Étude exploratoire de l'influence modulatrice du système nerveux autonome sur l'interprétation de la douleur
Texte intégral
Figure







Documents relatifs
Deux objectifs principaux sont visés par cette étude. Premièrement, nous visons à décrire le système de reproduction du saumon atlantique pour l'ensemble d'une population sauvage
Enfin, les résultats obtenus mettent en évidence le potentiel détoxifiant du système enzymatique de glutathion exprimé par la diminution de la concentration de glutathion
cycles de production entre le système semi intensif et le système extensif viendrait de ce que, dans le système semi intensif, les poules qui bénéficient des conditions de
Les résultats montrent une relation de causalité négative entre certains indicateurs de solidité du système bancaire marocain (le ratio de solvabilité et le taux
Ce processus de régulation se manifeste par des stratégies ou des modalités de régulation qui, dans le cadre de cette étude, ont été définies comme des
Le but de cet étude est de caractériser la régulation de 1' expression des différents récepteurs de LDL dans le placenta en fonction du profil lipidique maternel,
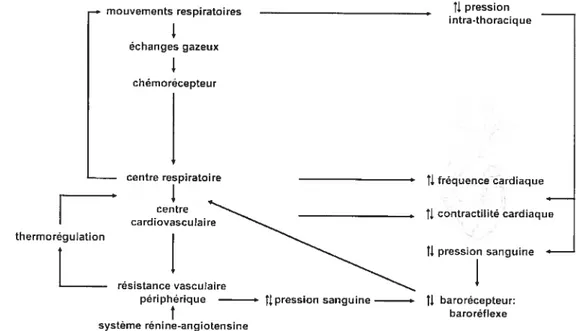
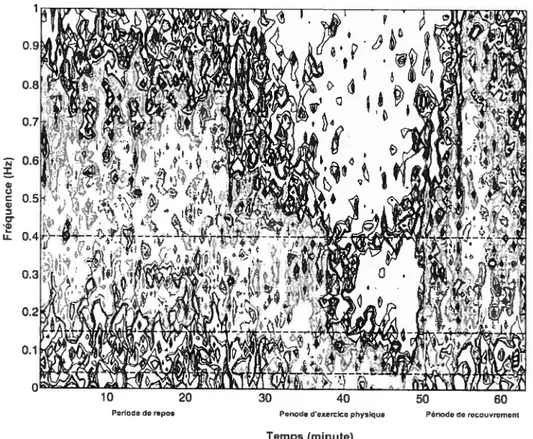
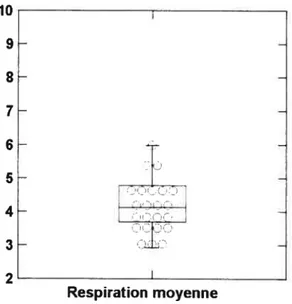
Les r´ esultats pr´ eliminaires montrent une bonne s´ eparation, dans le domaine fr´ equentiel, des tendances SNAS et SNAP, incluant un recouvrement fr´ equentiel dans les
Pour une meilleur connaissance de la flore algérienne et dans le but d’évaluer la variabilité interspécifique et l’organisation des génomes, une étude cytogénétique
