ANTOINE RICHARD
DESCRIPTION DU SYSTEME DE REPRODUCTION
DU SAUMON ATLANTIQUE (SALMO SALAR L.) ET
IMPACT DE LA REMISE À L'EAU : UNE ÉTUDE
MOLÉCULAIRE
Mémoire présenté
à la Faculté des études supérieures et postdoctorales de l'Université Laval dans le cadre du programme de maîtrise en Biologie
pour l'obtention du grade de Maître es sciences (M.Sc.)
DEPARTEMENT DE BIOLOGIE FACULTÉ DES SCIENCES ET DE GÉNIE
UNIVERSITÉ LAVAL QUÉBEC
2012
Résumé
Dans cette étude, nous tirons avantage de l'outil moléculaire et des techniques d'assignation parentale afin de décrire le système de reproduction du saumon atlantique (Salmo salar L.) et de mesurer l'impact de la remise à l'eau sur son succès reproducteur (SR). La variance du SR observée au sein des mâles était plus importante qu'au sein des femelles. Mâles et femelles sont majoritairement polygames et le nombre de partenaires est d'ailleurs positivement corrélé au SR. Les tacons précoces ont produit 44% des alevins réassignés. Les saumons remis à l'eau participent à la reproduction, mais la taille du saumon ainsi que l'interaction entre la température de l'eau et l'exposition à l'air ont un impact négatif sur le SR d'un saumon remis à l'eau. Finalement, nos résultats améliorent notre compréhension du système de reproduction du saumon atlantique et confirment que les saumons remis à l'eau se reproduisent et participent à la pérennité des populations exploitées.
11
Abstract
In this study, we took advantage of the molecular tool to document in details the breeding system of a wild population of Atlantic salmon (Salmo salar L.) and to assess the impacts of catch and release (C&R) on its reproductive success. Both sexes showed high variance in individual reproductive success and estimated standardized variance was higher for males than for females. Males and females were polygamous and their fitness was strongly and positively correlated with the number of mates. Mature male parr sired 44% of the analyzed offspring. Catch and released salmon participated in the reproduction, but the size of the fish and the interactive effect of water temperature and air exposure have a negative impact on reproductive success. This study contributes to improve our understanding on the species' complex reproductive biology and confirm that C&R represents an efficient management tool to promote conservation of salmon populations with limited negative impact on mating and reproductive success.
Avant-Propos
Ce mémoire est constitué de trois chapitres. Le corps de ce mémoire, le chapitre 2, est rédigé en anglais sous forme d'article scientifique en vue d'une future publication. Le premier et le dernier chapitre, soit l'introduction et la conclusion générale, sont rédigés en français. Mon directeur Louis Bernatchez ainsi que ma co-directrice Melanie Dionne sont co-auteurs de l'article formant le chapitre 2 puisqu'ils ont participé à l'élaboration de l'étude et qu'ils ont grandement contribué, par leurs judicieux commentaires et suggestions, à améliorer la qualité de ce manuscrit. Le Dr Jinliang Wang, de l'institut de zoologie de Londres, a développé les outils statistiques et logiciels nécessaires à l'analyse des données d'assignation parentale et est également co-auteur du chapitre 2.
Remerciements
Tout d'abord, je tiens à remercier mon directeur Louis Bernatchez ainsi que ma co-directrice Melanie Dionne. Louis, je veux premièrement te remercier de m'avoir fait découvrir, lors de mes études au baccalauréat, les innombrables opportunités qu'offre l'approche moléculaire aux écologistes. Sans ta passion contagieuse, ma phobie des micropipettes et du sarreau m'aurait empêché de découvrir les joies de la biologie moléculaire et je n'aurais jamais entrepris cette maîtrise qui m'a tant passionné. Merci aussi de ta confiance et de l'intérêt que tu as porté envers ce projet. Melanie, je veux te remercier pour ton enthousiasme et pour ton positivisme qui m'ont fait foncer quand j'aurais pu reculer. Merci aussi pour toute la confiance que tu m'as accordée tout au long de ces années.
Je pense que je peux dire sans exagérer que ce projet de recherche était très ambitieux et sa réussite découle de la collaboration d'un nombre impressionnant de personnes. À vous tous, merci! Ce fut une grande joie et un honneur de travailler et d'apprendre à vos côtés. Plus spécifiquement, merci à mes muses d'été; Éliane, Jade et Catherine, vos efforts, vos judicieux conseils ainsi que vos sourires ont non seulement permis de mener à bien les campagnes d'échantillonnages, mais ils les ont rendu fort agréables! Merci également à Marc, Daniel, Denis, Nicolas, Vanessa et William du MRNF pour vos compétences, votre efficacité et votre disponibilité.
IV
Je tiens à remercier tous les gens de la ZEC de la rivière des Escoumins et de l'organisme de bassin versant de la haute Côte-Nord. Amélie, Sylvain, Yves, Maurice, Mike, Gislaine, Marie-Hélène et Chantai : votre accueil et votre collaboration ont été extraordinaires!
Un grand merci aux pêcheurs de la rivière des Escoumins. Sans vous ce projet était tout simplement impossible et votre participation a été exceptionnelle. La passion et l'amour que vous portez au saumon atlantique sont les meilleurs garants de l'avenir du roi de nos rivières.
Merci aussi à toute la gang du laboratoire Bernatchez pour les bonnes discussions. Un merci spécial à Lucie et à Guillaume. Grâce à vous, l'irréductible écologiste que j'étais est aujourd'hui devenu généticien. Merci aussi, Scott, pour ton aide de correction de mon anglais aux forts accents québécois.
Je tiens à remercier mes amis qui m'ont soutenu lorsque les difficultés me faisaient croire que je ne verrais jamais la fin de ce travail et qui ont surtout su rendre cette période de ma vie purement mémorable.
En terminant merci aussi à ma famille. Louise et Pierre, vous avez su dès mon plus jeune âge me transmettre votre amour pour la nature. Encore poupon, vous m'apportiez avec vous en canot sur la Ristigouche et cela n'est sûrement pas étranger à l'écriture de ce mémoire!
«[...] les saumons nous donnoient bien plus de peine, il en entre en cette rivière une si grande quantité, que la nuit l'on ne peut dormir tant est grand le bruit qu'ils font en tombant sur l'eau après s'estre jettez ou élancez en l'air [...] » (Denys 1672)
« Amateur fishermen would bask in the sunshine of their desires, always sure of sport. [...] Mr. Strang, of Quebec, carried off the palm from the Escoumins, having killed his fifty fish in two days' fishing. » (Nettle 1857)
Table des matières
Résumé i Abstract ii Avant-Propos iii Table des matières vi Liste des tableaux viii Liste des figures ix I. Introduction 1
Prologue 1 Sélection sexuelle et tactiques de reproduction alternatives 1
Le saumon atlantique (Salmo salar L.) 2
Distribution géographique 2
Cycle de vie 2 Système de reproduction du saumon atlantique 3
Pourquoi la capture de saumon atlantique doit-elle être réglementée ? 4
Statut actuel du saumon atlantique 4 La pêche commerciale d'hier à aujourd'hui 5
Impact de la pêche sportive sur les populations de saumon atlantique 6 Pourquoi la remise à l'eau est-elle un outil de gestion et de conservation efficace ? 7
La remise à l'eau comme outil de gestion et de conservation 7
Efficacité de la remise à l'eau 7 Est-ce que la remise à l'eau affecte la valeur sélective des saumons graciés? 8
Valeur sélective : définition 8 Taux de survie suivant un événement de remise à l'eau 9
Effet de la remise à l'eau sur la migration en rivière 9 Effet de la remise à l'eau sur la reproduction 12 Assignation parentale grâce aux marqueurs moléculaires polymorphes 14
Objectifs 14 IL Does catch and release affect the mating system and individual reproductive success of
wild Atlantic salmon {Salmo salar L.)? 15
Résumé 16 Abstract 17 Introduction 18 Materials and methods 21
Study site 21 Sample collection 22
Genetic analysis 24 Parentage analysis 24 Reproductive success analysis 26
Results 29 Description of the mating system 31
Determinants of the reproductive success 33
Discussion 41 Female mating pattern 42
M l
Mature males parr 44 Impact of catch and release 45
Acknowledgments 48 III. Conclusion générale 50
Bibliographie 54 Annexe 1 62
Liste des tableaux
Table 1. Parameter estimates for each of the models in the confidence set of logistic models for grilse access to reproduction. When a variable is not part of a model, the corresponding cell is left blank. Models are ranked by AICC. For each model, the number of parameter (k),
the AAICC and the Akaike weight are shown. The model-averaged estimate, and its
associated 95% unconditional CI, of each parameter as well as it's importance weight are also shown. In bold is the only parameter (Date of River Entrance) with a significant effect
on the grilse capacity to produce at least one offspring 35 Table 2. Parameters of the 10 best negative binomial models for the number of produced
offspring by large salmon. For each model, parameter estimates are given for included quantitative variables whereas inclusion of a qualitative variable is represented by a "+". When a variable is not part of a model, the corresponding cell is left blank. Models are ranked by AICC. For each model, the number of parameter (k), the AAICC and the Akaike
weight (co) are shown. The model-averaged estimate, and its associated 95% unconditional CI, of each parameter, as well as its importance weight are also shown. In bold are the parameters with a significant influence on the number of produced offspring for large
Liste des figures
Figure 1. The Escoumins River, and its tributaries, on the north shore of the St-Lawrence River in Québec showing the positions of the obstacles to salmon migration, as well as the
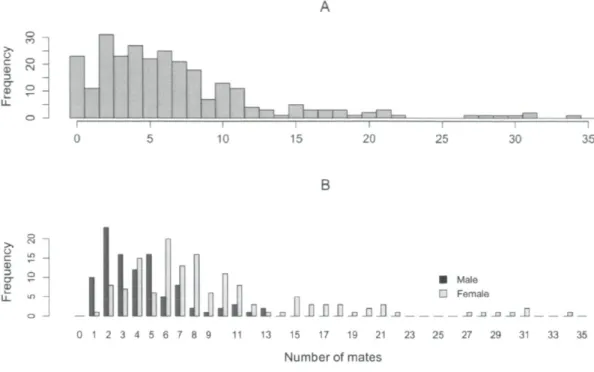
3 pools (•) where salmon were caught and released in 2009 21 Figure 2. Observed distribution of the number of inferred offspring (reproductive success)
in the Escoumins River for the 2009 spawning season when A) males and females are pooled together and B) when males (black) and females (grey) are separated. The sex of
salmon with no reproductive success remains unknown 32 Figure 3. Observed distribution of the number of mates in the Escoumins River for the
2009 spawning season when A) males and females are pooled together and B) when males (black) and females (gray) are separated. The sex of salmon with no reproductive success,
and thus with no mate, remains unknown 33 Figure 4. Model-averaged predictions (solid line) and their associated unconditional
standard errors (dashed lines) of grilse chances to access reproduction (i.e. to produce at least one offspring) as a function of the date of river entrance. Real observations are plotted
in gray 36 Figure 5. A) The effect of the interaction between C&R and size on the number of inferred
fry as illustrated by the model-averaged predictions (solid lines) and their associated unconditional standard errors (dashed lines) for C&R (black) and non-caught (gray) salmon. Plotted predictions are for females that entered the river in late July and who mated with 9 partners. B) Model-averaged predictions (solid lines) and their associated unconditional standard errors (dashed lines) of the number of inferred offspring for males (black) and females (gray) un-caught salmon of 78 cm that entered the river in late July...39 Figure 6. Model-averaged predictions of the number of inferred fry for C&R salmon that were exposed to air for 0 (solid line), 1-10 (dashed line) and >10 seconds (dotted line)
I. Introduction
Prologue
Dans les années 1860. une espèce précédemment commune et répandue de salmonidé, Salmo salmulus, disparut complètement de la surface du globe (Mills 1991). Bien que subite, la disparition de Salmo salmulus n'était pourtant attribuable ni à la surpêche, ni à la destruction de son habitat, mais bien aux scientifiques qui étudiaient l'espèce. En fait, les spécimens traditionnellement identifiés comme faisant partie de l'espèce se sont révélés être des tacons précoces de saumon atlantique {Salmo salar L.). L'étonnant dimorphisme exprimé par les mâles saumons d'une même population (les tacons précoces sont 10 fois plus petits que les mâles anadromes) est un exemple classique de tactiques de reproduction alternatives (ATRs) (Oliveira et al. 2008). L'existence de pareilles tactiques, tout comme le développement de caractères sexuels secondaires, dont l'impressionnante queue du mâle paon {Pavo sp.) est un spectaculaire exemple, résultent de l'action de la sélection sexuelle (Darwin 1871).
Sélection sexuelle et tactiques de reproduction alternatives
Selon la théorie de la sélection sexuelle telle qu'élaborée par Darwin (1871) dans son livre « The descent of man and selection in relation to sex », les mâles et les femelles d'une même espèce investissent différemment l'énergie qu'ils allouent à la reproduction. Bien qu'il existe des exceptions à la règle, les femelles investissent typiquement dans la production des rejetons, alors que les mâles rivalisent entre eux afin d'accéder aux gamètes produites par les femelles. Le succès reproducteur des femelles repose alors principalement sur la capacité de produire et de protéger ses rejetons ainsi que sur le choix du (ou des) meilleur partenaire possible. Pour les mâles, qui produisent une grande quantité de gamètes, le succès reproducteur est fortement corrélé à leur nombre de partenaires, ce qui résulte en une importante sélection intra-masculine (Bateman 1948). Conséquemment, la variance du succès reproducteur est typiquement plus grande chez les mâles que chez les femelles. La variance de succès reproducteur au sein des individus d'un même sexe (le plus souvent les mâles) permet l'apparition et l'évolution de tactiques de reproduction alternatives (ARTs) (Oliveira et al. 2008). On définit une ARTs comme étant un ensemble de traits sélectionnés
pour maximiser le fitness par une ou plusieurs voies alternatives dans le contexte d'une compétition intra-espèce et intra-sexuelle pour l'accès à la reproduction (Oliveira et al. 2008). Les ARTs sont présentes de manière ubiquitaire dans le règne animal et sont particulièrement abondantes chez les poissons (Taborsky 2008). Le saumon atlantique, de par la complexité de son système de reproduction et de par l'impressionnant dimorphisme exprimé par les mâles adoptant différentes tactiques, est l'une des espèces dont le système de reproduction est le plus étudié.
Le saumon atlantique {Salmo salar L.)
Distribution géographique
Le saumon atlantique (Salmo salar L.) est un salmonidé de l'ordre des salmoniformes que l'on retrouve naturellement dans les rivières de la côte est et ouest de l'Atlantique Nord. Historiquement, son aire de répartition s'étendait du Connecticut (E-U) jusqu'à la Baie d'Ungava (PQ) en Amérique du Nord et de la rivière Duoro (Portugal) à la rivière Pechora (Russie) en passant par les îles Britanniques, l'Irlande et le Groenland en Europe (Maccrimmon and Gots 1979, Parrish et al. 1998). Dans toute son aire de répartition, le saumon est présent, de manière sympatrique ou non, sous la forme anadrome et sous la forme résidente demeurant en eau douce (potamodrome) (Klemetsen et al. 2003) connue au Québec sous le nom de ouananiche. C'est traditionnellement la forme anadrome, migratrice, qui retient le plus l'attention du public puisqu'elle constitue les stocks les plus abondants et qu'on lui attribue une valeur économique supérieure (Verspoor et al. 2007).
Cycle de vie
Au moment de la reproduction, les saumons anadromes migrent généralement en direction de leur rivière natale après avoir séjourné typiquement de 1 à 3 hivers en mer à se nourrir (Klemetsen et al. 2003). On les nomme alors respectivement madeleineaux (1 hiver en mer) et rédibermarins (2 hivers ou plus en mer). Le frai se déroule en automne; les femelles creusent alors le gravier du lit de la rivière pour former un ou plusieurs nids regroupés en redds où elles déposent leurs œufs qui sont immédiatement fécondés par les mâles anadromes ainsi que par des tacons sexuellement matures. Les alevins sortent du nid l'été
suivant après avoir épuisé les réserves de leur sac vitellin et se déplacent ensuite vers les zones d'alevinage propices à leur alimentation. Lors de leur deuxième été, les alevins deviennent tacons et continuent de se nourrir en rivière pour une période variant de 1 à 7 ans selon la latitude (Klemetsen et al. 2003). Suivant cette période de croissance relativement lente, les tacons subissent une métamorphose physiologique les préparant à la vie en eau salée (smoltification). On les nomme alors saumoneaux et ils entreprennent une migration printanière vers la mer, la dévalaison, qui est déclenchée, en partie, par la température de l'eau (Byrne et al. 2003). Certains tacons mâles, appelés tacons précoces, adoptent une toute autre stratégie : plutôt que d'entreprendre la migration risquée en mer, ils deviennent matures sexuellement dès leur troisième été de vie, sans jamais quitter leur rivière natale. En mer, le comportement des saumons reste encore peu documenté. Généralement, les rédibermarins de l'Amérique du Nord et de l'Europe se nourrissent à l'ouest du Groenland alors que les saumons de la Scandinavie et de l'Europe utilisent également la mer de Norvège comme zone d'alimentation (Mills 1991). Les madeleineaux utilisent plutôt des zones d'alimentation plus près de leur rivière natale. Dans le cas des madeleineaux nord-américains, ces zones seraient situées au large de Terre-Neuve et du Labrador (Mills 1991).
Système de reproduction du saumon atlantique
Chez le saumon atlantique, en accord avec les prémisses de la sélection sexuelle, les mâles et les femelles utilisent différemment l'énergie allouée à la reproduction. Les femelles investissent six fois plus d'énergie dans la production de gonades que les mâles et elles utilisent principalement le reste de leur énergie au choix du site de frai et à la construction des nids (Fleming 1996). Le succès reproducteur des femelles serait corrélé à la profondeur des nids qu'elles creusent (évitant ainsi la destruction de leurs nids par superposition) ainsi qu'à leur fécondité, deux variables dépendantes de leur taille (Fleming 1996). De leur côté, les mâles consacrent une grande part de leur énergie afin d'accéder aux femelles. Au sein des mâles anadromes, qui luttent entre eux de manière agressive afin de monopoliser les femelles, la taille ainsi que les traits sexuels secondaires qu'ils arborent (coloration, kype, etc.) jouent un rôle important lors des combats sur les sites de frai (Fleming 1996). Les mâles précoces adoptent une toute autre stratégie ; ne faisant pas le poids face aux mâles
anadromes qui font près de dix fois leur taille, ils accèdent furtivement aux femelles au moment où ces dernières déposent leurs œufs et où les autres mâles sont distraits, occupés à déposer leur sperme. Bien qu'ils produisent un volume de sperme réduit si on les compare aux mâles anadromes, les tacons précoces investissent relativement plus d'énergie dans la production de leurs gonades, produisant ainsi un sperme plus compétitif que celui de leurs concurrents anadromes (Gage et al. 1995). De récentes études génétiques ont d'ailleurs révélé qu'en milieu naturel, les tacons précoces fécondent de 22 à 65% des œufs déposés par les femelles lors du frai (Garcia-Vazquez et al. 2001, Taggart et al. 2001, Saura et al. 2008, Weir et al. 2010). D'autres études ont proposé que contrairement à la théorie de la sélection sexuelle, la variance du succès reproducteur des mâles et des femelles était également élevée (Fleming et al. 1997, Garant et al. 2001). D'autre part, on sait aujourd'hui que le saumon atlantique est une espèce à la fois polygame et polyandre (Garant et al. 2001, Taggart et al. 2001). De récentes études suggèrent d'ailleurs que le nombre de partenaires serait une des variables qui expliqueraient le mieux le succès reproducteur du saumon et de la truite brune (Salmo truttd) (Garant et al. 2001, Serbezov et al. 2010). Malgré ces récentes découvertes, il n'y a, à ce jour et à ma connaissance, aucune étude qui a documenté le système de reproduction de l'ensemble d'une population de saumon atlantique dans une rivière nord américaine. Une bonne compréhension du système de reproduction et des facteurs déterminant le succès reproducteur en nature d'une espèce permet d'évaluer et ultimement, d'améliorer l'efficacité des programmes de gestion et de conservation la visant (Araki et al. 2008, Thériault et al. 2011). Dans cette étude, en plus de décrire le système de reproduction d'une population sauvage de saumon atlantique, nous nous attardons à mesurer l'impact de la remise à l'eau, une mesure de gestion qui croît en popularité, sur le succès reproducteur du saumon.
Pourquoi la capture de saumon atlantique doit-elle être
réglementée ?
Statut actuel du saumon atlantique
L'industrie aquicole produit annuellement plusieurs millions de saumons afin de garnir nos tables, si bien que plus de 94 % des saumons atlantique adultes présents sur terre vivent
aujourd'hui en captivité (Gross 1998). Bien que certaines populations sauvages de saumon atlantique, notamment au Canada et en Norvège, comptent encore plusieurs milliers d'individus (Dodson et al. 1998), la plupart, surtout celles situées au sud de l'aire de répartition de l'espèce, sont soit en déclin, ou encore carrément disparues (Parrish et al.
1998, Kellogg 1999, ICES 2011). Au Canada, les populations de l'intérieur de la Baie de Fundy sont d'ailleurs aujourd'hui protégées en raison de leur statut d'espèce en voie de disparition (COSEWIC 2010) alors que les populations de l'état du Maine sont protégées sous l'acte des espèces menacées (Species at Risk Act) aux États-Unis (NRC 2003). Au niveau mondial, l'abondance des stocks de saumon atlantique sauvage est en déclin depuis les trois dernières décennies et a maintenant atteint un creux historique (Hard et al. 2008, ICES 2011).
La pêche commerciale d'hier à aujourd'hui
D'abord utilisé par les différentes nations autochtones comme une importante source d'alimentation saisonnière dans l'Est du Canada, le saumon atlantique a par la suite soutenu une fructueuse pêche commerciale se déroulant notamment dans les estuaires des rivières de la Gaspésie et de la Côte-Nord du Québec (Dunfield 1986). Au 19e siècle, les revenus découlant de l'industrie de cette pêche en faisaient la deuxième pêcherie la plus lucrative derrière la pêche à la morue (Gadus morhua) (Chase 2003, Beaudin and Côté 2008). Au 20e siècle, la découverte des zones d'alimentation en mer du saumon atlantique entraîna l'ouverture de nouvelles pêcheries de stocks mixtes (une pêche touchant des saumons provenant de différentes populations) le long de la côte ouest du Groenland et autour des îles Féroé en mer de Norvège. C'est à la suite de l'ouverture de ces pêcheries, au cours des années 1960 et 1970, que les captures mondiales atteignirent leur maximum alors que 12 000 tonnes de saumons étaient pêchées annuellement (Crozier et al. 2004, Hindar et al. 2010). À la suite de la constante diminution de l'abondance des stocks de saumon atlantique dans les années 1970 et 1980, un programme de rachat des droits de pêche au saumon a été mis en place par le gouvernement canadien dans les années 1990s, ce qui mena à la fermeture complète de la pêche commerciale en 2000 (Chase 2003, FQSA 2004). Sur le plan international, l'Organisation pour la Conservation du Saumon de l'Atlantique Nord (OCSAN) gère depuis 1984 les pêcheries de l'ouest du Groenland et des îles Féroé
par un système de quotas fixés selon les recommandations du comité scientifique du Conseil International pour l'Exploration de la Mer (CIEM) (Crozier et al. 2004, Hindar et al. 2010). Au Groenland, un moratoire a été instauré et l'exploitation du saumon atlantique se limite maintenant à une pêcherie de subsistance (Crozier et al. 2004).
Impact de la pêche sportive sur les populations de saumon atlantique
L'essor de la pêche sportive à la mouche en Amérique du Nord est généralement attribué aux officiers de l'armée britannique qui auraient rapporté cette mode d'Europe au 19e siècle (Dunfield 1986). La réglementation encadrant cette pêche récréative fut d'abord inexistante et les premiers règlements furent pour le moins permissifs, comme en fait foi l'étonnante histoire de M. Strang, qui captura, au milieu du 19e siècle, 50 saumons en deux jours sur la rivière des Escoumins (Nettle 1857). Cent ans plus tard, au milieu du 20e siècle, la limite des prises était de 6 saumons par jour et de 21 par semaine (Maheux 1956). À la fin du 20e siècle, en parallèle avec le rachat des permis de pêches commerciales par les gouvernements, les autorités ont mis en place un système de gestion plus strict de la pêche sportive au saumon. En 1984, le gouvernement canadien imposait la remise à l'eau de tous les grands saumons (63 cm et plus) pris dans les rivières des provinces atlantiques (FQSA 2004). La même année, le Québec adoptait un système de gestion « rivière par rivière », où la remise à l'eau était, et est encore aujourd'hui, un outil de gestion important permettant la conservation de l'espèce sans forcer la fermeture de la pêche (FQSA 2004). Actuellement, un pêcheur détenant son permis au Québec peut garder 7 saumons annuellement. La limite journalière varie d'une rivière à l'autre et se situe entre 0 (la pêche est totalement fermée, ou seule la remise à l'eau est permise) et 2 saumons (MRNF 2012). Cette limite est déterminée selon un seuil de conservation qui correspond au nombre minimal d'oeufs requis devant être déposés dans une rivière de manière à obtenir une production optimale pour une population donnée (Caron et al. 2006). Une gestion adéquate de la pêche à la ligne est primordiale puisque, contrairement à la croyance populaire, la pêche sportive, au même titre que la pêche commerciale, représente une menace réelle à la conservation des espèces exploitées (Cooke and Cowx 2004). De plus, depuis la fermeture des pêcheries commerciales à l'ouest du Groenland et dans les eaux intérieures du Canada, la majorité (62 % en 2007) des captures américaines s'effectue en rivière (CIEM 2008). Au Québec, en
201 L 13 % des saumons remontant une des 114 rivières à saumon de la province ont été capturés par les pêcheurs sportifs (Dionne et al. 2012).
Pourquoi la remise à l'eau est-elle un outil de gestion et de
conservation efficace ?
La remise à l'eau comme outil de gestion et de conservation
L'abondance de plusieurs populations de saumon atlantique est donc en déclin et les pêcheurs sportifs exploitent aujourd'hui une part importante de cette ressource. D'autre part, la pêche sportive au saumon atlantique soutient actuellement une industrie rapportant annuellement plus de 100 millions de dollars dans l'est du Canada (POC 2007, ASF 2011). Dans ce contexte, il est primordial de protéger adéquatement ce poisson ayant une valeur biologique, culturelle et économique importante. La remise à l'eau est un des outils permettant aux gestionnaires de préserver la ressource sans forcer la fermeture de la pêche. Cette pratique se définit comme étant l'action de libérer à l'endroit où il a été capturé, et dans les meilleures conditions possibles, un poisson péché à l'aide d'un équipement de pêche sportive (Arlinghaus et al. 2007). Les pêcheurs du monde entier pratiquent aujourd'hui la remise à l'eau afin de se conformer à la réglementation en place, ou encore suivant une initiative personnelle vouée à la conservation de l'espèce. Selon 1TCES (2011), la remise à l'eau du saumon atlantique fait l'objet d'une popularité toujours grandissante au niveau mondial. En 2010, 222 000 saumons furent remis à l'eau, soit 44 000 de plus qu'en 2006 (ICES 2011). Depuis 1997, les pêcheurs au saumon du Québec peuvent se procurer un permis de remise à l'eau, à coût moindre que pour le permis de capture, leur permettant de pratiquer ce type de pêche sportive. En 2011 au Québec, 52 % des saumons capturés sur les rivières québécoises par pêche sportive furent graciés (Dionne et al. 2012).
Efficacité de la remise à l'eau
Bien qu'à première vue la remise à l'eau semble être une solution idéale, elle n'est pas sans détracteurs, particulièrement en Europe, au sein d'organisations pour la protection des droits des animaux, qui considèrent cette pratique comme étant cruelle (Aas et al. 2002). Nous nous en tiendrons ici au débat scientifique entourant la remise à l'eau. (Pour de plus
amples informations sur les aspects sociaux culturels, consultez Aas et al., 2002 et Arlinghaus et a l , 2007). L'efficacité de la remise à l'eau comme outil de gestion et de conservation repose sur la prémisse que le poisson capturé puis relâché survit à cet événement et se reproduit aussi bien qu'un poisson non péché. Les pêcheurs et les gestionnaires ont donc deux raisons les motivant à promouvoir cette pratique. Premièrement, un poisson relâché pourra éventuellement être capturé à nouveau, augmentant ainsi la valeur économique de chaque poisson. Dans une étude sur la pêche au saumon atlantique sur la rivière Alta en Norvège, Thorstad et al. (2003) rapportent que 4 % des saumons ont été capturés plus d'une fois dans une saison. Dans une étude semblable menée par Whoriskey et al. (2000) en Russie, sur la rivière Ponoi, 11 % des saumons furent pris deux fois et 0,5 % trois fois au cours de la saison de pêche. Deuxièmement, il est assumé qu'un poisson relâché participera à la reproduction, assurant ainsi le renouvellement des populations exploitées. D'un point de vue biologique, on considère donc que la remise à l'eau permettra au saumon gracié de se reproduire et d'obtenir un succès reproducteur comparable au saumon non péché ayant le même statut.
Est-ce que la remise à l'eau affecte la valeur sélective des
saumons graciés?
Valeur sélective : définition
Le concept de valeur sélective (fitness) vise à résumer les interactions complexes entre les individus d'une même population amenant certains génotypes et phénotypes à devenir relativement plus ou moins dominants, d'un point de vue démographique, dans cette population (Stearns 1992). On peut définir la valeur sélective comme étant le succès reproducteur relatif total d'un individu, c'est à dire sa capacité à produire des descendants qui survivront jusqu'à maturité, et qui, à leur tour, se reproduiront, transmettant ainsi leurs gènes aux générations futures. Il est particulièrement difficile de mesurer la valeur sélective d'un individu (génotype), car cela demande de suivre des individus sur plusieurs générations. De manière plus réaliste, on peut documenter différentes composantes de la valeur sélective. Notamment, Steams (1992) propose deux composantes soit le taux de survie et le taux de reproduction. Dans le cadre de l'étude de l'effet de la remise à l'eau, il
est utile de diviser la valeur sélective en trois composantes : (1) le taux de survie, (2) la migration, (3) la reproduction.
Taux de survie suivant un événement de remise à l'eau
Les estimés du taux de mortalité des saumons atlantique suivant un événement de remise à l'eau varient grandement d'une étude à l'autre et se situent entre 0 % et 80 % (Wilkie et al. 1996, Anderson et al. 1998, Dempson et al. 2002, Thorstad et al. 2003) avec une moyenne autour de 12 % (Bartholomew and Bohnsack 2005). Différents facteurs augmentant le taux de mortalité des salmonidés suivant la remise à l'eau ont été identifiés (Bartholomew and Bohnsack 2005). Entre autres facteurs relevés comme significatifs on dénombre: la présence d'ardillon sur l'hameçon (saumon coho ; Oncorhynchus kisutch) (Bartholomew and Bohnsack 2005), une faible concentration d'ions dissous dans l'eau (saumon atlantique) (Kieffer et al. 2002), la phase de migration (saumon atlantique) (Brobbel et al.
1996, Halttunen et al. 2010), mais voir (Thorstad et al. 2007), le temps d'exposition à l'air (Salvelinus fontinalis) (Schreer et al. 2005), l'interaction entre l'exposition à l'air et la température de l'eau (Gingerich et al. 2007), les techniques de manipulation des poissons (truite arc-en-ciel; Oncorhynchus mykiss) (Ferguson and Tufts 1992). la profondeur où l'hameçon s'est accroché (Schill 1996) et finalement, une température de l'eau élevée (saumon atlantique et truite arc-en-ciel) (Booth et al. 1995, Brobbel et al. 1996, Wilkie et al. 1996, Thorstad et al. 2003, Boyd et al. 2010). Ce dernier facteur semble être particulièrement important pour le saumon atlantique puisque les taux de mortalité les plus élevés (40 % et 80 %) ont été observés alors que la température de l'eau approchait 22°C (Wilkie et al. 1996, Anderson et al. 1998). En effet, lorsque la température de l'eau dépasse 18°C, le risque de mortalité relié à la pêche augmente de façon exponentielle chez le saumon atlantique (Thorstad et al. 2003). À Terre-Neuve, les rivières à saumon sont d'ailleurs fermées à toutes formes de pêches, remise à l'eau comprise, lorsque la température atteint ou dépasse les 22 C (Dempson et al. 2001).
Effet de la remise à l'eau sur la migration en rivière
Dans l'Est du Canada, la montaison des grands géniteurs (saumons anadromes) s'étale de juin à octobre. La montaison peut normalement être divisée en trois phases distinctes : (1)
10
un déplacement relativement rapide vers l'amont de la rivière qui est typiquement ponctué de périodes de repos plus ou moins longues; (2) une phase de recherche au cours de laquelle les saumons se déplaceraient de manière erratique tout en demeurant près du site de frai et; (3) une longue période de résidence sur le site de frai (Okland et al. 2001). Ces périodes de migration et le passage d'obstacles sont par ailleurs influencés par des facteurs extrinsèques comme le débit et la température de l'eau (Jensen et al. 1986, Erkinaro et al.
1999).
En raison des difficultés techniques que cela engendre, il existe peu d'études évaluant l'impact de la remise à l'eau sur les salmonidés en milieu naturel. La majorité des études relatives à la remise à l'eau du saumon atlantique a été réalisée en milieu contrôlé afin de détecter les effets physiologiques, létaux ou non, de cette pratique (Thorstad et al. 2003). L'utilisation de la radio télémétrie permet toutefois de mesurer l'impact de la remise à l'eau sur le comportement migratoire des saumons en milieu naturel (Pollock and Pine 2007, Donaldson et al. 2008). À ma connaissance, six études ont été publiées à ce sujet (Makinen et al. 2000, Whoriskey et al. 2000, Thorstad et al. 2003, Thorstad et al. 2007, Halttunen et al. 2010, Jensen et al. 2010). Dans la majorité de ces expériences, les scientifiques ont observé des comportements anormaux chez tous les saumons suivis. Ces études ont permis de montrer que les saumons semblent réagir à la remise à l'eau en migrant vers l'aval de la rivière ou en retardant leur migration. Les répercussions de cette dévalaison dépendent du stade de migration. Lorsque les saumons étaient capturés lors de la première phase de migration, leur montaison était retardée de 28 à 34 jours (Makinen et al. 2000, Thorstad et al. 2007). Selon Fleming (1996), les saumons n'auraient pas d'avantages adaptatifs à arriver tôt sur les sites de frai, réduisant ainsi l'effet potentiel de la remise à l'eau sur la valeur sélective. Par contre, si une arrivée hâtive procure au saumon un quelconque avantage compétitif (au niveau de la dominance ou du nombre d'occasion de reproduction chez les mâles ou du choix de site pour les femelles), le retard causé par l'événement de remise à l'eau pourrait influencer la valeur sélective. Chez les salmonidés comme chez plusieurs autres animaux, il semble que les mâles arrivent plus tôt que les femelles sur les sites de reproduction et que cette arrivée hâtive pourrait leur apporter quelques avantages en terme de succès reproducteur. Par exemple, il peut être avantageux pour un mâle de se reproduire avec une vierge puisque le nombre d'œufs déposés par les femelles à chaque
11 oviposition diminue au cours de la saison du frai (Fleming and Reynolds 2003). De plus, la date d'entrée en rivière, très variable chez le saumon (Klemetsen et al. 2003), serait en partie héréditaire (Hansen and Jonsson 1991) et pourrait correspondre à une adaptation dont les avantages seraient perdus en cas de retard attribuable à la pêche. Chez les saumons capturés tard dans la saison, la période de résidence sur les sites de frai n'était pas observée, les saumons bougeaient plutôt de manière erratique jusqu'au moment du frai (Thorstad et al. 2003). Donc, si la période de résidence permet au saumon d'établir sa dominance sur un bon site de reproduction, la remise à l'eau aurait un effet négatif sur la reproduction des poissons péchés tard en saison. Finalement, une autre conséquence probable de la remise à l'eau suggérée dans ces études serait que les saumons, retardés ou perturbés, n'atteignent pas leur section natale de rivière, diminuant ainsi l'avantage de l'adaptation locale probable à une petite échelle spatiale (Thorstad et al. 2007). Les saumons suivis dans cette étude se retrouvaient toutefois sur des sites de frai connus au moment de la reproduction (Thorstad et al. 2007). Ces six études ont été menées sur des rivières Scandinaves ou subarctiques à fort débit (la rivière Alta en Norvège, d'où quatre des six études citées proviennent, draine un bassin versant de 74000 km. et a un débit moyen de 75 rro/s (Thorstad et al. 2003)) et avec une température moyenne et maximale beaucoup moins élevée que celles observées lors de la saison de pêche sur l'ensemble des rivières du sud du Québec (en août, la température de la rivière Alta atteint 14-16°C (Thorstad et al. 2003) alors que sur la rivière des Escoumins (le site de la présente étude), elle dépasse fréquemment les 22°C à la même période). Il est donc incertain que les conclusions de ces études européennes soient applicables au Québec, du moins au sud de la province. Aussi, les premières études télémétriques souffraient du manque d'un groupe témoin rendant hasardeux de départager l'effet du marquage des individus suivis de l'effet de la variable d'intérêt, soit l'événement de remise à l'eau. D'ailleurs, dans des travaux récents avec groupe témoin, Jensen et al. (2010) ont observé que le comportement de dévalaison normalement associé à la remise à l'eau semblait plutôt normal alors qu'aucune différence de comportement n'a été relevée entre le groupe remis à l'eau et le groupe témoin.
12
Effet de la remise à l'eau sur la reproduction
Chez le saumon atlantique, chaque événement de reproduction est associé à un coût énergétique important. C'est en effet entre 35 % à 70 % de leur énergie totale que les saumons atlantique adultes, mâles ou femelles, investissent pour la migration et la reproduction (Jonsson et al. 1997, Jonsson and Jonsson 2003). Contrairement au saumon du Pacifique (genre Oncorhynchus) qui est sémelpare, le saumon atlantique est itéropare et peut donc se reproduire plus d'une fois au cours de sa vie (Klemetsen et al. 2003). Ce n'est toutefois généralement qu'une faible proportion des saumons anadromes (11 % en moyenne (Fleming 1998)) qui se reproduisent effectivement plus d'une fois, et de ce nombre, les femelles sont de loin les plus nombreuses (Mills 1991, Fleming 1996).
Il est reconnu qu'au moment d'entrer en rivière, les saumons atlantique anadromes entreprennent un jeûne qui ne se termine qu'au printemps suivant, lors du retour en mer (Kadri et al. 1995). Ils sont alors contraints de puiser dans leurs réserves d'énergie somatique afin d'assurer le développement de leurs gonades et leur migration jusqu'aux frayères (Jonsson et al. 1997). L'énergie totale emmagasinée par un saumon au moment de son entrée en rivière détermine l'investissement de cet individu dans la migration et la reproduction et serait donc un bon indicateur de son potentiel de reproduction (Jonsson et al. 1997). La perte d'énergie et le stress subi par un saumon lors de sa lutte avec un pêcheur pourraient donc affecter négativement son succès reproducteur.
Tout de même, le saumon atlantique gracié semble avoir un certain succès au moment de la reproduction selon certaines études. Le nombre de nids de saumon observé dans la rivière Alta en Norvège a augmenté d'un facteur 2.3 à la suite de l'introduction de la remise à l'eau obligatoire (Thorstad et al. 2003). Un effet comparable a été enregistré sur la rivière Ponoi en Russie où le nombre de juvéniles a augmenté à la suite de l'implantation de la remise à l'eau (Whoriskey et al. 2000). Également, une étude en laboratoire suggère que la survie et la viabilité des gamètes ne sont pas différentes entre les saumons péchés ou non (Booth et al. 1995).
Par contre, à ce jour, malgré le fait que le saumon soit une des espèces les mieux étudiées en ce qui a trait à la remise à l'eau (Cooke and Suski 2005), aucune étude permettant de
13 mesurer directement 1*impact de la remise à l'eau sur le succès reproducteur du saumon atlantique en milieu naturel n'a été réalisée. Il existe pourtant un consensus dans la littérature actuelle soutenant la nécessité d'entreprendre des études permettant de quantifier l'effet de la remise à l'eau sur la valeur sélective individuelle et ultimement, sur la persistance des populations de saumons à plus long terme, des informations essentielles permettant l'utilisation optimale de la remise à l'eau comme outil de gestion (Arlinghaus et al. 2007, Pollock and Pine 2007). En effet, pour l'instant au Québec, tous les saumons péchés et remis à l'eau sont considérés comme des géniteurs potentiels au même titre que ceux n'ayant pas été péchés. Au Nouveau-Brunswick, on considère une mortalité variant de 3 à 5 % au sein des saumons graciés (B. Chaput, communication pers.). L'interprétation des gestionnaires du taux de survie et de la capacité de se reproduire des saumons graciés peut donc avoir un impact non négligeable sur l'atteinte des objectifs de conservation en rivière (si on considère un taux de mortalité de 12 % associé à la remise à l'eau, 1488 saumons ont faussement été identifiés comme géniteurs sur les rivières québécoises en 2011, et ce, sans compter les saumons qui ont survécu, mais qui ont potentiellement subi une diminution de valeur sélective). Il serait donc avantageux de quantifier plus précisément l'influence de la remise à l'eau sur le succès reproducteur des individus et ainsi ajuster les mesures de gestion en conséquence. Toutefois, comme le souligne Cooke et Suski (2005), les études portant sur la valeur sélective en lien avec la remise à l'eau sont très difficiles à réaliser en nature. La difficulté majeure de telles études, comme pour l'étude des systèmes de reproduction en milieu naturel, réside dans la mesure du nombre de descendants produits par chaque individu. Les estimés de succès reproducteur étaient traditionnellement obtenus de manière indirecte, par interprétations découlant d'observations comportementales au moment du frai. Cette approche demande de longues heures d'observation, rendant l'étude au niveau populationnelle pratiquement impossible en nature, et peut donner des résultats trompeurs. Il peut en effet y avoir une différence importante entre le nombre de copulation effectuée par un individu et le nombre de rejetons effectivement produits (Gibbs et al.
14
Assignation parentale grâce aux marqueurs moléculaires polymorphes
Le développement de marqueurs moléculaires codominants hautement polymorphiques (plus particulièrement des microsatellites depuis la fin des années 1980) et leur application aux analyses d'assignation parentale ont permis aux scientifiques de mesurer le succès reproducteur d'individus en milieu naturel (Avise 2004). Les microsatellites sont de courtes séquences d'ADN formées de répétitions en tandem de deux, trois ou quatre nucleotides et sont distribués dans l'ensemble du génome. Le taux de mutation (ajout ou perte d'éléments répétés) élevé au sein des séquences microsatellites (10"3 à 10~5mutations/locus/génération)
résulte en une grande diversité allélique dans une population. Avec un nombre approprié de loci microsatellites suffisamment polymorphes (Bernatchez and Duchesne 2000) et en considérant leur mode de transmission mendélien, il est possible de réassigner un jeune à ses parents biologiques en comparant son génotype multilocus à celui de ses parents potentiels. Chez les salmonidés, l'assignation parentale moléculaire s'est avérée être un outil puissant qui a, entre autres choses, permis de démystifier les patrons de reproduction du saumon atlantique, de la truite brune (Salmo trutta) et de la truite arc-en-ciel (Oncorhynchus mykiss) (Garant et al. 2001, Seamons et al. 2004, Serbezov et al. 2010), de mieux comprendre la présence de tactiques alternatives de reproduction chez le saumon et l'omble de fontaine (Salvelinus fontinalis) (Garant et al. 2002, Garant et al. 2003, Thériault et al. 2007) et de mettre en évidence l'impact de la domestication sur la valeur sélective de la truite arc-en-ciel (McLean et al. 2003, Araki et al. 2007a, Araki et al. 2007b).
Objectifs
Deux objectifs principaux sont visés par cette étude. Premièrement, nous visons à décrire le système de reproduction du saumon atlantique pour l'ensemble d'une population sauvage d'Amérique du Nord et à identifier les déterminismes du succès reproducteur individuel. Deuxièmement, nous visons à mesurer l'impact de la remise à l'eau sur le succès reproducteur du saumon atlantique et testons l'impact de la température de l'eau et de l'exposition à l'air sur l'efficacité de cet outil de gestion et de conservation.
II. Does catch and release affect the mating system and
individual reproductive success of wild Atlantic salmon
{Salmo salar L.)?
Antoine Richard1, Melanie Dionne2, Jinliang Wang3 and Louis Bernatchez1
1 Institut de Biologie Integrative et des Systèmes (IBIS), Pavillon Charles-Eugène
Marchand, Université Laval, Québec (QC), G1V 0A6, Canada
■y
Ministère des Ressources Naturelles et de la Faune du Québec (MRNF), Direction de la faune aquatique, Québec (QC), GIS 4X4, Canada
3 Institute of Zoology, Zoological Society of London, Regent's Park, London, NW1 4RY,
16
Résumé
L'utilisation des marqueurs moléculaires permet de décrire le système de reproduction et d'étudier les déterminismes du succès reproducteur (SR) individuel des espèces où l'observation du comportement reproductif est ardue et parfois trompeuse. Dans cette étude, nous tirons avantage des avancées en biologie moléculaire et des méthodes d'assignation parentale afin de décrire le système de reproduction et d'identifier les déterminismes du SR individuel du saumon atlantique (Salmo salar L.). De plus, nous utilisons les résultats d'assignation parentale afin de mesurer l'impact de la remise à l'eau, un outil de gestion et de conservation qui croît en popularité, sur le SR du saumon atlantique. Nous avons observé une importante variance du SR chez les deux sexes, mais la variance standardisée observée au sein des mâles (2,86) était plus importante qu'au sein des femelles (0,73). Mâles et femelles sont majoritairement polygames, ayant respectivement de 1 à 13 et de 1 à 34 partenaires, et le nombre de partenaires est d'ailleurs positivement corrélé au SR, cette relation étant plus importante chez les mâles. Les tacons précoces ont participé activement à la reproduction, produisant 44% des alevins réassignés. Les saumons remis à l'eau, qui représentaient 20% des rédibermarins de la rivière, ont produit 22% des alevins inclus dans nos analyses. La remise à l'eau a un faible impact négatif sur le nombre d'alevins produit par un individu et l'ampleur de cet impact est directement proportionnel à la taille de l'individu. Outre la taille, l'interaction entre la température de l'eau et l'exposition à l'air a également un impact négatif sur le SR d'un saumon remis à l'eau. Nos résultats améliorent notre compréhension du système de reproduction du saumon atlantique et confirment que, conditionnellement à de bonnes pratiques de remise à l'eau, les saumons remis à l'eau se reproduisent et participent à la pérennité des populations exploitées.
17
Abstract
The use of molecular markers enables the study of reproductive success in species for which behavioral observations are difficult and can be misleading. In this study, we took advantage of a river where we could genetically sample every returning adult in order to document in details the breeding system of a wild population of Atlantic salmon (Salmo salar L.) and to assess the determinants of salmon individual fitness. A second objective was to assess the impacts of catch and release (C&R), a widely used conservation and management tool, on mating and reproductive success. Both sexes showed high variance in individual reproductive success and, in accordance with classic sexual selection theory, estimated standardized variance was higher for males (2.86) than for females (0.73). Mature male parr sired 44% of the analyzed offspring. We found a weak but positive relationship between body size and fitness. Yet, male and female fitness was strongly and positively correlated with the number of mates, and that relationship was stronger among males. Females mated with 1 to 34 partners (including mature parr) while males mated with
1 to 13 different females. Catch and released salmon sired 22% of the progeny while representing 20% of the multi-sea-winter segment of the population in the river. We found that C&R had a mitigated impact on a salmon capacity to reproduce. However, C&R impact on the number of produced offspring was dependant on fish size as larger fish reproductive success was more impaired than smaller ones. Finally, we detected an interactive negative effect of water temperature and air exposure duration on reproductive success of caught and released salmon. This study represents one of the very few that have documented the mating system and determinants of reproductive success at the scale of a whole population in Atlantic salmon and as such, contributes to further improve our understanding of the species' complex reproductive biology. This study is also the first to have investigated the impact of catch and release on reproductive success, which shows that C&R represents an efficient management tool to promote conservation of salmon populations with limited negative impact on mating and reproductive success, pending appropriate C&R practices.
IX
Introduction
Studies of breeding systems enable us to understand the processes involved in sexual selection, which in turn determines the different reproductive strategies adopted by both sexes of a species. In particular, variance in reproductive success is typically thought to be higher among males than among females, which is interpreted as reflecting intra-masculine selection resulting from the strong correlation, in males, between the number of mates and fertility (Bateman 1948). Intra-sexual selection drives the evolution of alternative reproductive tactics (ARTs) (Oliveira et al. 2008) which are particularly frequent in fish (Taborsky 2008). Namely, species of the salmonid family are well known for the great diversity of their breeding systems and for the spectacular morphological differences expressed by males adopting different ARTs (Fleming 1998). In Atlantic salmon (Salmo salar L.) males may either mature as parr, which never leaves their natal rivers, or after one (they are then called grilse and measure < 63 cm) or two winters at sea (large salmon > 63 cm), whereas females normally mature after two winters at sea (Klemetsen et al. 2003). Early parentage genetic analysis on the species breeding system revealed an unexpected high level of polygamy for both sexes (Garant et al. 2001, Taggart et al. 2001) and, in apparent discordance with sexual selection theory (Andersson 1994), an equally high variance in reproductive success for both sexes (Fleming et al. 1997, Garant et al. 2001). Other studies have found differences in reproductive success (RS) realized by males adopting different reproductive strategies. For instance, Garant et al. (2003) showed that large, multi-sea-winter, salmon had higher RS than grilse. Previous studies also confirmed that mature parr, though at least one order of magnitude smaller than their anadromous counterparts, actively participate in reproduction fertilizing up to 60% of the eggs/fry in a given population (Taggart et al. 2001, Saura et al. 2008, Weir et al. 2010). Despite these findings, to date, there is to our knowledge no study that documented the mating system and reproductive success at the scale of a whole Atlantic salmon population in a natural North American river. An in-depth understanding of the factors determining mating system and reproductive success in nature may in turn contribute to improve the efficiency of management and conservation programs (Araki et al. 2008, Theriault et al. 2011).
19 Despite the concerns about the species status (COSEWIC 2010, ICES 2012), Atlantic salmon remains a highly prized game fish and many populations still sustain important recreational fisheries. In order to maintain the socioeconomic benefits resulting from angling while reducing its impact on wild population restoration, catch and release (C&R) angling has been introduced in many Atlantic salmon rivers draining in the North Atlantic. While the percentage of the total rod catch that is released differs greatly from one country to another, the total number of released salmon is increasing yearly and was over 206 000 in 2011 (ICES 2012). Catch and released salmon represented 59% of the total catches in Canada in 2011. In this country, C&R is mandatory for large salmon (> 63 cm fork length) since 1984, with the exception of the province of Québec, where large salmon retention is allowed only on rivers sustaining healthy populations that comply their conservation limit (Dionne et al. 2012, MRNF 2012).
The effectiveness of C&R as a conservation/management tool relies on the assumption that released fish do not only survive, but also reproductively contribute to the preservation of the exploited populations (Arlinghaus et al. 2007). Previous studies show that post release survival is highly variable and seems to be mainly related to air exposure and water temperature (Bartholomew and Bohnsack 2005, Cooke and Suski 2005, Arlinghaus et al. 2007, Gingerich et al. 2007). Experiments performed at low water temperature reported high (up to 100%) survival rates suggesting that when done in favorable conditions, C&R salmon do survive (Booth et al. 1995, Wilkie et al. 1996, Wilkie et al. 1997, Anderson et al. 1998, Whoriskey et al. 2000, Dempson et al. 2002, Thorstad et al. 2003, Boyd et al. 2010). However, to date, most sublethal C&R studies focused on physiological and behavioral aspects but never on reproductive success in natural settings (Thorstad et al. 2008).
Although essential to our understanding of salmon response to C&R, previous sublethal studies leave unanswered one of the crucial questions for a manager: does C&R impairs salmon fitness? Although Atlantic salmon is one of the most studied species in terms of the impact of C&R (Cooke and Suski 2005), there is, to date, no direct evidence that, when surviving, released salmon successfully reproduce. Anadromous mature Atlantic salmon migrate to rivers for the sole purpose of reproducing. The energetic cost of spawning and of the related upstream migration represent an investment of up to 70% of their body reserve
20
prior to fresh water entrance (Jonsson et al. 1997) resulting in a low level of iteroparity. Indeed, less than 10% of the anadromous Atlantic salmon reproduce more than once (Fleming 1998). It is therefore essential to quantify the impact of C&R on salmon reproductive success throughout the spawning period following release. Indirect measurement of one aspect of reproductive success of released salmon was made in the laboratory where the eggs from C&R and non-caught salmon showed no difference in terms of survival (Booth et al. 1995). In nature, the introduction of mandatory C&R resulted in a 2.3-fold increase in the number of spawning redds on the Sautso River in Norway (Thorstad et al. 2003). Similar observations were made in Russia, where Whoriskey et al. (2000) observed higher parr and fry densities on the Ponoi River after the introduction of C&R. These observations, however, cannot confirm the participation of C&R salmon in the reproduction, as too many factors possibly contributing to the observed increase were not controlled. The lack of information on sublethal effects of C&R on individual fitness, and therefore on population long term impacts, is not specific to Atlantic salmon (Arlinghaus et al. 2007) and can be explained by the difficulty to undertake fitness studies, especially in natural settings (Cooke and Suski 2005). Over the last two decades, the use of highly polymorphic genetic markers such as microsatellite, together with the development of parentage statistical method, led the way to the analysis of breeding system and of individual reproductive success in nature (Avise 2004, Jones et al. 2010).
The purpose of this paper is to document the genetic mating system at the scale of an entire population of North American Atlantic salmon and to determine the factors affecting individual reproductive success. We also established the variance in reproductive success for both sexes and related them to their respective reproductive strategies. Secondly, we aimed at quantifying for the first time the impact of C&R on salmon reproductive success in nature. More precisely, based on previous C&R studies, we expected C&R to impact on the potential to reproduce and reduce the number of produced offspring. Finally, we also tested the impact of air exposure and water temperature on C&R salmon fitness.
21
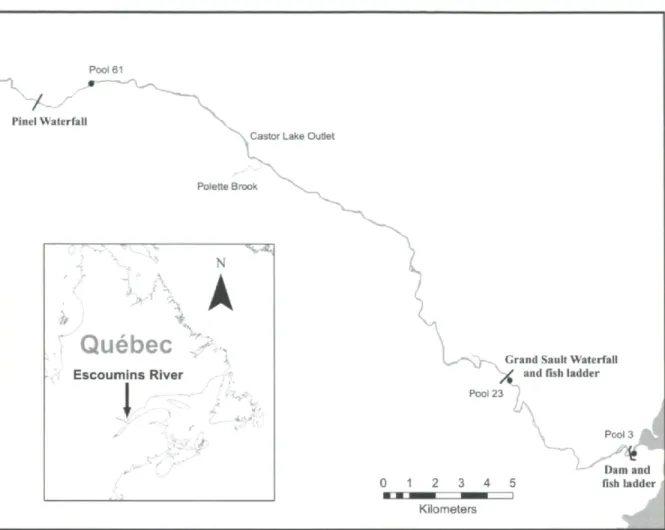
Materials and methods
Pool 61
Castor Lake Outlet
-Grand Sault Waterfall and fish ladder
0 1 2 3 4 5 Kilometers Pool 3
_^¥
Dam and fish ladderFigure 1. The Escoumins River, and its tributaries, on the north shore of the St-Lawrence River in Québec showing the positions of the obstacles to salmon migration, as well as the 3 pools (•) where salmon were caught and released in 2009.
Study site
The study was conducted on the Escoumins River, located on the north shore of the St.Lawrence River, in Québec, Canada (fig. 1 ). Mean annual flow 9 km upstream from the outlet is 14,8 m3s"'. The river is 84 km long with a catchment area of 800 km2. During
summer months, water temperature frequently exceeds 20°C and can reach 26°C. In 1901, a dam was built 1 km upstream of the river mouth, just upstream of tidal influence. In 1969, a fish ladder was built to allow the passage of Atlantic salmon. In the same year, a second fish ladder was built at the Grand Sault waterfall, a natural obstacle to upstream salmon
22
migration situated 9 km upstream of the dam, thus giving salmon access to 2 tributaries and a total of 36 km of river ending at Pinel waterfall, an impassable obstacle to wild salmon migrating from the sea. Restoration of the salmon population was also undertaken by means of fry stocking. Hatchery reared fry (3250 in 2009) resulting from adults that are captured in the Escoumins River are released in the river section located upstream of the Pinel waterfall. The river is now home to a small Atlantic salmon population (annual mean of 312 ± 100 returning salmon for the 2004-2009 period).
Salmon fishing (fly-fishing only) on the river is open from June 1st until September 15th.
All fishermen must register at the "ZEC (Zone d'Exploitation Contrôlée or Area of Controlled Exploitation) de la rivière des Escoumins", a local organization that manages the exploitation of the river. The river is divided in 4 fishing sectors and is composed of 66 pools easily accessible by a road that runs along the river. Provincial regulation allows fishermen to keep up to 2 grilse (salmon of < 63 cm fork length) a day, but they must release all large salmon (> 63 cm).
S a m p l e collection Adult sampling
In 2009, all Atlantic salmon entering the Escoumins River were measured and sampled (punch of 5mm diameter from the adipose fin) at the fish ladder situated 1 km from the mouth of the river. During measurement and sample collection, fish were kept in the water and free to swim, so it was only possible to measure length to the nearest 5 cm. Adult hatchery salmon (n=8) that produced the fry used to stock the river in 2010 were also sampled.
Caught and released fish sampling
Fishermen collaboration was solicited throughout the 2009 fishing season to collect tissue samples (punch of 5mm diameter from the adipose fin) and information from catch and release events. When registering, all fishermen received punch pliers and indications on how to collect sample on the fish they would eventually release. They were also receiving a few fishermen kits. Each kit contained a 1,5 ml Eppendorf filled with 95% ethanol to
23
preserve the adipose punch and a questionnaire to record information about the catch and release event. In the questionnaire, fishermen were asked to record, for each C&R, the date, the time of the day, the pool, the duration of the event (from hooking to landing), the kind of hook used (single or double, barb or barbless), the hooking emplacement, presence of bleeding, air exposure duration, condition at release (a subjective evaluation as whether the fish was in excellent, good or bad shape) and any other observations about the fish condition (injuries, etc). Since water temperature may influence catch and release issue (Thorstad et al. 2003), temperature data loggers were placed in all important pools of the river. By combining the information provided by the fishermen in the questionnaire and the data recorded by the temperature data loggers, it was possible to find the exact prevailing water temperature for each event. Fishermen participation was not mandatory, but strongly recommended. To promote participation, a salmon fly was given in exchange of each returned sample and the scientific team was consistently present on the riverbank promoting the project and assisting fishermen in the sample collection.
Fry sampling
In the 2010 summer, from August 2nd to 24th, electrofishing was performed on a total of 94 preselected rearing sites to capture a large number (n=2577) of fry produced by the fish that had entered the river to reproduce in 2009. Sites were preselected as a function of the quality of the habitat and to be representative of the river and its tributaries. To control for an eventual bias due to the sampling date of a site, two teams of two experienced technicians were simultaneously fishing the river: sites that were visited in a day were randomly chosen in the first half of the river for the first team and in the second half of the river for the second team. Each site was approximately 100 m long and 2 m wide and was electro-fished only once, with the exception of 13 sites with larger fry (age 0) that were initially misidentified as parr (age 1 ) and thus not collected. Those sites were revisited once and fished to collect only the large fry that were initially discarded. Each of the 2577 fry sampled was measured (to the nearest mm), weighted (± 0.02 g) and then preserved in 95% ethanol for later genotyping.
24
Genetic analysis
DNA was extracted from adult adipose fin tissue and from fry caudal fin tissue using a salt-based method (Aljanabi and Martinez 1997). Microsatellite polymorphism was analyzed at
12 loci: SSal71, SSal97, SSa202, SSa85 (O'Reilly et al. 1996), SSOSL417 (Slettan et al. 1995), SSaD85 (T. King, unpublished), SSaD71, SSaD58, SSaD486 (King et al. 2005), SSsp2201, SSspG7 (Paterson et al. 2004) and MST-3 (Presa and Guyomard 1996). Polymerase chain reaction (PCR) amplifications were performed in 2 multiplexes (annexe 1). The same PCR protocol was used for both multiplexes and were performed on Biometra® Tl thermocycler or Biometra® Tgradient. Cycling conditions were as follows:
15 minutes at 95°C to activate the QUIAGEN kit, 34 cycles of 94°C for 30 sec. (denaturation), 58°C for 3 min. (annealing), 72°C for 60 sec. (elongation) and a final elongation step at 60°C for 30 min. DNA amplified fragment were electrophoresed on a ABI™ 3130 (Applied Biosystems) and results were subsequently analyzed with GENEMAPPER™ software. Software implanted automatic scoring option was used, but alleles were visually inspected twice to minimize scoring errors.
To control for possible human errors and sample contamination, negative and positive controls were placed in each 96 samples plate used for PCR and electrophorèse. Presence of null allele, large allele dropout and the scoring of stutter peaks were estimated using
MICRO-CHECKER 2.2.3 (Van Oosterhout et al. 2004). Fis at each locus, and the significance
of either excess or deficit of hétérozygote, were computed using FSTAT (GOUDET 1995).
Finally, to estimate the genotyping error rate, all the samples from 2009 (the putative parents) were re-amplified, re-electrophoresed and re-analyzed with GENEMAPPER.
P a r e n t a g e analysis
First step: Identity check analysis
We used the identity check function implemented in CERVUS 3.0.3 (Kalinowski et al. 2007)
to find matching genotypes in our data set. Results from this analysis allowed us to identify fish that had been C&R more than once. Identity analysis was also used to determine if a
25 C&R salmon crossed the fish ladder and. if so. to associate the information recorded in the fish ladder (length and date of river entrance) to the C&R salmon.
Second step: sex determination
Since sexing of the putative parents at their passage in the dam fish ladder was most of the time impossible, and since having sexed putative parents greatly improves the power of parentage analysis, sex determination was done with the help of a preliminary parentage analysis and of the known sex ratio for grilse and large salmon of the Trinité River, an
index river situated 190 km northeast of the Escoumins River. First, using CERVUS we
found fry (n=1134) for which the most probable trio (fry, parent #1 and parent #2) fitted perfectly, i.e. 0 loci mismatching, and for which the second most probable trio showed at least 2 mismatching loci. It was then possible to find the mating partners of each parent present in the 1134 trios, thus creating a mating matrix (one row per individual, the first column contains the ID of that individual and subsequent columns contain the ID of its mates). The created mating matrix contained 2 clusters (i.e. groups of individuals that are linked in the pedigree). In a given cluster, the sex of any single individual determines the sex of all other individuals belonging to that cluster (otherwise we would have male-male or female-female pairs) so there are only two possible configurations regarding the sex of the individuals of each cluster, resulting in four possible configurations for the whole matrix. Using the known sex ratio of grilse and large salmon of the Trinité River (grilse =
11.5 % female, large = 93,8% female, for the 1992-2010 period) and the length, and thus the type (grilse or large), of each salmon in the matrix, we computed the log of the probability of each of the four configurations and retained the most probable configuration
(which was more than 1010 time more probable than the second most probable
configuration). Using this method, it was possible to find the sexes of the 235 parents present in the 1134 trios (out of a total of 268 adults). We have high confidence in these
attributed sexes since the exclusion approach used with CERVUS was extremely
conservative (we retained much less couples than what proposed by CERVUS at a 95%
confidence level) and that the sex ratio of the Escoumins River is likely similar to the one observed in the Trinité River. Namely, the vast majority of the grilse captured on the Escoumins River are reported to be males.
26
Third step: Assignment with PASOS, CERVUS and COLONY
In order to reduce the computation time required by COLONY (Jones and Wang 2010) to
analyze our large data set, CERVUS and PASOS (Duchesne et al. 2005) were first used to find
highly probable mother-offspring and father-offspring pairs that were then identified in the input file of COLONY as known maternity and known paternity. The only pairs that were
retained were pairs that fitted perfectly (0 mismatch) and that were selected both by
CERVUS and PASOS as the more probable pair for a specific offspring. A total of 1247
known paternity and 2395 known maternity were identified that way. Then, the full likelihood approach implemented in COLONY was used to allocate a mother to the 153 fry
that were not in the known maternity file and to allocate a father to the 1301 fry that were not in the known paternity file. Since we sampled absolutely all anadromous salmon that entered the river in 2009, and considering that female mature parr are extremely rare in Atlantic salmon, the probability that an actual mother was included in the candidates female file was set to 1. The probability that an actual father was included in the candidates male file was set to 0.6 as suggested by the results of preliminary assignments performed with PASOS and CERVUS and in agreement with the participation of mature parr in egg production reported previously (Garcia-Vazquez et al. 2001, Weir et al. 2010).
R e p r o d u c t i v e success analysis
Description of the mating system of Atlantic salmon
The best configuration given by COLONY was used to determine the number of fry produced
by each anadromous salmon that entered the river in 2009 and by mature parr. Although
mature parr were not sampled, COLONY could define their genotypes from the pedigree
analysis. Results from COLONY also allowed to find the number and type of mate
(anadromous or/and mature parr) for each salmon. The total number of fry inferred to mature parr was used to estimate the relative contribution of mature parr and anadromous male to the production of fry in the river.