Regulation of mitotic BubR1 phosphorylation by the BubR1 pseudokinase domain
Texte intégral
Figure


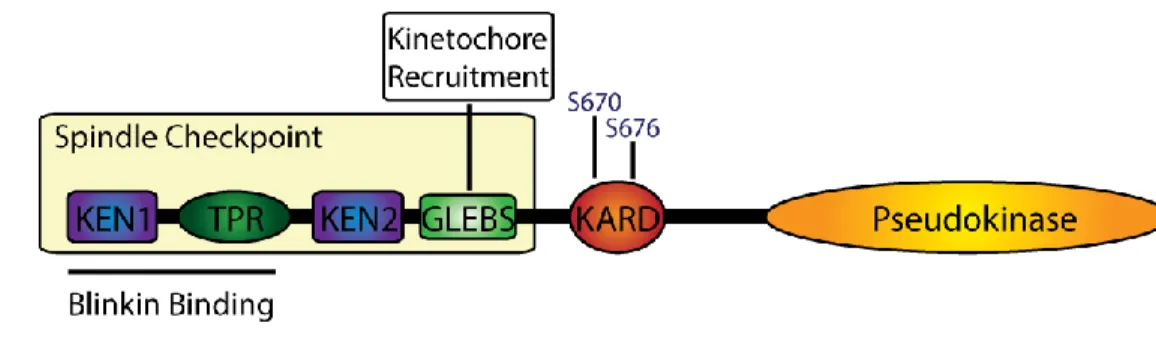
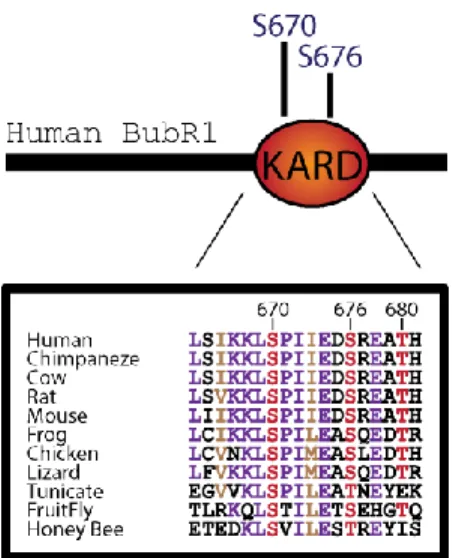
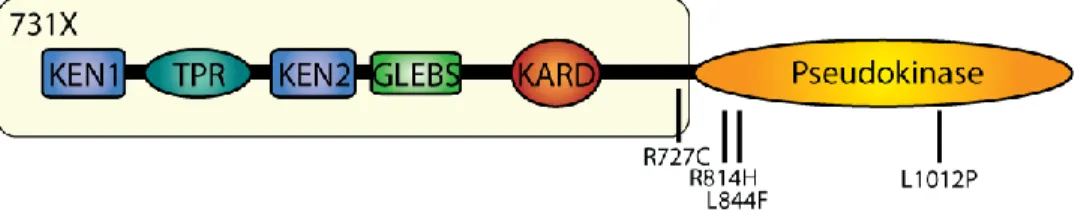
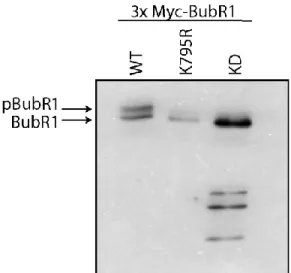
Documents relatifs
In the end, we observed that actin binding to Ezrin was maximum two hours after nocodazole release (Figure 5D), likely a consequence of elevated T567 phosphorylation
METHODS: We tested for the presence of copy number variations in the pseudoautosomal region of the sex chromosomes in 735 individuals with idiopathic short stature and compared
The agents that cause infant diar- rhoea have been largely identified, and vaccine candidates against most of the important pathogens (rotavirus, enterotoxigenic E.
We use func- tional assays to describe the effects of multiple different CHD3 mutations on ATPase activity and chromatin remodeling capa- cities.. Taken together, our data
Dans un objectif de pilotage agro-écologique des agrosystèmes, et plus largement d’évaluation environnementale de l’effet de l’action de l’Homme sur les écosystèmes agricoles,
Amounts of fumonisins (FB 1 and FB 2 ) and beauvericin (BEA) produced in the healthy, adjacent, and inoculated fruitlets of pineapple fruits artificially infected with
Nevertheless, the mitotic phenotypes are similar between neuroblasts and HeLa cells: the centrosomal microtubule asters are present, but they only nucleate short
Using siRNA knockdown of hGwl we show in human cells that this kinase mediates mitotic entry and maintains the mitotic state by inhibiting PP2A and thus, keeping the
