the isolation of PSB...………56
Table 2.S2. Selection of PSB used in this study...57 Table 2.S3. Identity, siderophores production and phosphatases activity of selected
PSB...58
Table 3.1. Evaluation of biofilm formation by PSB in the presence of Quebec PR or
hydroxyapatite using three different methods………...71
Table 3.2. Organic acids production by microbial biofilm forming PSB grown in microtiter
plates………..74
Tableau 4.1. Quelques caractéristiques bénéfiques aux plantes chez les BSP…………....95 Tableau 4.2. Effet antifongique des BSP vis-à-vis de quelques champignons
pathogènes……….96
Tableau 4.3. Caractéristiques du sol de jardin utilisé………..97 Tableau 4.4. Analyse de la variance de la matière fraîche et sèche (g plant-1) et du
pourcentage de mycorhization (%) des plants de maïs fourrager (Zea mays cv. FOCUS) fertilisés par le PR du Québec ou le SP et coinoculés avec les BSP et
Ri………...………97
Tableau 4.5. Effet de l’inoculation avec les BSP et des différents traitements de
fertilisation avec le superphosphate (SP) ou le phosphate de roche du Québec (PR), sur la masse fraîche et sèche des parties aériennes et le pourcentage de
mycorhization des plants de maïs fourrager (Zea mays cv. FOCUS) mycorhizés avec le Ri………...……99
Tableau 4.6. Structure de la matrice obtenue à la suite de l’analyse factorielle des dix
éléments chimiques absorbés au niveau des parties aériennes des plants de maïs fourrager..………....100
Tableau 4.7. Analyse de la variance des deux facteurs représentant les dix éléments
chimiques absorbés par les plants de maïs fourrager (Zea mays cv. FOCUS) fertilisés par le PR du Québec ou le SP et inoculés avec les BSP et Ri………...101
Tableau 4.8. Effet de l’inoculation des plants de maïs fourrager (Zea mays cv. FOCUS)
avec les BSP, Ri et les différents traitements de fertilisation sur les dix éléments chimiques absorbés par les parties aériennes………...………...103
Annexe 4.1. Quelques activités bénéfiques aux plantes chez les BSP……….…………..115 Annexe 4.2. Analyse de la variance des deux facteurs représentant les dix éléments
chimiques analysés au niveau des organes aériens des plants de maïs fourrager (Zea
mays cv, FOCUS) fertilisées par la PR du Québec ou le superphosphate et inoculés
avec les BSP et Ri………...117
Annexe 4.3. Effet de l’inoculation des plants de maïs fourrager (Zea mays cv. FOCUS)
avec les BSP, Ri et les différents traitements de fertilisation sur les dix éléments
chimiques analysés au niveau des parties
Liste des figures
Figure 1.1 : Les différents types d’associations mycorhiziennes d’après (Selosse and Le
Tacon 1998) repris par (F. Halle dans le livre Aux origines des plantes, Ed. Fayard 2008)………3
Figure 1.2 : Morphologie des mycorhizes arbusculaires (Fortin et al. 2008)………5 Figure 1.3 : Les cinq étapes du développement d’un biofilm sur une surface dure.
Étape 1 : attachement initial; étape 2 : attachement irréversible; étape 3 : apparition et maturation I du biofilm; étape 4 : maturation II du biofilm; étape 5 : dispersion. Les photomicrographies, présentées toutes à la même échelle, sont celles d’un biofilm de Pseudomonas aeruginosa en développement (Monroe D, 2007)………12
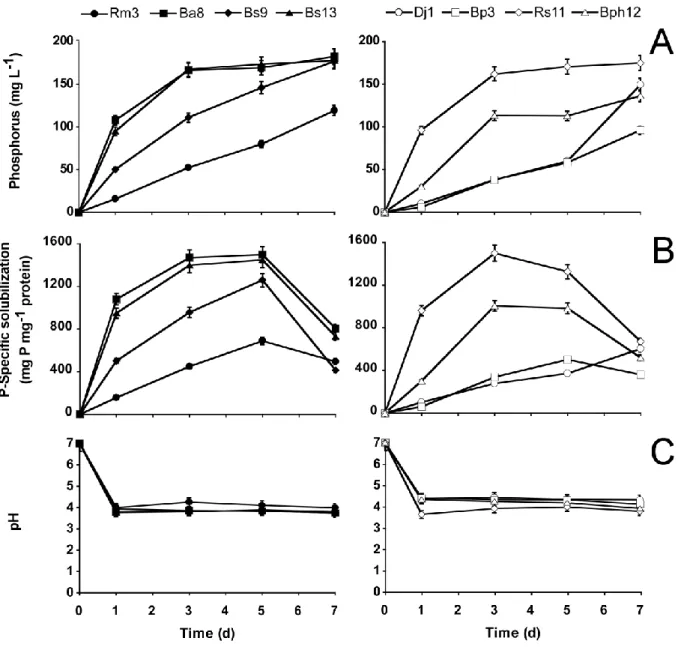
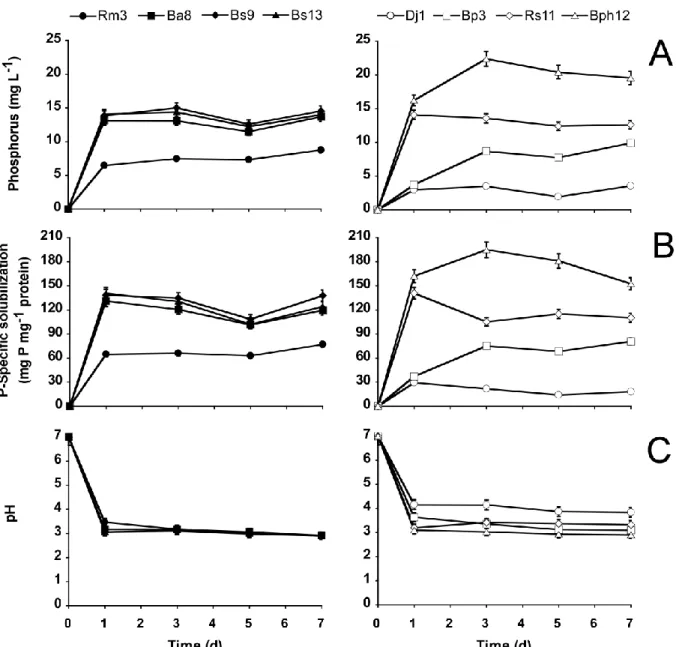
Figure 2.1 : (A) Soluble phosphate ; (B) specific phosphate solubilization activity, LSD0.05
values: d1 = 63.98, d3 =177.78, d5 =110.80, d7 = 55.28 mg P mg-1 protein; and (C)
pH variation in the supernatant of PSB growing on NBRIP medium containing hydroxyapatite. Phosphate solubilizing hyphobacteria are shown on the left and rhizobacteria on the right. PSB identity is described in Table 2.3. Error bars indicate standard deviation of means from three independent assays………40
Figure 2.2 : (A) Soluble phosphate; (B) specific phosphate solubilization activity LSD0.05
values: d1 = 22.75, d3 = 23.61, d5 = 27.55, d7 = 26.59 mg P mg-1 protein; and (C)
pH variation of PSB growing on NBRIP medium containing Quebec phosphate rock. Phosphate solubilizing hyphobacteria are shown on the left and rhizobacteria on the right. PSB identity is described in Table 2.3. Error bars indicate standard deviation of means from three independent assays………...41
Figure 2.3 : Production of siderophores by PSB in AT minimum medium after 5 days of
incubation. Means followed by the same letter are not significantly different according to Fisher’s protected LSD test (P ≤ 0.05). Error bars are standard deviations of the means from three independent assays. Table 2.3 shows PSB identity………...43
Figure 2.4 : Soluble phosphate and pH variation in the minimal medium extract obtained
from the distal compartment containing AMF Rhizophagus irregularis (Ri), Ri plus
Rhizobium miluonense (Ri+Rm3), Ri plus Burkholderia anthina (Ri+Ba8), Ri plus Rahnella sp. (Ri+Rs11) and Ri plus Burkholderia phenazinium (Ri+Bph12) after 6
weeks of dual incubation. Means followed by the same letter are not significantly different according to Fisher’s protected LSD test (P ≤ 0.05). Error bars are standard deviations of the means from three independent assays………46
Figure 3.1 : P-solubilization by PSB biofilms (A) and pH values (B) after 5 days of
incubation in liquid modified NBRIP containing 5 g L-1 Quebec PR or
hydroxyapatite in microtiter plates. Significant differences (LSD test; P ≤ 0.05) are indicated by different letters. Bars are standard deviations of the mean (n=12)………..………..73
Figure 3.2 : Confocal laser scanning micrograph of PSB. Figures show average projections
of CLSM images of two hyphobacteria : Rhizobium miluonense Rm3 and
Burkholderia anthina Ba8 and two rhizobacteria : Rahnella sp. Rs11 and Burkholderia phenazinium Bph12 through FDA stained biofilms (green
fluorescence) on Quebec PR (left panel) or hydroxyapatite (right panel) after 5 days of incubation. Scale bars (white) correspond to 20 µm, magnification 630×. The confocal images were taken along z-sections into the scaffolds and shown as 2D pictures composed by ZEN 2011 Software……….…………..76
Figure 3.3 : Scanning electron micrograph of Burkholderia anthina Ba8 bacterial matrix
formed on Quebec PR (A) or hydroxyapatite (B) after 5 days of incubation. White arrows indicate individual cells scattered on the surface of hydroxyapatite particles………...…..77
Figure 3.4 : Scanning electron micrograph of Burkholderia anthina Ba8 bacterial matrix
formed on Rhizophagus irregularis (Ri) hyphae using Quebec PR as sole P source after 6 weeks of dual culture……….……77
Liste des abréviations
AIA : Acide indole acétique ANOVA : Analyse de la variance
BSP : Bactérie(s) solubilisant les phosphates
CMA : Champignon(s) mycorhizien(s) arbusculaire(s) DO : Densité optique
HCN : Acide cyanhydrique
LSD : Test de différence significative minimale de Fisher
NBRIP : de l’anglais « National botanical institute’s phosphate growth medium » P : Phosphore
PCR : Réaction de polymérisation en chaîne
PGPR : de l’anglais « Plant Growth Promoting Rhizobacteria » PR : Phosphate de roche
Ri : Rhizophagus irregularis
TSA : de l’anglais « Tryptic Soy Agar » TSB : de l’anglais « Tryptic Soy Broth »
À la mémoire de mon grand-père, Ali. À mon ange Rahma, envolée au Paradis. À mon aimable famille du Canada à l’au-delà de l’Atlantique, Tunisie.
Remerciements
Il y a maintenant un peu plus de quatre ans que j’ai traversé l’Atlantique pour entreprendre le travail présenté dans cette thèse de doctorat. Arrivée à Québec, j’étais chaleureusement accueillie par Dr. Antoun et son épouse Dina, qui m’ont aidé, guidé et fait visiter l’université Laval et ses environs. Après tout cela, je me suis sentie très motivée avec la certitude que les raisons qui m’avaient amenées ici étaient les bonnes. À présent, je me rends compte que ma thèse a en effet été un voyage que j’ai commencé avec motivation, parcouru avec enthousiasme et terminé avec certitude. Je veux donc prendre ces quelques lignes pour remercier ceux qui ont contribués à mon avancement et qui sans leurs aides ce travail n’aurait pas pu se réaliser.
En premier lieu, je dois remercier Dr. Hani Antoun qui a conservé sa disponibilité dont il a fait preuve le jour de mon arrivée et qui m’a transmis son efficacité et sa rigueur scientifique. Je l’en remercie sincèrement.
Je souhaite également remercier mon co-directeur, Dr. Yves Piché pour m’avoir transmis ses connaissances scientifiques sur la microscopie et aussi pour m’avoir encouragé à persévérer pour mettre ce travail sur pied.
Mes vifs remerciements s’adressent également à Dr. J.-André Fortin. Il a été un catalyseur et une personne ressource pour le cheminement de mes travaux de doctorat. Je lui suis redevable pour son soutien constant, son aide et sa disponibilité.
Je souhaite aussi remercier Dr. Marc St-Arnaud pour avoir accepté d’être membre de mon comité aviseur et pour avoir été toujours prêt à partager son savoir pour me faire des commentaires constructifs très utiles.
Merci également au Dre. Valérie Gravel pour avoir accepté d'évaluer ma thèse et aussi pour ses corrections et commentaires pertinents.
Par ailleurs, je tiens à remercier Martin Trépanier pour son soutien, son aide et son amitié. Sa détermination et sa bonne humeur ont été une source d'encouragement et d’appui.
Je remercie également Marie-Claude Julien pour ses conseils et son agréable amitié. Merci aussi pour m’avoir offert l’opportunité de travailler avec elle plusieurs sessions et d’approfondir mes connaissances en microbiologie générale.
thèse. Les anciens, Paola et Mélissa pour leur aide, encouragements et surtout leur amitié éternelle, Antoine pour sa bonne humeur et ses bras quand j’avais besoin d’aide et Joseph pour ses belles discussions. Les nouveaux, Vicky pour sa collaboration, Héla pour son amitié et ses encouragements, Martine, Andrée et Amélie pour leur agréable compagnie. J'ai également une pensée particulière pour les stagiaires Jean Sébastien, Marc Antoine, Stéphanie et Laetitia qui ont contribué au bon déroulement de certains de mes travaux. Je leur souhaite également beaucoup de succès dans leur carrière. Je remercie Richard Janvier et son équipe de la plateforme de l’institut de biologie intégrative et des systèmes (IBIS) pour leur aide, disponibilité et leur soutien technique lors des travaux reliés à la microscopie. Jean Martin mérite également mes plus sincères remerciements pour son implication dans mon travail. Mes remerciements s’adressent aussi à Gaétan Daigle pour ses coups de main lors des analyses statistiques. Merci à tous les membres de l’Envirotron et à l’équipe des serres pour leur aide et collaboration.
La réalisation de ce travail a été possible grâce aux subventions de recherches du Fonds de Recherche du Québec Nature et Technologies et du Conseil de Recherches en Sciences Naturelles et en Génie du Canada. Je remercie également les organismes suivants pour leur appui financier sous la forme de bourses pour participer à des congrès internationaux et avoir l’opportunité de présenter mes travaux de recherche : le centre sève, le centre de recherche en horticulture ainsi que le centre d’étude sur la forêt.
Bien entendu, je tiens à remercier ma famille, mes proches et mes ami(e)s pour leur inestimable soutien et leurs encouragements. Je tiens à faire des remerciements à mes parents qui malgré les distances ont su être présents tout au long de mon doctorat. Un grand merci à Habib, mon mari, pour son soutien moral, son temps consacré pour m’aider ainsi que ses suggestions lors de l’élaboration de ce travail. Merci de m’avoir encouragé à continuer de croire que je suis capable et surtout de m’avoir offert une ambiance propice à la réussite. Finalement, une pensée particulière à Ahmed, mon garçon, pour son sourire, sa joie de vivre et surtout pour avoir été un enfant facile me permettant ainsi de me rendre jusqu’au terme de ce projet.
Avant-propos
Les travaux de recherche réalisés durant cette thèse ont été présentés lors de plusieurs conférences nationales et internationales et feront l'objet de trois publications dans des revues avec comité de lecture. Les chapitres deux et trois de cette thèse sont sous forme d’articles scientifiques. La contribution des coauteurs est détaillée ci-dessous :
Chapitre 2 - Trapping of phosphate solubilizing bacteria on hyphae of the arbuscular mycorrhizal fungus Glomus irregulare DAOM 197198
Salma Taktek, Martin Trépanier, Paola Magallon, Marc St-Arnaud, Yves Piché, J.-André Fortin et Hani Antoun
La répartition des contributions des co-auteurs est la suivante : J’ai rédigé l’article sous la direction de Dr. Antoun et Dr. Piché. J’ai également réalisé la quasi-totalité des manipulations. Martin Trépanier a contribué à l’identification des acides organiques. Paola Magallon a participé à l’isolement des souches solubilisant les phosphates. Marc St-Arnaud et J.-André Fortin ont révisé et corrigé le manuscrit. Cet article est soumis à Soil Biology &
Biochemistry.
Chapitre 3 - Solubilization of igneous phosphate rock by biofilm forming mycorrhizospheric bacteria
Salma Taktek, Marc St-Arnaud, Yves Piché, J.-André Fortin et Hani Antoun
La répartition des contributions des co-auteurs est la suivante : J’ai rédigé l’article sous la direction de Dr. Antoun et Dr. Piché. J’ai également réalisé la totalité des manipulations. Marc St-Arnaud et J.-André Fortin ont révisé et corrigé le manuscrit. Cet article sera soumis à FEMS Microbiology Ecology.
Article en préparation
Can igneous phosphate rock, phosphate solubilizing bacteria and arbuscular mycorrhizal fungi substitute chemical fertilizers?
Conférences et symposiums
Salma Taktek, Martin Trépanier, Hani Antoun. Mycorrhiza competent bacteria for
sustainable use of phosphates in agriculture. 15th International Symposium on Microbial Ecology, 24 - 29 août 2014, Séoul, Corée du Sud.
Salma Taktek, Yves Piché, Hani Antoun. Biofilm formation and phosphorus mobilization
by mycorrhizosphere bacteria. International Union of Microbiological Societies, 27 juillet - 1 août 2014, Montréal, Canada.
Salma Taktek, Martin Trépanier, Hani Antoun. Improving the availability of phosphorus
from a Quebec rock phosphate by using biofilm forming phosphate solubilizing bacteria associated with Glomus irregulare. 63rd Annual Conference of the Canadian Society of Microbiologists, 17 - 20 juin 2013, Ottawa, Canada.
Salma Taktek, Hani Antoun. Amélioration de la disponibilité du phosphore à partir de la
roche phosphatée du Québec par des bactéries solubilisant les phosphates en formant des biofilms. Journée annuelle étudiante du centre de recherche en horticulture, 23 mai 2013, Québec, Canada.
Salma Taktek, Martin Trépanier, Hani Antoun. Une nouvelle méthodologie d'isolement de
bactéries solubilisant les phosphates. Colloque Mycorhizes 2012, 05 octobre 2012, Montréal, Canada.
Salma Taktek, Martin Trépanier, J.-André Fortin et Hani Antoun. Isolements,
identification et caractérisation des bactéries solubilisant les phosphates. Congrès annuel conjoint Association Québécoise des Spécialistes en Sciences du Sol Société Canadienne de la Science du Sol, 03 - 08 juin 2012, Lac-Beauport, Canada.
Salma Taktek, Paola Magallon, Martin Trépanier et Hani Antoun. Dissolution biologique
des phosphates : Interaction bactéries-mycorhizes. Congrès de bactériologie intégrative: symbiose & pathogenèse, 10 - 11 novembre 2011, Québec, Canada.
Chapitre 1 :
1.1. La vie en symbiose des plantes
Dans la nature, la symbiose végétale représente le mode de vie de l’ensemble des organismes. Au cours des dernières années, une multitude de travaux ont clairement démontré l’intérêt scientifique et pratique de ces symbioses pour l’ensemble des végétaux (Fortin et al. 2008). Plus que 90 % des végétaux vivent de façons symbiotiques. Il s’agit d’une association durable entre deux ou plusieurs êtres vivants hétérospécifiques et dont chacun tire bénéfice. Les symbiotes s’aident mutuellement à se nourrir, se protéger ou se reproduire.
Au niveau des sols, les sites privilégiés de multiplication des microorganismes sont la rhizosphère et la mycorhizosphère, zones très riches en nutriments exsudés par les racines et/ou par le mycélium (sucres, acides aminés, acides gras, facteurs de croissance…) (Bonfante and Anca 2009).
1.2. Les mycorhizes
1.2.1. GénéralitésLa majorité des plantes supérieures terrestres vivent en étroite symbiose avec les champignons. Il s’agit d’un phénomène fondamental et universel qui s’élabore au niveau du système racinaire des plantes vasculaires et des bryophytes. Les organes résultant de cette association sont appelés mycorhizes. D’origine gréco-latine, le terme mycorhize (« mukês » pour champignon et « rhiza » pour racine) décrit de nombreuses et diverses associations racine-champignon. Actuellement, on définit donc les mycorhizes comme étant des associations symbiotiques contractées par les racines des végétaux avec certains champignons du sol. On les trouve dans de nombreux environnements et leur succès écologique reflète une forte diversité des capacités génétiques et physiologiques des champignons endophytes (Bonfante and Anca 2009).
Environ 6000 espèces dans le Glomeromycotina, Ascomycotina et Basidiomycotina ont été enregistrées comme mycorhiziennes. Les symbioses mycorhiziennes (353 - 462 Millions d’années) ont joué un rôle crucial dans l’évolution des plantes terrestres, bien que quelques groupes (Chenopodiaceae, Caryophyllaceae, Brassicaceae, Urticaceae) se soient
affranchis tardivement de toute relation avec des champignons symbiotiques (Simon et al. 1993; Selosse and Le Tacon 1998).
L’association racine-champignon est basée sur des profits réciproques et des échanges bilatéraux d’éléments nécessaires au bon développement des deux partenaires. Les symbioses mycorhiziennes jouent un rôle essentiel dans l’absorption des nutriments dans les écosystèmes terrestres : en effet, les champignons, grâce à leur structure mycélienne, fournissent des éléments minéraux et de l’eau à la plante, en échange de squelettes carbonés issus de la photosynthèse.
Figure 1.1 : Les différents types d’associations mycorhiziennes d’après (Selosse and Le
Tacon 1998) repris par (F. Halle dans le livre Aux origines des plantes, Ed. Fayard 2008). Les différents types de mycorhizes (Figure 1.1) existant se distinguent à la fois par les groupes taxonomiques des partenaires symbiotiques impliqués et par les structures typiques formées par la symbiose. La position taxonomique de la plante et des partenaires fongiques définit les types de mycorhizes. Ainsi, les structures générées par l’association mycorhizienne peuvent être classées sur la base de critères écologiques, morphologiques et physiologiques. On distingue plusieurs types de mycorhizes : les endomycorhizes, les ectomycorhizes, et les ectendomycorhizes, ainsi que les mycorhizes arbutoïdes,
monotropoïdes et orchidoïdes. Dans la partie qui suit, nous nous intéresserons particulièrement aux endomycorhizes arbusculaires constituant le type de mycorhizes le plus ancien et ayant coévolué semble-t-il avec les plantes terrestres depuis 460 millions d’années (Redecker et al. 2000; Simon et al. 1993).
1.2.2. Les endomycorhizes ou mycorhizes arbusculaires
Les mycorhizes arbusculaires (MA) constituent la symbiose végétale la plus répandue et la plus ancienne dans la nature, du fait de leur ubiquité et du nombre élevé d’espèces végétales concernées (400 000 espèces connues) (Smith and Read 1997; Garbaye 2013). On rencontre le mutualisme endomycorhizien communément chez les plantes herbacées mais aussi chez un grand nombre d’espèces ligneuses. À ce jour, on dénombre environ 250 espèces de champignons distribuées dans quatre ordres, onze familles et dix-sept genres, capables de former ce type de mycorhizes. Ils appartiennent tous aux gloméromycètes (anciennement appelées Glomales) et ne peuvent être cultivés axéniquement. Ce sont des mycosymbiotes obligatoires dont le cycle biologique dans le sol repose entièrement sur la présence de racines vivantes de la plante hôte.
Les gloméromycètes développent un réseau de filaments mycéliens de type siphonné ou cénotique (Figure 1.2). À ce jour, aucune forme de reproduction sexuée n’a encore été décrite chez les gloméromycètes. Ils se propagent d’une façon végétative principalement
via la formation de grosses spores généralement sphériques de 40 à 500 µm de diamètre
comportant quelques milliers de noyaux (Fortin et al. 2008).
Les mycorhizes arbusculaires se caractérisent par l’absence du manteau fongique autour de la racine. Au contact de la cellule racinaire, l’hyphe forme un appressorium. Le mycélium pénètre dans les cellules racinaires, franchit les parois et repousse le plasmalemme des cellules hôtes sans le traverser (Figure 1.2). Les hyphes passent ensuite de cellule à cellule en direction apicale et progressent également dans les espaces intercellulaires. Dans les cellules corticales, le champignon développe des arbuscules et dans la plupart des cas, des vésicules.
Figure 1.2 : Morphologie des mycorhizes arbusculaires (Fortin et al. 2008). 1.2.3. Rôles des symbioses mycorhiziennes
Read (1991) a écrit qu’indépendamment du type de mycorhizes, si la symbiose mycorhizienne est autant répandue dans le monde végétal, c’est parce qu’elle est bénéfique à la plante sous plusieurs aspects. En effet, les champignons mycorhiziens ont un effet fertilisant et jouent un rôle dans la stabilité du sol (formation d’agrégats). Grâce à leur réseau mycélien extraracinaire qui prospecte l’environnement en trois dimensions, les champignons mycorhiziens fournissent aux plantes l’eau et les nutriments à quantité limitante ou à faible mobilité dans le sol. Et enfin, ils ont un effet protecteur contre les organismes pathogènes et contre les stress hydriques.
Par ailleurs, le mycélium extraracinaire des champignons mycorhiziens arbusculaires (CMA) peut avoir des relations directes ou indirectes avec les différents organismes du sol. En effet, ces mycorhizes sont capables de modifier la physiologie de la plante hôte et l’exsudation racinaire d’où leur action indirecte sur les organismes de la mycorhizosphère lesquels, dans certains cas peuvent être étroitement liés aux hyphes permettant ainsi un échange métabolique direct (Johansson et al. 2004). Les communautés bactériennes associées à la mycorhizosphère sont les plus concernées par l’action des mycorhizes
arbusculaires. Une section détaillée sera donc dédiée ultérieurement à l’étude de l’interaction entre les bactéries et les mycorhizes.
1.3. Le phosphore et ses différentes formes dans le sol
Le phosphore (P) est l’un des éléments importants sur terre. En effet, il s’agit de l’élément essentiel du métabolisme énergétique de toutes les formes de vie. Le P est indispensable pour les cellules vivantes vu qu’il est un composant principal de l’ADN, de l’ARN, de l’ATP et des phospholipides (composant majeur des membranes cellulaires). Il est conséquemment impliqué dans la division cellulaire, la transmission de l’information génétique, le transfert et le stockage d’énergie et aussi dans le système photosynthétique. Toutes ces propriétés font de cet élément l’un des trois macronutriments nécessaires pour la croissance et le développement des plantes (avec l’azote (N) et le potassium (K)), généralement présents dans les engrais chimiques (Liu and Chen 2008).
Le P contrôle de nombreux processus biogéochimiques se produisant dans la biosphère. Pour comprendre l’interaction entre le P, les processus biogéochimiques et d’autres distributions élémentaires, il est nécessaire de comprendre la distribution du P sur la surface terrestre et les processus contrôlant sa distribution (Liu and Chen 2008).
1.3.1. Importance mondiale du phosphore
L’évolution des plantes terrestres a été associée à des activités volcaniques au cours de la période de l'Ordovicien (il y a 450 millions d’années) (Parnell and Foster 2012). Une étude récente a démontré la présence de teneurs assez importantes de P2O5 (allant jusqu’à 0.17 %)
dans des cendres volcaniques en provenance du Japon et des Philippines (Shoji and Takahashi 2002). Des expériences réalisées avec ces cendres (utilisées comme substrat) ont prouvé leur importance en tant que source de P pour la croissance des plantes (Joergensen and Castillo 2001). Par conséquent, on en déduit que la teneur en apatite ignée provenant des cendres volcaniques au niveau de la croûte terrestre a adéquatement permis la croissance et le développement des plantes au fil des siècles.
De nos jours, l’agriculture dépend essentiellement de l’apport régulier d’engrais phosphaté. Cet apport indispensable en P fait de cette ressource non renouvelable un produit non
substituable. L’institut d’études géologiques des États-Unis (USGS) s’avère le seul organisme fournissant des informations sur l’état de ‘santé’ des ressources naturelles dont nous dépendons. D’après l’USGS, les ressources terrestres de P seront complètement épuisées dans 50 à 100 ans et la production mondiale du P connaîtra un pic dans les années à venir (approximativement en 2030) (Cordell et al. 2009).
Le Maroc, la Chine et les États-Unis sont les trois principaux producteurs de phosphate de roche dans le monde. Ils détiennent plus que 80 % des gisements exploitables constitués de réserves et ressources potentielles mais à durée de vie limitée. Ces gisements se répartissent en deux groupes suivant leur origine géologique : les gisements ignés qui résultent de l’intrusion du magma dans les roches cristallines (dépôts volcaniques) et ceux sédimentaires qui tiennent leur origine des innombrables débris d’animaux marins.
Le Canada est pourvu de quelques gisements de phosphate : au Québec, en Ontario, au Nouveau Brunswick et dans la partie du sud de la zone frontalière Alberta – Colombie-Britannique (USGS, World phosphate deposit). On y trouve quelques mines de roches phosphatées notamment Arianne phosphate Inc. qui se consacre au développement d’un projet minier au Québec (Lac à Paul). Les réserves minérales de ce gisement sont d’environ 472 millions de tonnes avec une durée de vie d’environ 26 ans.
L’exploitation minière produit un phosphate brut qui ne convient qu’exceptionnellement à un emploi immédiat comme engrais phosphaté. Généralement, pour obtenir un produit marchand ou phosphate naturel, des procédés d’épierrage, criblage, concassage, broyage, séchage, flottation et calcination s’avèrent indispensables.
Dans ce travail, l’étude du potentiel d’utilisation de l’apatite d’Arianne comme fertilisant en agriculture pourrait remédier aux énormes pertes énergétiques et aux impacts environnementaux majeurs (tel que l’accumulation du gypse) générés par les procédés d’extraction, transformation et livraison à longues distances des phosphates.
1.3.2. Cycle du phosphore et ses différentes formes
La disponibilité du P influe fortement sur le processus par lequel les organismes photosynthétiques fixent le carbone inorganique au niveau de la biomasse cellulaire. Par
conséquent, la connaissance du cycle du P est très importante pour la compréhension du bilan global du carbone ainsi que les différents cycles biogéochimiques.
La flore microbienne du sol a un rôle essentiel dans le cycle du P, car elle établit un lien entre le réservoir de P dans l’environnement vivant et non vivant. Certains microorganismes facilitent, en effet, l’altération, la minéralisation, et la solubilisation des différentes formes de P, rendant l’orthophosphate à la disposition des communautés microbiennes et végétales. Cependant, ces microorganismes contribuent également à l’immobilisation du P, un processus qui diminue la biodisponibilité du P en convertissant les formes solubles réactives du P en formes insolubles. Le mécanisme de participation microbienne dans ces processus varie d’un mécanisme passif à un mécanisme très actif. Le cycle du P englobe de nombreux réservoirs environnementaux vivants ou non vivants ainsi que différentes voies de transport. En suivant le mouvement du P dans l’environnement, l’interaction entre le processus physique et biologique devient apparente. En effet, en plus d’agir comme des réservoirs de P dans l’environnement, les microorganismes contribuent à la transformation du P dans les autres réservoirs, comme dans le sol ou dans les environnements aquatiques environnants.
Le P est un élément peu mobile dans le sol (Holford 1997). Certaines formes de P sont insolubles ou ont une solubilité modérée. Dans la nature, le P existe sous forme d’orthophosphate (PO43-) plutôt que sous sa forme élémentaire puisqu’il réagit facilement
avec l’oxygène. Au niveau du sol, les deux principales formes d’ions phosphatés sont le H2PO4- (condition acide) et le HPO42- (condition alcaline) (Busman et al. 2002).
Le P du sol n’a aucun effet toxique direct sur les humains ou les animaux, mais il peut causer l’eutrophisation des eaux de surface. Ce phénomène désigne un état de déséquilibre caractérisé par une suraccumulation de nutriments minéraux (essentiellement, l’azote sous forme de nitrates ou nitrites et/ou le phosphore sous forme phosphate) au niveau des écosystèmes aquatiques conduisant ainsi à la prolifération excessive d’organismes autotrophes notamment les cyanobactéries et les algues (Correll 1998). Ces organismes ont une haute productivité et des taux respiratoires élevés conduisant à l’anoxie (réduction de l’O2 dissous et biodisponible dans le milieu) d’où la dégradation du milieu aquatique et la
Comme indiqué plus haut, l’utilisation irrationnelle des engrais chimiques en agriculture est souvent ciblée comme la cause principale des teneurs élevées en P au niveau des cours d’eau et des eaux souterraines. Plusieurs remèdes sont proposés afin d’atténuer l’eutrophisation en particulier la réduction des apports agricoles tout en privilégiant les engrais écologiques comme le phosphate naturel.
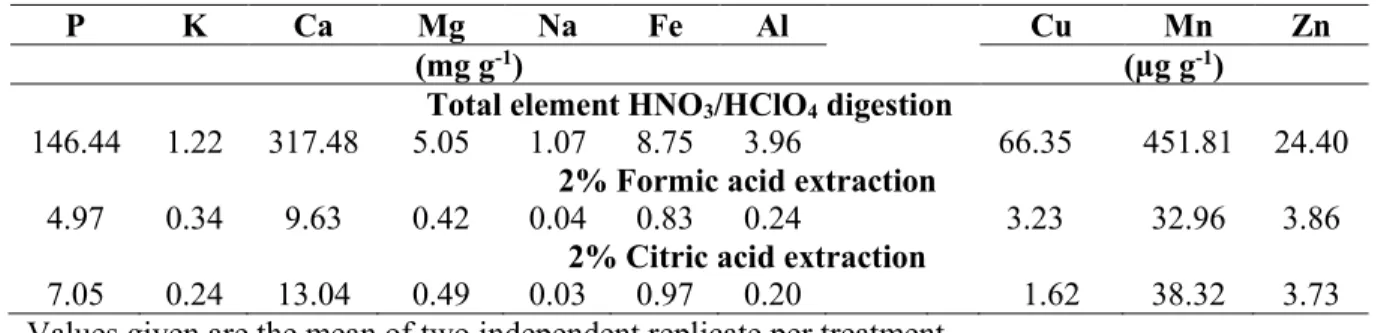
Dans la croûte terrestre, l’abondance du P est de 0.04 à 0.12%, dont la majeure partie est sous sa forme inorganique minérale phosphatée et d’autres composés contenant du P organique. C’est sous cette forme que les groupements anioniques du phosphate forment des complexes tétraédriques en se liant à des cations, substituant ainsi l’arsénate (AsO43-)
ou le vanadanate (VO43-) dans la structure cristalline. L’apatite est la forme la plus
abondante du phosphate minéral, comprenant l’hydroxyapatite, le fluorapatite et le chlorapatite. Ces trois formes d’apatite ont presque la même structure cristalline, mais diffèrent dans leurs proportions relatives d’hydroxyde, de fluorure et de chlorure, chacun étant nommé pour l’anion qui est le plus abondant dans le minéral (Mackey and Paytan 2009).
Dans les sols, le P inorganique est généralement associé à d’autres composés comme le Ca, Fe et Al dont chacun a des caractéristiques de solubilité unique qui détermine la disponibilité des phosphates pour la plante.
Il faut noter que la mobilité et la biodisponibilité des phosphates dans les sols sont principalement limitées par l’adsorption (c’est-à-dire l’adhésion physique ou les liaisons des ions phosphates sur les surfaces d’autres molécules) et l’importance de la flore microbienne qui va transformer le P sous sa forme organique.
1.4. Mobilisation du phosphore dans le sol
Bien que le contenu en P total (inorganique et organique) des sols ne dépasse généralement pas les 0.12%, seulement 0.1% de ce P total existe sous la forme inorganique soluble facilement assimilable par les plantes (Goldstein 1994). Quant au P organique, sa contribution au P total, peut dépasser les 50% dans certains sols. Il peut exister sous plusieurs formes comme les phytates et polyphosphates faiblement biodisponibles et formant des complexes avec des cations, à l’origine de certaines limitations en minéraux
La déficience en P soluble limite la productivité végétale de plusieurs sols agricoles de manière universelle (Arcand and Schneider 2006). Cependant la protection de l’environnement exige l’utilisation de pratiques durables de gestion, faisant usage de peu d’intrants chimiques. Par ailleurs, l’application de fertilisants s’effectue toujours sans tenir compte des mycorhizes et de la microflore présente au niveau de la mycorhizosphère, ce qui conduit à des applications excessives et souvent néfastes. Ainsi, une énorme quantité de P est immobilisée dans les sols, ce qui présente un risque potentiel pour l’environnement si ce sol est transféré par érosion dans les cours d’eau.
Dans cette partie nous nous intéresserons à détailler les différents procédés induits de transformation des phosphates comme la désagrégation, la solubilisation, la minéralisation et l’immobilisation. La composition chimique des minéraux phosphatés dépend de l’ion ou des ions présents au moment de la transformation.
1.4.1. La désagrégation
Dans la nature, certaines roches phosphatées s’altèrent suite à de nombreux processus écologiques. Les processus d’altération sont classés en deux principales catégories : la désagrégation mécanique et chimique.
Dans la désagrégation mécanique, des procédés physiques (incluant l’expansion thermique, la pression, l’action hydraulique, la formation de cristaux de sel, le gel et le dégel…) peuvent causer une détérioration ou encore une fragmentation du matériel rocheux sans modifier sa composition chimique.
En revanche, la désagrégation chimique (incluant divers produits chimiques) cause l’altération de la roche phosphatée en modifiant la structure chimique des minéraux à partir desquels la roche phosphatée est faite. Le processus de l’altération chimique comprend la dissolution, l’hydrolyse, l’hydratation et l’oxydoréduction (Mackey and Paytan 2009).
1.4.2. La solubilisation
La solubilisation microbienne du phosphate joue un rôle important dans la conversion du P insoluble en P soluble. En effet, il a été démontré que certains microorganismes du sol sont impliqués dans la solubilisation des phosphates insolubles. Ces microorganismes bénéficient directement du P biodisponible nécessaire pour leur croissance. De même,
d’autres organismes sont en mesure de profiter du P solubilisé, tels que les champignons et les plantes supérieures.
Notons que ces microorganismes produisent des acides organiques et relâchent des protons, qui à travers leurs groupements carboxyliques, chélatent les cations fixés aux phosphates insolubles ce qui permet de les convertir en formes solubles (mono et dibasiques) (Mackey and Paytan 2009).
1.4.3. La minéralisation
Dans le sol, les plantes et les détritus animaux constituent un énorme réservoir de P organique qui est généralement indisponible pour la plupart des organismes vivants. Pour devenir biodisponible, le P contenu dans la matière organique doit tout d’abord être minéralisé en phosphate.
Le processus de minéralisation, au cours duquel les complexes de P organique sont convertis en minéraux phosphatés, est un processus modulé grâce à l’activité de certaines enzymes microbiennes, notamment les phosphatases qui sont classées en fonction du type des groupements carbonés liés aux phosphates qu’elles clivent.
Les phosphatases ont des exigences spécifiques vis-à-vis de certains substrats. Les catégories les plus courantes des phosphatases microbiennes contribuant à la minéralisation du P comprennent les phosphomonoestérases, les phosphodiestérases, les nucléases, et les nucléotidases, ainsi que les phytases (Mackey and Paytan 2009).
1.4.4. L’immobilisation
Lors du processus de l’immobilisation, le P labile est séquestré et retiré de l’environnement pour une période de temps.
Les procédés d’immobilisation peuvent être regroupés en deux catégories : La première catégorie, l’immobilisation transitoire ou l’assimilation cellulaire, comprend tous les processus de séquestration du P dans les cellules vivantes microbiennes et est rapidement réversible à la mort cellulaire. La deuxième catégorie appelée la formation de minéraux phosphatés, englobe les processus de minéralisation influencé par l’activité microbienne qui génèrent des minéraux contenant du P : il s’agit de la phosphogénèse (Mackey and Paytan 2009).
Les bactéries, champignons, micro-algues, protozoaires représentent la majorité de la biomasse vivante sur Terre. La plupart d’entre eux sont organisés en biofilms et sont très utiles à l’équilibre de notre planète.
1.5. Les biofilms associés à la mycorhizosphère
1.5.1. Les biofilms : définition, mécanisme de formation, structure et habitats
Les biofilms constituent l’ensemble des microorganismes (plus que 90 %) capables de s’attacher à certaines surfaces et former des communautés microbiennes (un mode de vie privilégié) plus ou moins complexes et symbiotiques. Ces microorganismes, qui adhérent entre eux et à une surface, sont marqués par la sécrétion d’une matrice adhésive, protectrice, extracellulaire et composée de polymères (Donlan and Costerton 2002).
Figure 1.3 : Les cinq étapes du développement d’un biofilm sur une surface dure.
Étape 1 : attachement initial; étape 2 : attachement irréversible; étape 3 : apparition et maturation I du biofilm; étape 4 : maturation II du biofilm; étape 5 : dispersion. Les photomicrographies, présentées toutes à la même échelle, sont celles d’un biofilm de
Pseudomonas aeruginosa en développement (Monroe D, 2007).
Dans la nature, chaque niche de biofilm est colonisée par un microbiote complexe adapté aux conditions de son microhabitat local (pH, température, oxygène et nutriments). Il s’agit en effet, d’un système ouvert et dynamique, au niveau duquel des échanges et des
migrations de populations affectent continuellement son organisation. D’autres acteurs extérieurs peuvent également venir ponctuellement sculpter cette architecture.
Généralement, les microorganismes forment des biofilms de la même manière, quelle que soit la nature de l’écosystème où ils vivent. Le cycle de développement d’un biofilm comporte cinq étapes qui peuvent se répéter indéfiniment. Ces étapes sont résumées dans la Figure 1.3.
La formation des biofilms est un perpétuel recommencement depuis l’adhésion des bactéries pionnières à une surface, la construction du biofilm, l’édification de sa structure tridimensionnelle (ou maturation) jusqu'à la dispersion et la libération de cellules spécialisées de la dissolution du biofilm. Les observations microscopiques effectuées sur des biofilms matures en utilisant la microscopie électronique à balayage montrent que ces derniers possèdent une structure hétérogène de cellules microbiennes organisées en monocouches à la surface.
Habituellement, on trouve les biofilms sur des substrats solides immergés ou exposés à une solution aqueuse. Ils se composent de nombreuses espèces de bactéries et d’archées vivants dans une matrice de polymères complexes. La matrice du biofilm contient des éléments synthétisés par des organismes qu’elle abrite (protéines, lipides, ADN, ARN, ...), elle est également constituée de polysaccharides et d’une importante proportion d’eau, elle assure la protection des cellules et facilite la communication entre elles par des signaux physicochimiques.
Dans un biofilm mature, les cellules produisent des messagers chimiques sous la forme de petites molécules médiatrices appelées aussi autoinducteurs. Ces molécules diffusibles activent l’expression de certains gènes codant pour une panoplie de fonctions essentielles, et ce en fonction de leur concentration elle-même reflétée par la densité de la population bactérienne du biofilm. Il s’agit en fait d’un mode de signalisation bactérien nommé le « Quorum Sensing » définit comme étant un processus très complexe faisant partie d’un réseau global de régulation de gènes intégrant divers signaux externes en vue de favoriser l’adaptation des bactéries aux conditions environnementales. Ceci implique que les bactéries formant le biofilm sont capables de percevoir les différents paramètres du milieu et exprimer les fonctions régulées adéquatement au type d’environnement rencontré.
1.5.2. Les biofilms associés aux plantes
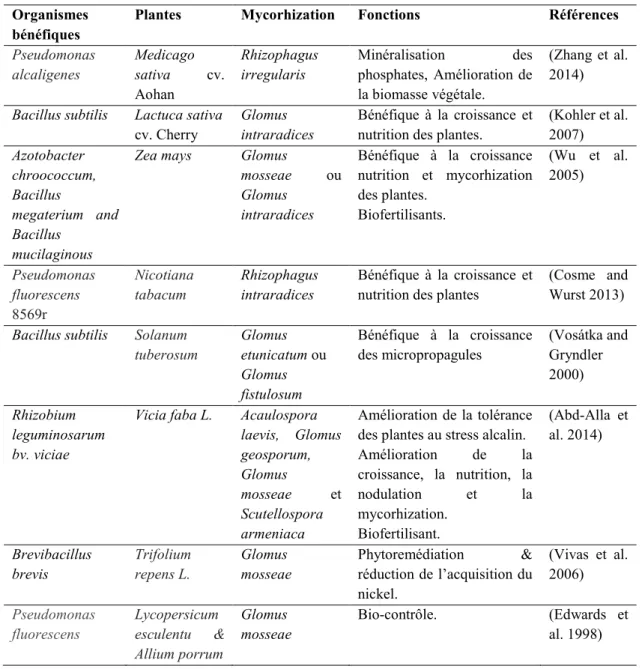
Le processus de la rhizodéposition (rejet de matières organiques des racines) et celui de la décomposition de la matière organique fraîche (agglomérat de molécules organisées d’une certaine manière) ont permis de développer les sols et de faire de la mycorhizosphère une niche écologique assez riche. La composition et la structure des constituants des sols représentent une véritable mine énergétique et structurante ce qui confère aux sols une meilleure disponibilité en nutriments facilement assimilables. Ces constituants représentent un stock de matières carbonées favorisant ainsi la stabilité de la structure du sol et le développement à long terme de l’activité microbiologique. Les microorganismes, dont la quasi-totalité sont hétérotrophes, ont développé des mécanismes pour en tirer profit (Davey and O'Toole 2000). Par exemple, certains microorganismes bénéfiques du sol vivent en symbiose étroite avec les racines des plantes, allant jusqu’à former des structures spécialisées et complexes comme les mycorhizes, les nodules et les biofilms. Ces modes de vie permettront à la fois d’améliorer la croissance microbienne et végétale. Dans un biofilm, les mucus bactériens, communément appelés matrice d’exopolysaccharides, permettent d’agglutiner diverses substances (fibres végétales, cristaux de roche et débris d’animaux) et de former par conséquent des microcavités assurant la protection des bactéries ainsi que le transport de l’eau et des nutriments. Dans la mycorhizosphère, les biofilms possèdent diverses particularités intéressantes en agriculture à savoir l’amélioration des échanges sol/plante, la synergie avec d’autres microorganismes du sol, la protection contre les pathogènes… (Tableau 1.1).
Tableau 1.1. Exemples de quelques bactéries bénéfiques dans la mycorhizosphère. Organismes
bénéfiques
Plantes Mycorhization Fonctions Références Pseudomonas alcaligenes Medicago sativa cv. Aohan Rhizophagus irregularis Minéralisation des phosphates, Amélioration de la biomasse végétale. (Zhang et al. 2014) Bacillus subtilis Lactuca sativa
cv. Cherry
Glomus intraradices
Bénéfique à la croissance et nutrition des plantes.
(Kohler et al. 2007) Azotobacter chroococcum, Bacillus megaterium and Bacillus mucilaginous
Zea mays Glomus
mosseae ou Glomus intraradices Bénéfique à la croissance nutrition et mycorhization des plantes. Biofertilisants. (Wu et al. 2005) Pseudomonas fluorescens 8569r Nicotiana tabacum Rhizophagus intraradices Bénéfique à la croissance et nutrition des plantes
(Cosme and Wurst 2013) Bacillus subtilis Solanum
tuberosum Glomus etunicatum ou Glomus fistulosum Bénéfique à la croissance des micropropagules (Vosátka and Gryndler 2000) Rhizobium leguminosarum bv. viciae
Vicia faba L. Acaulospora laevis, Glomus geosporum, Glomus mosseae et Scutellospora armeniaca Amélioration de la tolérance des plantes au stress alcalin. Amélioration de la croissance, la nutrition, la nodulation et la mycorhization. Biofertilisant. (Abd-Alla et al. 2014) Brevibacillus brevis Trifolium repens L. Glomus mosseae Phytoremédiation & réduction de l’acquisition du nickel. (Vivas et al. 2006) Pseudomonas fluorescens Lycopersicum esculentu & Allium porrum Glomus mosseae Bio-contrôle. (Edwards et al. 1998)
1.5.2.1. Implication des mycorhizes dans la croissance des bactéries
Diverses études ont rapporté l’influence des populations bactériennes présentes dans le sol sur la colonisation du système racinaire des plantes par les champignons mycorhiziens. En revanche, l’influence directe du mycélium extraracinaire des champignons mycorhiziens sur la croissance bactérienne et la composition des communautés n’est toujours pas très bien comprise.
De nombreux mécanismes de l’interaction mycélium-bactérie ont été discutés, à savoir la translocation du carbone dans les tissus fongiques (Andrade et al. 1997; Bago et al. 2002; Trépanier 2005), le changement de la structure du sol (Tisdall and Oades 1979), la compétition pour les nutriments disponibles (Ravnskov et al. 1999) et les changements physiologiques des exsudats racinaires (Söderberg et al. 2002). Mais, certaines informations concernant la composition des exsudats fongiques des MA, et leur disponibilité dans le sol pour le profit des communautés microbiennes n'ont pas été très bien étudiées (Bending et al. 2006).
Filion et ses collaborateurs (1999) ont étudié l'effet des exsudats fongiques sur la croissance de quelques souches bactériennes présentes dans le sol. Ils ont constaté que ces extraits pourraient parfois avoir un effet antagoniste, mais dans la plupart des cas des effets stimulants de la croissance des microorganismes, ce qui suggère que les produits exsudés par le mycélium peuvent jouer un rôle important dans les interactions directes entre les CMA et les autres microorganismes du sol.
D’autres chercheurs ont testé la capacité d'un certain nombre de bactéries du sol à coloniser les hyphes des CMA mortes ou vivantes (Toljander et al. 2006). Ils ont montré que certaines souches ont tendance à s’attacher aux hyphes vivantes tandis que d’autres préfèrent les hyphes mortes. Ils ont conclu que certaines bactéries ont une stratégie biotrophique, elles utilisent les exsudats libérés par les hyphes fongiques vivantes, alors que d’autres espèces bactériennes utilisent les hyphes eux-mêmes comme substrat (Toljander et al. 2006). Toljander et ses collaborateurs (2006) ont fini par conclure que les MA peuvent avoir une influence sur la composition des communautés bactériennes associées à la mycorhizosphère.
Des études in vitro ont permis aux chercheurs de démontrer que les exsudats fongiques MA extraits à partir d’un milieu de culture de racines peuvent avoir une importance quantitative et une influence qualitative sur les communautés bactériennes. Si le traitement des bactéries par les exsudats fongiques in vitro a permis d’améliorer la diversité des communautés bactériennes sans qu’il n’y ait un contact physique ‘CMA-bactérie’, on pourrait s’attendre à une rétroaction positive résultante de l’interaction ‘mycélium-bactérie’ dans les sols, puisque la quantité d’exsudats mycéliens dans le sol peut être stimulée par les bactéries se trouvant sur les hyphes ce qui permet d’avoir une exsudation accrue (Toljander et al. 2007).
Il a été montré que les exsudats du Glomus sp. MUCL 43205 contiennent des sucres de faibles masses moléculaires et des acides organiques, qui sont probablement métabolisés par les bactéries. D’autres composés de hautes masses moléculaires sont aussi présents, mais n’ont pas été identifiés, ces composés sont certainement responsables de la croissance des bactéries du sol (Toljander et al. 2007). D’autres recherches ont été menées sur la colonisation de Glomus sp. MUCL 43205 par Pseudomonas fluorescens, et ont montré que certaines souches de Pseudomonas sont capables de former des biofilms sur les racines (Lugtenberg et al. 2001). L'analyse du biofilm formé a montré qu'il se compose de bactéries couvertes d’un matériel mucilagineux permettant l’hydratation des surfaces pour l’ensemble de l’agrégat bactérien. C’est ce matériel mucilagineux qui facilite la colonisation des racines et des hyphes (Bianciotto et al. 1996; Bianciotto et al. 2001; Matthysse and McMahan 1998).
Certaines bactéries du genre Paenibacillus peuvent être intimement associées au mycélium de Glomus intraradices (Mansfeld-Giese et al. 2002). Artursson et Jansson (2003) ont constaté que Bacillus cereus isolée à partir du sol possède des niveaux plus élevés d’attachement aux hyphes de Glomus dussii en comparaison avec d'autres souches bactériennes.
Scheublin et ses collaborateurs (2010) ont prouvé que la colonisation de l’hyphosphère de deux CMA (Glomus intraradices et Glomus proliferum) est spécifique à certains groupes de bactéries. L’analyse des communautés bactériennes attachées ou non au mycélium fongique a montré l’abondance d’un groupe de bactéries de la famille des
Oxalobacteraceae intimement attaché aux hyphes, suggérant ainsi la présence d’un
dialogue spécifique entre les microorganismes (Scheublin et al. 2010).
En effet, il semble que certaines bactéries sont plus spécifiques à un type particulier de MA, qui pourrait être due à la sécrétion d'exsudats spécifiques par l’espèce fongique (Artursson and Jansson 2003). Bacillus pabuli a la capacité d’améliorer la colonisation des racines par les mycorhizes arbusculaires (Xavier and Germida 2003) et peut également améliorer la croissance des plantes (Artursson et al. 2006).
La double inoculation CMA et bactéries permet d’améliorer l'utilisation du phosphate de roche par les plantes. Ce phénomène pourrait être lié à la solubilisation du phosphate de roche par les bactéries solubilisant les phosphates et la libération du P dont une partie sera
prise par le CMA et assurera son développement et l’autre sera transportée à la plante par l’intermédiaire du réseau mycélien.
En résumé, il y aura un échange direct et réciproque mycélium-bactérie au niveau de la mycorhizosphère permettant à la plante une meilleure croissance.
Les résultats d’une étude menée par Toro et ses collaborateurs en 1997 visant à comprendre l'interaction entre une pratique biotechnologique (l'inoculation microbienne) et une technologie à faibles intrants (l'application du phosphate de roche) ont démontré l'efficacité de ces pratiques combinées à l'amélioration durable de l'apport d'éléments nutritifs pour les plantes. Cette efficacité repose sur l'amélioration des performances des microorganismes des sols (Toro et al. 1997).
Récemment des recherches en écologie microbienne ont révélé qu'un traitement, au niveau du compartiment distal, avec des spores de Glomus irregulare isolées à partir d'une rhizosphère d'Agrotis stolanifera sur des cultures in vitro de racines mycorhizées par la même espèce fongique ont permis d'observer, par microscopie à contraste interférentiel, une croissance bactérienne clairement visible autour des hyphes. Ces bactéries semblent être associées au réseau mycélien (Lecomte et al. 2011).
Par ailleurs, plusieurs études ont mis en évidence le rôle éventuel du tréhalose dans les interactions entre les bactéries et les champignons mycorhiziens. Il est notoire que lorsqu'il est associé avec les racines des plantes, le champignon mycorhizien recevra jusqu'à 30% du carbone total et le transforme en tréhalose (Wiemken 2007). Le tréhalose a été montré comme pouvant être responsable de la sélection de certaines communautés bactériennes dans la mycorhizosphère des racines des arbres dans les pépinières forestières et les plantations (Izumi et al. 2006; Uroz et al. 2007). En outre, il a été démontré in vitro, dans le cas des champignons ectomycorhiziens, que le tréhalose est impliqué dans la promotion de la croissance de Pseudomonas monteilii sur les hyphes de Pisolithus albus (Duponnois and Kisa 2006). D’autres chercheurs ont récemment observé que la souche de Pseudomonas
fluorescens BBc6R8 est capable de former un biofilm sur le mycélium de Laccaria bicolor
S238N lieu d’accumulation du tréhalose (Deveau et al. 2007).
Par conséquent, nous pouvons maintenant admettre l'hypothèse que les métabolites fongiques tels que le tréhalose peuvent faciliter la colonisation des hyphes par les microorganismes eux-mêmes capables de former des biofilms (Frey-Klett et al. 2007).
Toutes ces études fournissent de nouvelles preuves de la colonisation des hyphes de différentes espèces de champignons mycorhiziens par les bactéries, mais certaines données restent manquantes surtout concernant les CMA.
1.5.2.2. Avantages écologiques des biofilms
Il y a eu des spéculations sur les avantages de la formation des biofilms par rapport au mode de vie des cellules à l’état planctonique. Bien qu’il soit difficile de tester expérimentalement ces spéculations, des études ont été menées pour connaitre les raisons pour lesquelles la stratégie de formation des biofilms a été adoptée par autant de microbes. Parmi les fonctions connues des biofilms, on peut citer l’exemple de :
- La coopérativité métabolique et disponibilité des éléments nutritifs :
En effet, les canaux dispersés à travers les biofilms dans les zones entourant les microcolonies ont été comparés à un système primitif circulatoire (Davey and O'Toole 2000). Ces canaux constituent un moyen efficace d’échange de nutriments et de métabolites avec la phase aqueuse, d’amélioration de la disponibilité des nutriments ainsi que la suppression de métabolites potentiellement toxiques (Costerton et al. 1995). Les caractéristiques métaboliques des bactéries au sein d’une communauté de biofilms sont distinctes de ceux de leurs homologues planctoniques. Il est à noter que l’architecture complexe des biofilms prévoit la possibilité d’une coopération métabolique entre les différentes bactéries, qui sont exposées à un éventail de différents signaux environnementaux. Ainsi les biofilms constituent un environnement idéal pour l’établissement de relations syntrophiques (Davey and O'Toole 2000).
Les biofilms peuvent aussi augmenter l’activité biofertilisante des organismes qui en font partie assurant ainsi un effet positif sur la croissance et le rendement des cultures (Seneviratne et al. 2009). Ces biofilms peuvent contenir une seule espèce microbienne ou plusieurs. Seneviratne et al. (2009) ont décrit les études menées dans ce domaine essentiellement dans le développement de biofilms de bactéries fixatrices d’azote et de champignons solubilisant les phosphates (Seneviratne et al. 2009). Une co-culture in vitro de ces deux microorganismes a révélé que les bactéries colonisent le mycélium pour former