G-quadruplex : acteurs majeurs de la duplication du génome humain
Texte intégral
Figure
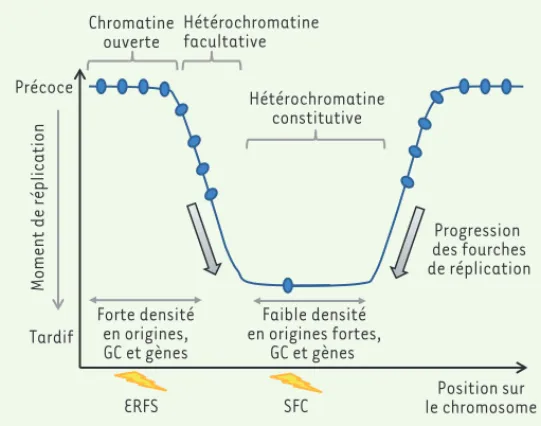

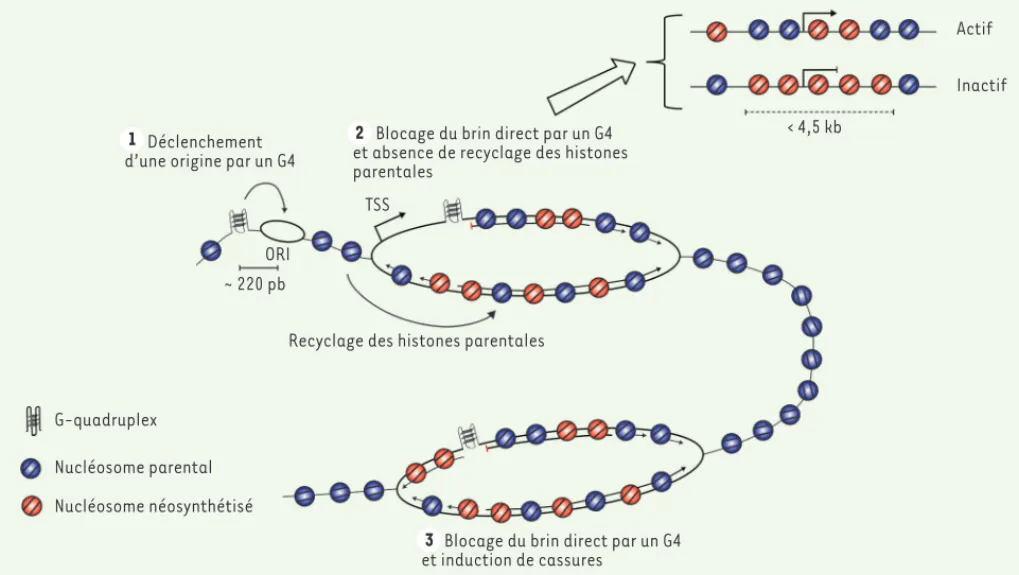
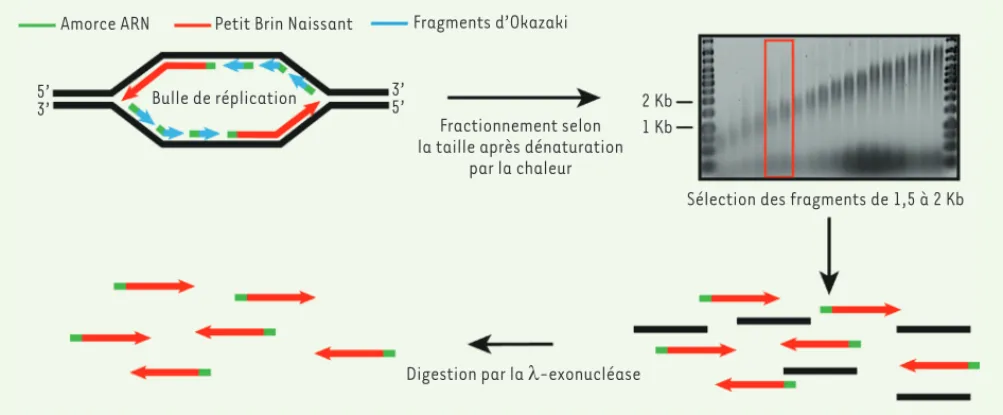
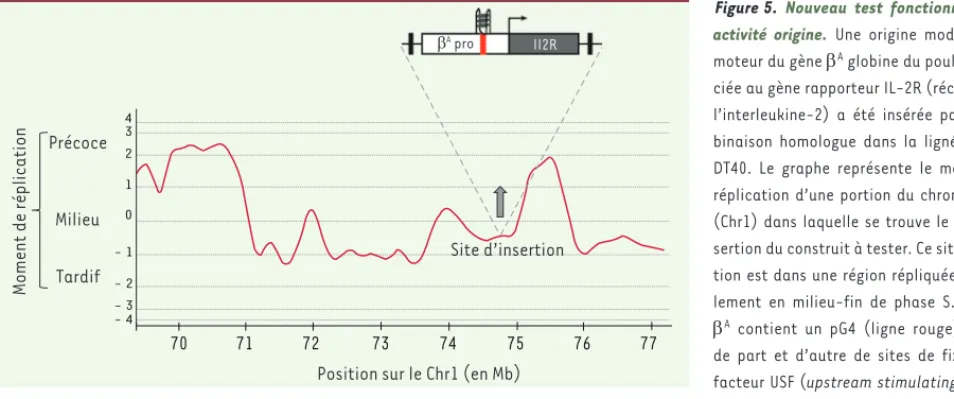
Documents relatifs
Après la phase S, la molécule d’ADN a été copié fidèlement et on obtient 2 molécules d’ADN jumelles : 2 chromatides réunies par un centromère = chromosome double qui va
En 1953 les travaux de Franklin, Watson, Crick et Wilkins sont publiés et élucident la structure de l’ADN, support de l’information génétique.. Mais on ignore toujours le
Accumulation d’ADN simple brin dans les cellules déplétées pour SAMHD1 co-transfectées avec un siRNA contrôle (si-Ctrl) ou un siRNA dirigé contre RecQ1 (si-RecQ1) ou traitées avec
reliés par des liaisons phosphodiester entre le carbone 3 ’ d ’un nucléotide et le carbone 5 ’ du suivant... Les acides nucléiques:
Il s'agit d'une hélicase, protéine qui sépare le double brin d'ADN. DnaC : Protéine codée par le
La réplication de l’ADN chez les eucaryotes est tout à fait comparable à la réplication de l’ADN chez les procaryotes.. Elle est généralement bidirectionnelle, elle est
Les grandes étapes de la biologie moléculaire Expérience historique de Meselson et Stahl
Topoisomérase de type II coupe les deux brins simultanément (Formes relaxée/hyperenroulée de l’ADN
