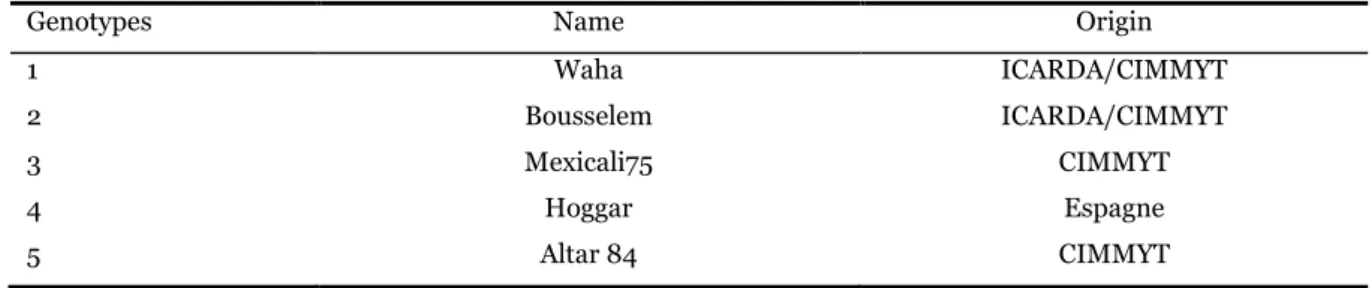
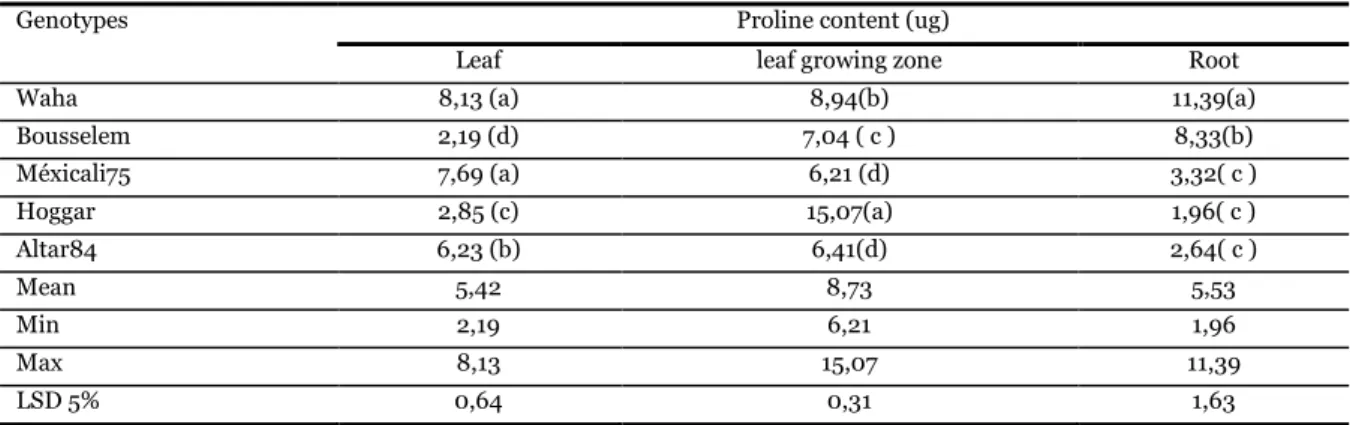
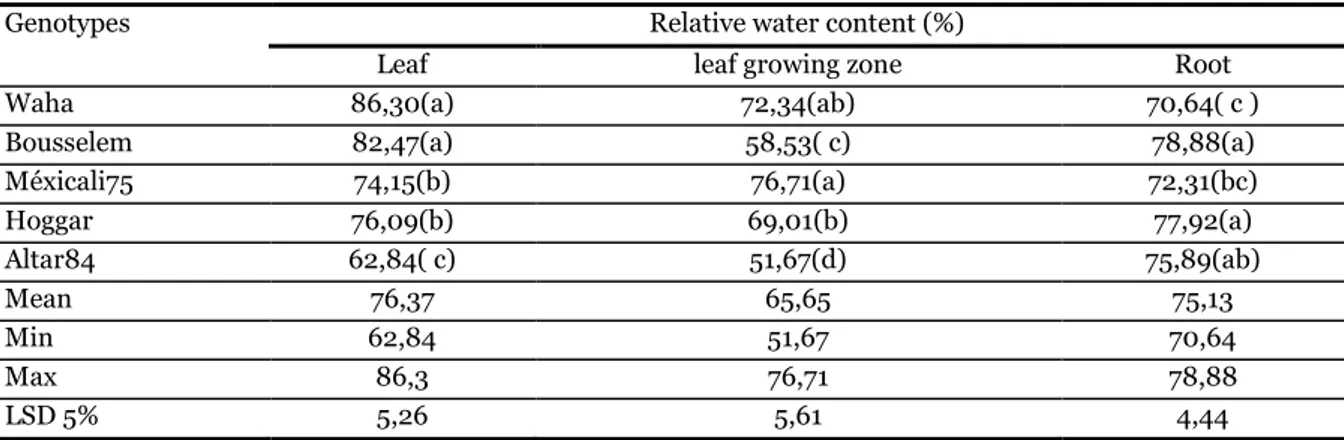
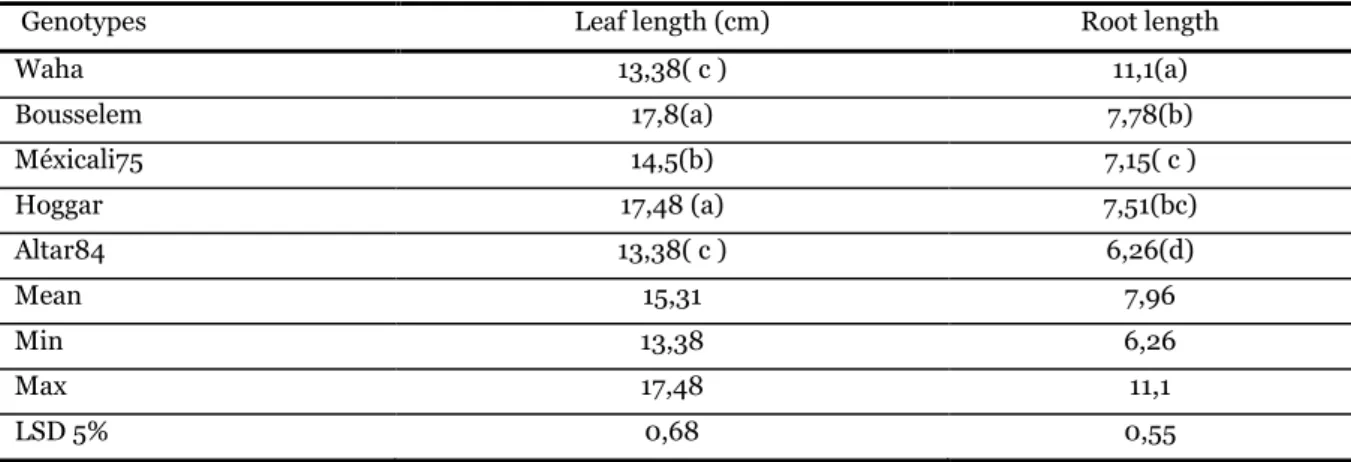
The effect of water stress on some physiological and biochemical traits in five durum wheat (Triticum durum Desf.) genotypes
Texte intégral
Figure
Documents relatifs
Silicon application in the nutrient solution provides some evi- dence that Si mitigates the effect of water stress by improving shoot and root development, and water uptake
Effet d’un stress osmotique sur l’accumulation de proline, de chlorophylle et des ARNm codant pour la glutamine synthétase chez trois variétés de blé dur (Triticum durum
As the durum wheat cultivars’ wet gluten content, gluten extensibility, semolina yield, the semolina’s yellow pigment content, highly depends on the growing conditions (area,
The purpose of this experiment was to determine whether genotypic differences in photosynthetically water use efficiency (WUEg) and related traits exist in 12 durum wheat
The aim of our work was: (i) to analyse the relationship between whole plant and cellular response to salt stress in durum wheat; and (ii) to determine the relative importance of
dactylon on seed germination of the three varieties of durum wheat: MBB, GTADUR and VITRON under controlled conditions (laboratory) revealed that there is an inhibition of
Altogether, our study confirm that GR and OR are 2 durum wheat varie- ties contrasted with respect to salt stress tolerance and define TdERF1 as puta- tive marker in selection
Variation in growth, canopy temperature, translocation and yield of four durum wheat (Triticum durum Desf.) genotypes under semi arid condition.. Genetic variability, correlation
