L'évaluation de l'ATP comme mesure du carbone vivant (biomasse) phytoplanctonique dans un écosystème aquatique.
Texte intégral
Figure



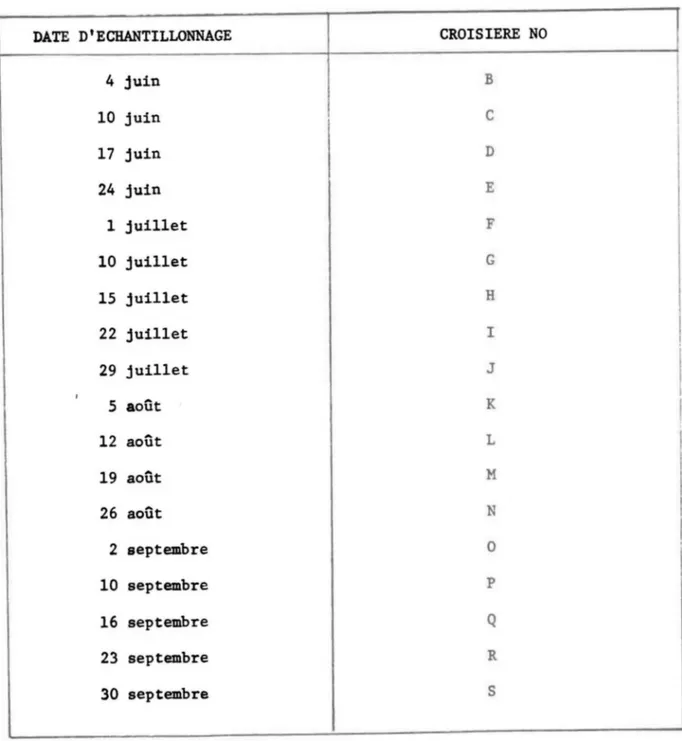
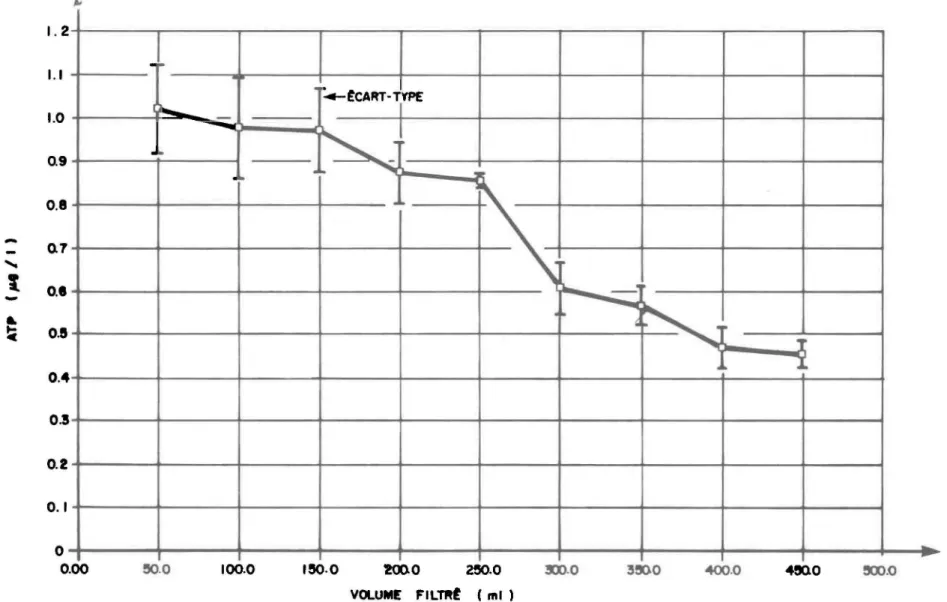
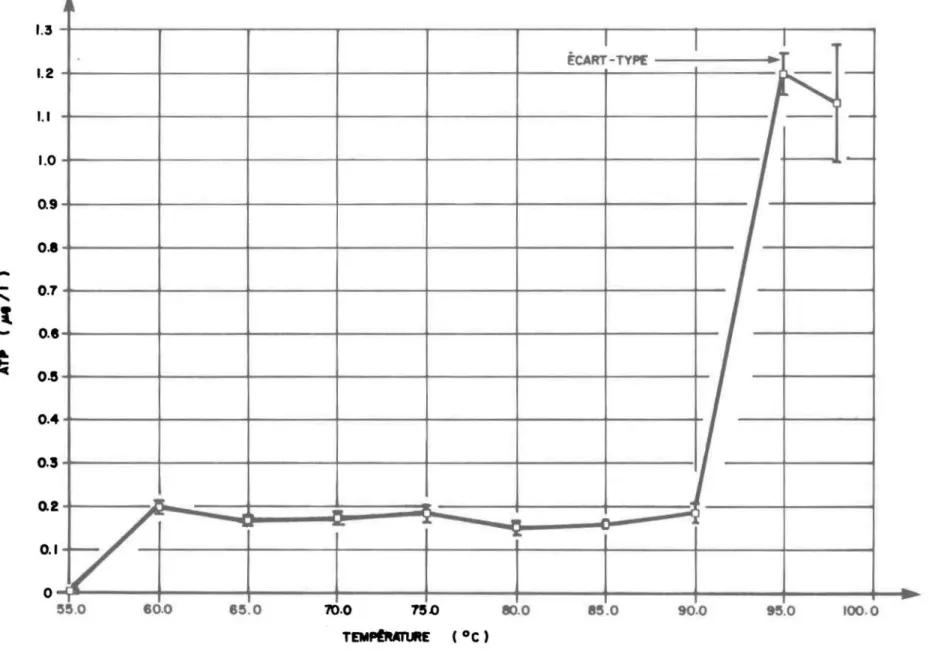
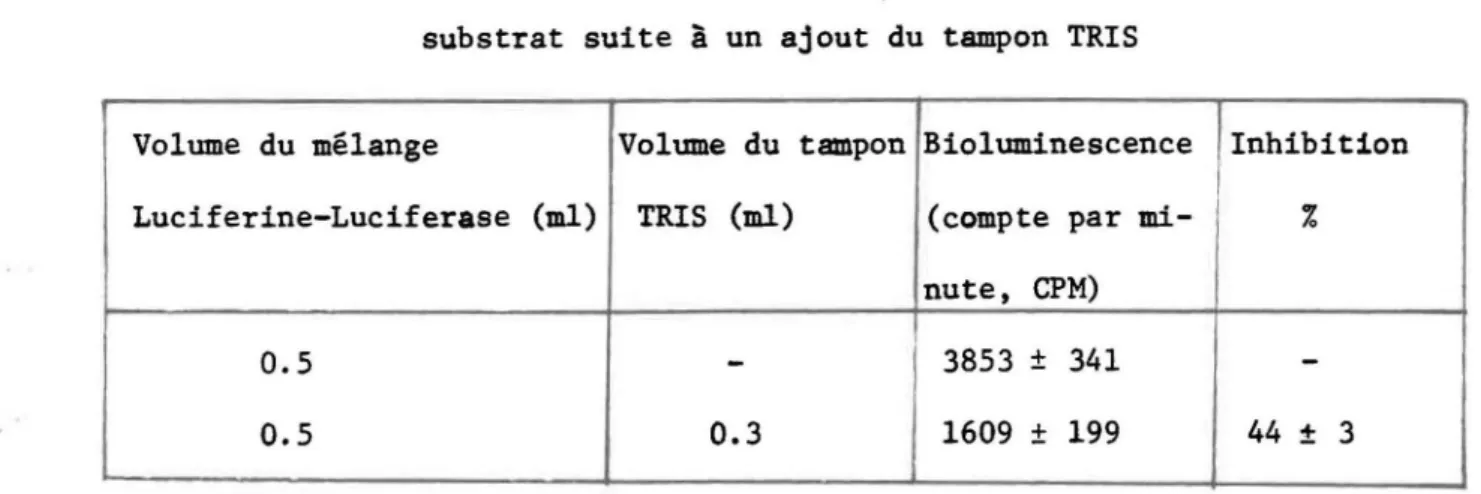
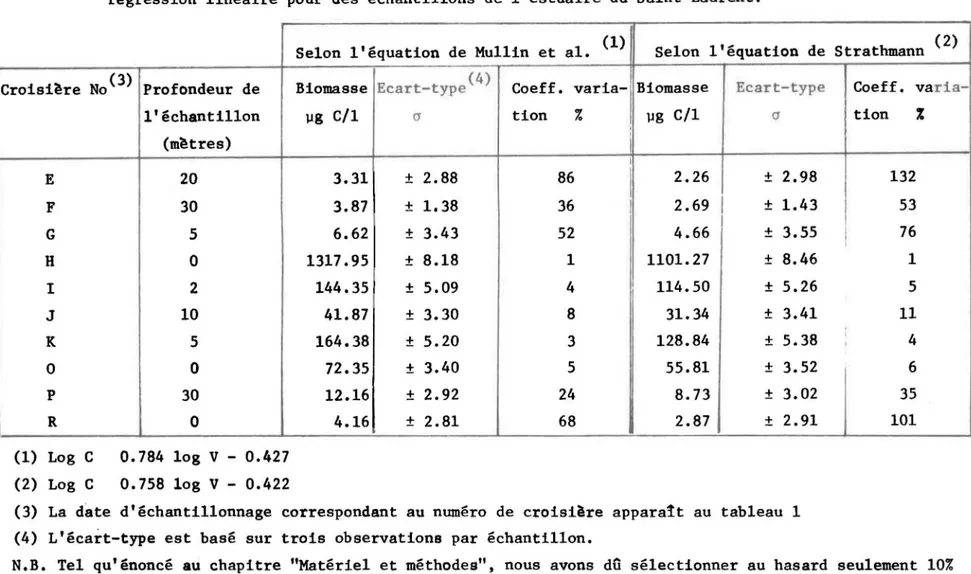
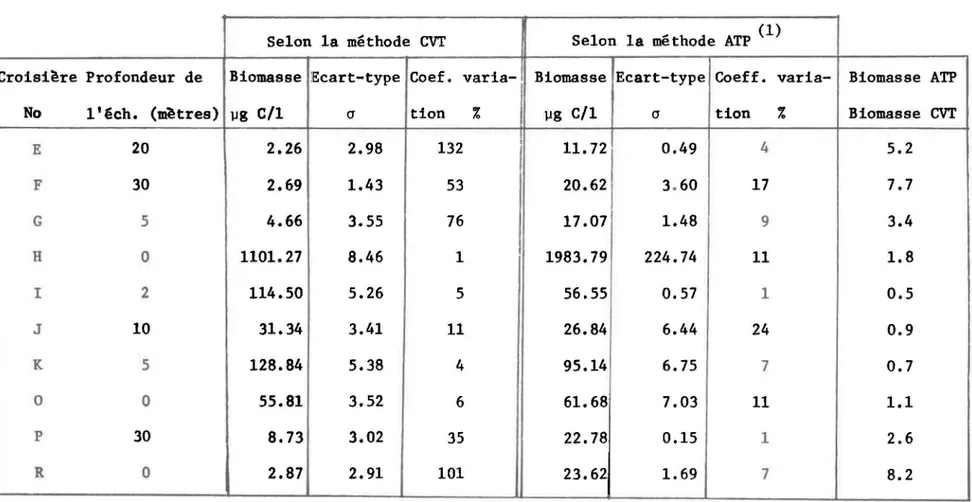
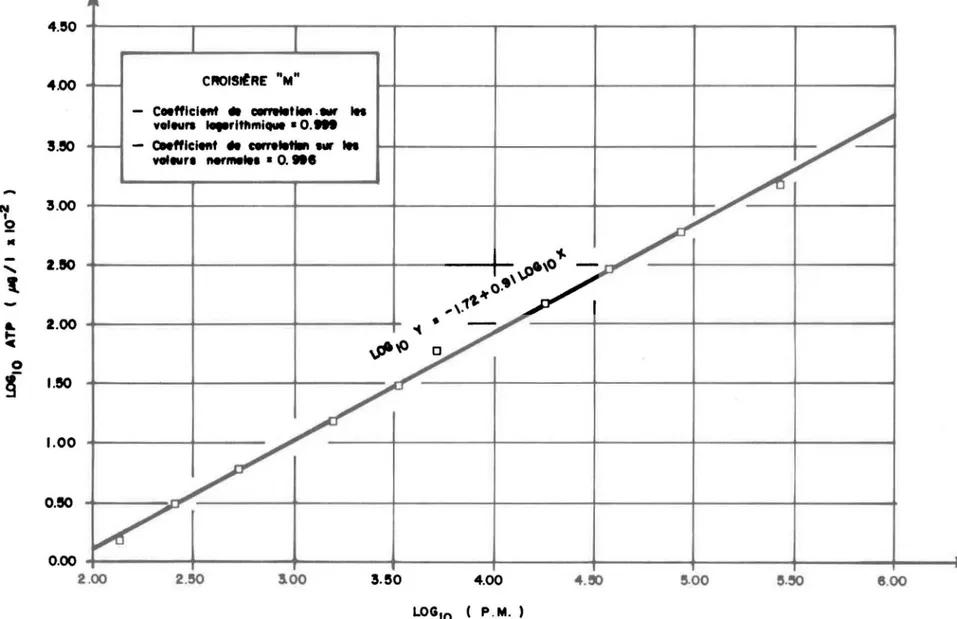
Documents relatifs
De profondeur moins 6 m, constitué d’un réservoir naturel d’eau douce, pour déterminer la qualité bactériologique, la qualité phytoplanctonique et quelque
- Certaines espèces du phytoplancton, peuvent être utilisées comme des indicateurs de pollution, ainsi Chamaesiphon polonius et Calothris sp sont caractéristiques des
Il ne s’agit pas d’un mod` ele biologique donc il n’y a pas de compartiment de nitrate mod´ elis´ e (cette remarque est ´ egalement valable pour les flotteurs lagrangiens qui
probability to be crossed by a source during a microlensing survey. In reality, it is not easy to detect a secondary caustic deviation, because it is impossible to predict when it
حايزنلاا ةيرعش ثيدحلا يبرعلا رعشلا يف : برتعي حايزنلاا موجه ا ىلع رقتسلما دكارلا و قرخ ل ،ةداعل وه طاب تسا يحلل يما يدلا وإ طاقس ،دف تسملل
442quater, § 4 du CIR 92: La responsabilité solidaire des dirigeants de la société ou de la personne morale ne peut être engagée que pour le paiement, en principal et intérêts,
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des
borée par Fieras et Elliott (1992), qui font une excellente revue de la conception de l’éducation multiculturelle dans le contexte canadien,.. Ces auteures distinguent
