Rôle de DSCAM dans le développement du circuit locomoteur
spinal
Thèse
Louise Thiry
Doctorat en neurobiologie
Philosophiae Doctor (Ph. D.)
Québec, Canada
© Louise Thiry, 2018
Rôle de DSCAM dans le développement du circuit locomoteur
spinal
Thèse
Louise Thiry
Sous la direction de :
iii
Résumé
La locomotion est contrôlée par des circuits spinaux qui génèrent le rythme locomoteur et coordonnent les activités musculaires entre la droite et la gauche du corps, et entre les muscles fléchisseurs et extenseurs. De plus, le rétrocontrôle sensoriel acheminé par les afférences proprioceptives et cutanées est crucial pour le fonctionnement normal du circuit locomoteur et pour l’adaptation du mouvement à l’environnement extérieur pendant la marche. La protéine DSCAM (Down Syndrom Cell Adhesion Molecule) est une molécule d’adhérence cellulaire impliquée dans un grand nombre de mécanismes nécessaires à la mise en place des réseaux neuronaux au cours du développement. Bien que DSCAM soit exprimée dans la moelle épinière de façon transitoire pendant le développement embryonnaire de la souris, et que sa mutation entraîne des défauts posturaux et moteurs très marqués, il y a très peu d’information sur son rôle dans le développement du circuit spinal lombaire contrôlant la locomotion. Dans ce contexte, les travaux présentés dans cet ouvrage visent à étudier l’implication de DSCAM dans la mise en place et le maintien du circuit locomoteur spinal. Pour cela, nous avons d’abord évalué les changements neurologiques du circuit spinal lombaire de souris néonatales et adultes mutantes pour DSCAM. Les souris portant la mutation systémique DSCAM2J présentent
des problèmes de coordination locomotrice associés à des changements anatomiques et neurophysiologiques dans le circuit interneuronal spinal.
Par ailleurs, nous avons étudié l’impact de la mutation DSCAM2J sur la coordination des membres pendant la
locomotion de la souris adulte. Nous montrons que la mutation de DSCAM altère la capacité des souris à courir et induit des changements dans leur répertoire locomoteur. En particulier, les souris mutantes DSCAM2J
présentent des patrons de marche auparavant jamais décrits pour la souris. De tels changements suggèrent une réorganisation des réseaux neuronaux spinaux et supraspinaux impliqués dans le contrôle locomoteur des souris mutantes DSCAM2J.
À l’aide de la technologie Cre-Lox, nous avons alors identifié et caractérisé la contribution de différentes populations interneuronales du circuit locomoteur spinal affectées par la mutation de DSCAM. Nous montrons dans cette étude que la mutation conditionnelle de DSCAM dans les interneurones spinaux excitateurs ou inhibiteurs entraîne un déséquilibre entre excitation et inhibition, induisant des défauts de coordination gauche/droite ou fléchisseur/extenseur, respectivement. Par immunomarquage, nous avons alors génétiquement identifié les populations interneuronales spinales impliquées dans le contrôle de la coordination gauche/droite ou fléchisseur/extenseur nécessaires à la locomotion.
L’ensemble des résultats de nos études fonctionnelles, électrophysiologiques et anatomiques suggère que la protéine DSCAM est nécessaire au développement du circuit locomoteur spinal des mammifères. En plus de
iv
caractériser les différents rôles de DSCAM dans la mise en place de ce réseau spinal, ce travail montre comment le développement de mutations conditionnelles de DSCAM dans différentes sous-populations neuronales permet d’étudier différentes composantes du circuit locomoteur spinal.
v
Abstract
Locomotion is controlled by spinal circuits that generate rhythm and coordinate left-right and flexor-extensor motoneuronal activities. The outputs of motoneurons and spinal interneuronal circuits are shaped by sensory feedback, relaying peripheral signals that are critical to the locomotor and postural control.Several studies in invertebrates and vertebrates have argued that the Down Syndrome Cell Adhesion Molecule (DSCAM) would play an important role in the normal development of neural circuits. Although there is evidence that DSCAM is expressed in the developing mouse spinal cord, and that its mutation induces postural and motor defects in adult mice, little is known about its functional contribution to the spinal circuits underlying locomotion.In this context, the work presented in this thesis aims at studying the implication of DSCAM in the establishment of the spinal locomotor circuit.
For this purpose, we first sought to evaluate the neurological changes in the spinal locomotor circuit of neonatal and adult DSCAM mutant mice. We show that a systemic mutation of DSCAM (DSCAM2J) induces
locomotor coordination defects associated with anatomical and neurophysiological changes in spinal interneuronal and sensorimotor circuits.
We then investigated the functional contribution of DSCAM to locomotor gaits over a wide range of locomotor speeds using freely walking mice. We show that the DSCAM2J mutation impairs the ability of mice to run and
modifies their locomotor repertoire, inducing the emergence of aberrant gaits for mice. Such changes suggest a reorganization of spinal and supraspinal neuronal circuits underlying locomotor control in DSCAM2J mutant
mice.
Finally, we used the Cre-Lox technology to genetically identify and characterize the neuronal populations underlying these functional changes. We show in this study that conditional mutations of DSCAM in either excitatory or inhibitory spinal interneurons induce an imbalance in excitatory-inhibitory signaling across the spinal midline that can impair the spinal locomotor circuit controlling either the bilateral coordination or the flexor/extensor coordination, respectively. Combining these studies with immunostaining experiments, we identified spinal interneuronal subpopulations implicated in either the bilateral or the flexor/extensor coordination during locomotion.
Collectively, our functional, electrophysiological, and anatomical studies suggest that the mammalian DSCAM protein is involved in the normal development of the spinal locomotor circuit. In addition to characterizing the different implications of DSCAM in the development of this spinal circuit, this work shows how the use of conditional mutations of DSCAM in different neuronal subpopulations allows the study of the spinal locomotor circuit components.
vi
Table des matières
Résumé ... iii
Abstract ... v
Table des matières... vi
Liste des abréviations ... ix
Liste des tableaux ... x
Liste des figures ... xi
Remerciements ... xiii
Avant-propos ... xiv
Introduction générale ...1
1. La locomotion ...1
1.1 Le cycle de marche...1
1.2 Organisation centrale et périphérique du contrôle locomoteur ...4
1.2.1 Le contrôle supraspinal descendant : initiation et contrôle de la locomotion ...5
1.2.2 Les afférences périphériques: modifications de la locomotion ...6
1.2.3 La moelle épinière : intégration des informations supraspinales et périphériques ...8
1.3 Le rythme et le patron locomoteurs sont générés au niveau de la moelle épinière ...9
2. La coordination des membres pendant la locomotion ...11
2.1 Chaque membre est contrôlé par un circuit spinal distinct ...12
2.2 Le rôle des interneurones propriospinaux ...13
3. Les interneurones spinaux : éléments clés du circuit locomoteur spinal ...14
3.1 Les interneurones dI6 ...16
3.2 Les interneurones V0...17
3.3 Les interneurones V1...18
3.4 Les interneurones V2...19
3.5 Les interneurones V3...20
3.6 Les interneurones Hb9 ...21
4. Les voies de signalisation moléculaire impliquées dans le développement du circuit locomoteur spinal ...22
4.1 Les éphrines ...23
4.2 Nétrine/DCC ...23
vii
5. La protéine DSCAM (molécule d’adhésion cellulaire associée au syndrome de Down) ...25
5.1 Caractéristiques moléculaires de la protéine DSCAM ...25
5.2 Gain de fonction de DSCAM ...27
5.2.1 Surexpression de DSCAM dans le syndrome de Down ...27
5.2.2 Surexpression de DSCAM dans d’autres maladies neuronales ...28
5.3 Perte de fonction de DSCAM ...29
5.3.1 Rôles de DSCAM dans le développement des réseaux neuraux ...29
5.3.1.1 DSCAM et organisation des neurones en réseaux ...29
5.3.1.2 DSCAM et guidage axonal ...33
5.3.2 Conséquences fonctionnelles de la mutation de DSCAM ...34
Problématique et objectifs des travaux ...37
1. Évaluation du circuit moteur spinal des souris néonatales DSCAM2J ...37
2. Évaluation de la coordination des membres pendant la locomotion des souris adultes DSCAM2J .38 3. Contribution des différentes populations interneuronales spinales affectées par la mutation de DSCAM ...38
Résultats ...39
CHAPITRE 1 : DSCAM est impliquée dans la mise en place des circuits spinaux locomoteur et sensorimoteur ...39
1.1 Notes introductives et contributions ...39
1.2 Résumé ...39
1.3 Role of DSCAM in the development of the spinal locomotor and sensorimotor circuits ...41
1.3.1 Abstract ...41
1.3.2 Introduction ...42
1.3.3 Materials and Methods ...44
1.3.4 Results ...49
1.3.5 Discussion ...68
1.3.6 Conclusion ...71
CHAPITRE 2 : La perte de fonction de DSCAM induit des changements de patron locomoteur dépendants de l’âge et de la vitesse de marche ...73
2.1 Notes introductives et contributions ...73
2.2 Résumé ...73
2.3 Age- and speed-dependent modulations of locomotor gaits in DSCAM2J mutant mice ...74
2.3.1 Abstract ...75
viii
2.3.3 Materials and Methods ...78
2.3.4 Results ...80
2.3.5 Discussion ...95
2.3.6 Conclusion ...99
CHAPITRE 3 : La mutation conditionnelle de DSCAM dans les interneurones spinaux excitateurs ou inhibiteurs permet d’étudier spécifiquement les circuits spinaux impliqués dans la coordination bilatérale ou fléchisseur/extenseur ... 100
3.1 Notes introductives et contributions ... 100
3.2 Résumé ... 100
3.3 Genetic dissection of the spinal motor circuitry of DSCAM conditional KO mice ... 101
3.3.1 Abstract ... 102
3.3.2 Introduction ... 103
3.3.3 Materials and Methods ... 104
3.3.4 Results ...107
3.3.5 Discussion ... 121
3.3.6 Conclusion ... 126
Discussion générale ... 127
1. Résumé des études ... 127
2. DSCAM est nécessaire au développement du circuit locomoteur spinal ... 128
2.1 Mutation DSCAM et coordination gauche-droite... 128
2.2 Mutation DSCAM et rythme locomoteur ... 130
2.3 Mutation DSCAM et coordination fléchisseur-extenseur ... 130
3. DSCAM est nécessaire à la coordination entre les membres pendant la locomotion ... 131
3.1 La mutation de DSCAM affecte l'adaptation des types de marche aux changements de vitesse………...……….132
3.2 Futurs travaux ... 132
4. DSCAM est nécessaire au développement du circuit réflexe spinal ... 133
4.1 La mutation de DSCAM affecte la transmission sensori-motrice ... 133
4.2 Futurs travaux ... 133
Conclusions ... 135
ix
Liste des abréviations
DCC Deleted in Colorectal CarcinomaDSCAM molécule d’adhérence cellulaire associée au syndrome de Down DSCR région critique associée au syndrome de Down
CAM molécule d’adhérence cellulaire CIN interneurone commissural CPG générateur de patron central DAC cellule amacrine dopaminergique EMG électromyographie
ENG électroneurographie
FSCD faisceau spinocérébelleux dorsal FSCV faisceau spinocérébelleux ventral IaIN interneurone de type Ia
Ig-C2 domaine immunoglobuline de type C2
IgSF molécules d’adhésion de la superfamille des immunoglobulines
IN interneurone
INe interneurone excitateur INi interneurone inhibiteur IPL couche plexiforme interne LTD dépression à long terme LTP potentialisation à long terme
MLR région locomotrice mésencéphalique
MN motoneurone
MRGC cellule ganglionnaire rétinienne à mélanopsine NMDAR récepteur NMDA
RBC cellule bipolaire rétinienne RGC cellule ganglionnaire rétinienne siRNA petit ARN interférant
x
Liste des tableaux
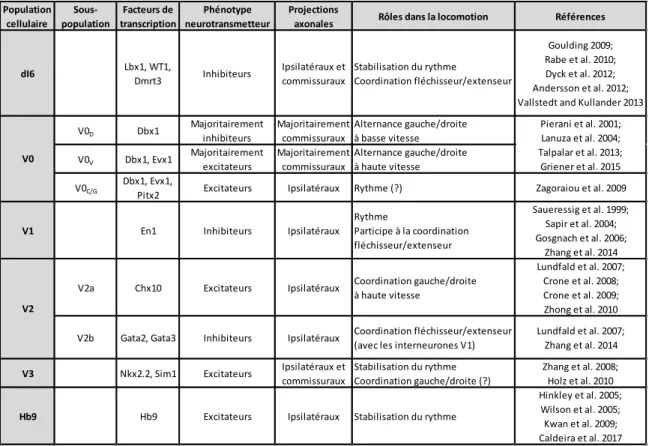
Tableau 1: Vision actuelle des populations interneuronales génétiquement identifiées et leur implication dans la locomotion... 22 Tableau 2: Les différents modèles murins de mutations de DSCAM étudiés à ce jour. ... 36
xi
Liste des figures
INTRODUCTION GÉNÉRALE
Figure 0.1: Les différentes phases du cycle de marche chez le chat ... 3
Figure 0.2: Le patron locomoteur dépend de la durée du cycle de marche, du ratio phase d’appui/phase de balancement et de la coordination entre les membres ... 4
Figure 0.3: Les préparations décérébrées : rôle des structures supraspinales dans l’initiation et le contrôle de la locomotion ... 6
Figure 0.4: Description anatomique de la moelle épinière ... 9
Figure 0.5: Capacité de la moelle épinière néonatale isolée à générer une activité rythmique locomotrice .... 11
Figure 0.6 : Détermination de l’identité neuronale des populations neuronales spinales ... 16
Figure 0.7 : Structure de la protéine DSCAM des vertébrés ... 26
Figure 0.8 : DSCAM et le principe d’auto-évitement des dendrites d’un même neurone ou de neurones voisins ... 31
CHAPITRE 1 Figure 1.1 : Properties of the locomotor activity in adult DSCAM2J and WT mice ... 50
Figure 1.2 : Pattern of locomotor-like activity in DSCAM2J and WT neonatal mice ... 52
Figure 1.3 : Bilateral and ipsilateral coordination of DSCAM2J and WT mice during neonatal fictive locomotion ... 53
Figure 1.4 : Number of commissural interneurons (CINs) in the L2 segment of WT and DSCAM2J neonatal spinal cords ... 54
Figure 1.5 : Fictive locomotion and spinal reflexes evoked upon dorsal root electrical stimulation in DSCAM2J and WT neonatal spinal cords ... 56
Figure 1.6 : Spinal reflexes evoked upon peripheral nerve stimulation in DSCAM2J and WT adult mice ... 58
Figure 1.7 : DSCAM expression in spinal motoneurons and dorsal root ganglions of neonatal and adult WT and DSCAM2J mice... 60
Figure 1.8 : Anatomical properties of lumbar L5 dorsal root ganglions in adult WT and DSCAM2J mice ... 61
Figure 1.9 : Anatomical properties of motoneuron pools in WT and DSCAM2J lumbar spinal cords ... 63
Figure 1.10 : Excitatory and inhibitory synaptic boutons on α-motoneurons in DSCAM2J and WT mice ... 66
CHAPITRE 2 Figure 2.1: Locomotor behaviour of C57BL/6J, C3H, C3H/HeDiSn and DSCAM2J mutant mice at different treadmill speeds ... 82
Figure 2.2: Gait occurrence for C57BL/6J, C3H, C3H/HeDiSn and DSCAM2J mice at different treadmill speeds at 3 and 8 weeks of age ... 84
Figure 2.3: Types of support at slow or fast locomotor speed ... 87
Figure 2.4: Distribution of weight on 2 supporting limbs at slow or fast locomotor speed ... 88
Figure 2.5: Transitions between gaits of C3H/HeDiSn and DSCAM2J 3-week-old mice at slow or fast locomotor speed ... 89
Figure 2.6: Stability and attractiveness of gaits for C57BL/6J, C3H, C3H/HeDiSn and DSCAM2J 3-week-old mice ... 91
xii
Figure 2.7: Probability of transitions between gaits for C3H/HeDiSn and DSCAM2J 3-week-old mice ... 93 CHAPITRE 3
Figure 3.1: Pattern of locomotor-like activity in vGluT2-DS and vGAT-DS neonatal spinal cords ... 109 Figure 3.2: Bilateral and ipsilateral coordination of vGluT2-DS and vGAT-DS mice during neonatal fictive locomotion ... 111 Figure 3.3: Number of excitatory and inhibitory intrasegmental commissural interneurons (CINs) in the L2 segment of vGluT2-DS and vGAT-DS neonatal spinal cords ... 114 Figure 3.4: Number of excitatory and inhibitory intersegmental commissural (CINs) and ipsilateral interneurons (ipsi-INs) in the L5 segment of vGluT2-DS and vGAT-DS neonatal spinal cords ... 116 Figure 3.5: Genetic identification of interneuronal subpopulations in vGluT2-DS and vGAT-DS embryonic spinal cords ... 118 Figure 3.6: Properties of the locomotor activity in adult vGluT2-DS and vGAT-DS mice ... 120 Figure 3.7: Modifications of the spinal locomotor circuit upon DSCAM conditional mutation in excitatory or inhibitory interneurons... 125
DISCUSSION GÉNÉRALE
xiii
Remerciements
Tout d’abord, j’aimerais remercier très sincèrement mon directeur de recherche, le Dr Frédéric Bretzner, pour son soutien, sa disponibilité et sa confiance, qui m’ont permis de développer un éventail de compétences techniques ainsi qu’un esprit critique, essentiels aux métiers de la recherche scientifique.
Je voudrais exprimer ici ma gratitude envers les professeurs Laurent Bouyer, Tuan Bui et Christian Éthier pour avoir accepté de participer à ce jury de thèse.
J’aimerais également remercier mes collègues de travail, pour leur aide, leurs conseils techniques et scientifiques, leur soutien moral et leur bonne humeur. Merci à Olivier D. Laflamme pour son accueil au Québec et pour sa gentillesse. Merci à Marie Roussel et Nicolas Josset, pour leurs conseils, leur générosité et leur amitié. C’était un réel plaisir de travailler avec vous. Je vous souhaite à tous les 3 une bonne réussite pour vos doctorats et vos projets futurs. Merci à Maxime Lemieux, pour ses précieux conseils, son esprit critique et sa disponibilité.
Je voudrais remercier tout spécialement Isabelle Bolduc pour son travail admirable à l’animalerie et sans qui j’aurais sans doute perdu tout espoir d’obtenir à temps le matériel nécessaire à la réalisation de mes expériences. Merci aussi aux étudiants et aux professionnels de recherche des autres équipes de l’Axe Neurosciences au bloc P du CHUL, ainsi qu’à tous les étudiants stagiaires qui ont participé à l’avancée des projets de notre laboratoire.
Je remercie également toutes les personnes que j’ai rencontrées au Québec, avec qui j’ai partagé de bons moments au laboratoire, mais également en dehors. Arriver seule dans un pays étranger n’est pas facile et vous m’avez aidé à trouver ma place. Merci à toi Anne, pour ton amitié inconditionnelle et ton soutien continu depuis toutes ces années. Merci également à tous nos amis rencontrés à Montréal, pour tous ces moments partagés à découvrir le Québec et ses merveilleux paysages. Merci à toi Nicolas, pour m’avoir fait redécouvrir ma passion pour l’escalade et pour m’aider tous les jours à avancer, à progresser et à ne pas oublier de respirer!
Enfin, j’aimerais remercier ma famille en France, qui m’a soutenue dans mes projets et qui a toujours été présente malgré la distance.
xiv
Avant-propos
La première partie de cette thèse est une revue de la littérature permettant d’introduire les principes essentiels à la compréhension des chapitres suivants. Les chapitres 1 à 3 de cette thèse sont présentés sous forme d’articles publiés, soumis ou en préparation pour publication. Ces articles sont issus de mes travaux de doctorat réalisés au cours de ces quatre dernières années dans le laboratoire du Dr Frédéric Bretzner. La dernière partie de cette thèse est consacrée à une discussion de l’ensemble des résultats présentés et des perspectives qui en découlent.
Les articles découlant de mes travaux et collaborations réalisés au cours de mon doctorat sont présentés ci-dessous. La numérotation des figures a été modifiée afin de faciliter leur intégration dans le manuscrit.
Articles inclus dans la thèse
CHAPITRE 1: DSCAM est impliquée dans la mise en place des circuits spinaux locomoteur et
sensorimoteur.
Ce premier chapitre vise à caractériser les défauts fonctionnels et anatomiques généraux du circuit spinal lombaire pouvant expliquer les défauts posturaux et locomoteurs observés en l’absence de la protéine DSCAM fonctionnelle. Pour cela, nous avons étudié la locomotion et les réflexes spinaux des souris mutantes DSCAM2J, à l’âge néonatal et à l’âge adulte. Nous avons complété ces analyses fonctionnelles par des études
de traçage et de marquage immunohistochimique afin de caractériser les changements anatomiques pouvant avoir des effets directs sur la fonctionnalité des circuits moteurs spinaux. Les résultats de ces travaux ont donné lieu à un article publié dans le Journal of Neurophysiology, présenté dans ce chapitre.
Role of DSCAM in the development of the spinal locomotor and sensorimotor circuits
Louise Thiry, Maxime Lemieux, Olivier D. Laflamme, Frédéric Bretzner. J Neurophysiol. 2016 Mar 1; 115(3):
1338–1354.
Ce projet a été conceptualisé et élaboré par moi-même sous la direction du Dr Bretzner. L’acquisition, l’analyse et l’interprétation de toutes les données électrophysiologiques (locomotion fictive et réflexes spinaux) et de toutes les données anatomiques (traçage rétrograde des interneurones commissuraux, immunohistochimie, imagerie et quantifications par microscopie à fluorescence) obtenues chez les souris néonatales ont été effectuées par moi-même sous la direction du Dr Bretzner. J’ai monté les figures présentant ces données néonatales. J’ai également effectué toute l’acquisition des données anatomiques sur les souris adultes (perfusion, dissection, congélation, coupe des tissus, et immunohistochimie), à l’exception du traçage rétrograde des motoneurones qui a été réalisés par O. D. Laflamme. L’imagerie et la quantification
xv
par microscopie à fluorescence, ainsi que le montage des figures présentant ces résultats ont été réalisées par moi-même. L’acquisition et l’analyse de la locomotion des souris adultes, ainsi que la figure présentant ces données ont été réalisées par le Dr Lemieux. L’acquisition et l’analyse des réflexes des souris adultes, ainsi que la figure présentant ces résultats ont été effectuées par O. D. Laflamme. J’ai rédigé l’article sous la direction du Dr Bretzner. Les Drs Tuan V. Bui, Izabela Panek, et Trevor Drew ont révisé la première version du manuscrit. Cet article a été publié dans le Journal of Neurophysiology en 2016.
CHAPITRE 2 : La perte de fonction de DSCAM induit des changements de patron locomoteur
dépendants de l’âge et de la vitesse de marche.
L’objectif de ce deuxième chapitre est d’étudier l’implication de DSCAM dans les circuits contrôlant la coordination entre les membres nécessaire à la locomotion. Pour cela, nous avons évalué l’impact de la mutation systémique DSCAM2J sur le répertoire locomoteur de la souris adulte. Afin d’identifier et de
caractériser les différents type de patrons de marche utilisés par les souris mutantes et contrôles, nous avons utilisé un protocole d’analyse précédemment mis en place et publié par notre laboratoire. Les résultats de ces travaux ont donné lieu à un article actuellement en révision, soumis dans le Journal of Neurophysiology, et présenté dans ce chapitre.
Speed and age-dependent modulation of locomotor gaits in DSCAM mutant mice
Louise Thiry, Maxime Lemieux, Frédéric Bretzner. Soumis dans le Journal of Neurophysiology.
Ce projet a été conceptualisé et élaboré par moi-même, avec l’aide du Dr Lemieux, sous la supervision du Dr Bretzner. J’ai effectué l’acquisition des données de cinématique avec l’aide de L-A. Porlier, stagiaire étudiant au Baccalauréat de Sciences biomédicales. J’ai effectué l’analyse des données de cinématique, le montage des figures et la rédaction d’une première version de l’article, avec l’aide du Dr Lemieux et sous la direction du Dr Bretzner. L’article, soumis dans le Journal of Neurophysiology et actuellement en révision, est présenté dans ce chapitre.
CHAPITRE 3 : La mutation conditionnelle de DSCAM dans les interneurones spinaux
excitateurs ou inhibiteurs permet d’étudier spécifiquement les circuits spinaux impliqués
dans la coordination bilatérale ou fléchisseur/extenseur.
Ce troisième chapitre a pour but d’identifier et de caractériser la contribution de différentes populations interneuronales du circuit locomoteur spinal affectées par la mutation de DSCAM. Pour cela, nous avons analysé la locomotion des souris portant la mutation conditionnelle de DSCAM dans les interneurones excitateurs ou inhibiteurs, à l’âge néonatal et à l’âge adulte. Nous avons complété ces analyses fonctionnelles
xvi
par des expériences de traçage et d’immunomarquage afin d’identifier génétiquement les différentes populations interneuronales spinales affectées par les mutations conditionnelles de DSCAM. Les résultats de ces travaux ont donné lieu à un article actuellement en préparation pour soumission dans le Journal of
Neurophysiology et présenté dans ce chapitre.
Genetic dissection of the spinal motor circuitry of DSCAM conditional KO mice Louise Thiry, Frédéric Bretzner. En préparation.
Ce projet a été conceptualisé et élaboré par moi-même sous la direction du Dr Bretzner. L’acquisition, l’analyse et l’interprétation de toutes les données électrophysiologiques (locomotion fictive néonatale) et de toutes les données anatomiques (traçage rétrograde des interneurones commissuraux, immunohistochimie, imagerie et quantifications par microscopie à fluorescence) obtenues chez les souris néonatales ont été effectuées par moi-même sous la direction du Dr Bretzner. J’ai également effectué l’acquisition, l’analyse et l’interprétation de toutes les données de cinématique obtenues durant la locomotion des souris adultes. J’ai réalisé les figures et rédigé une première version de l’article, sous la direction du Dr Bretzner. L’article est actuellement en préparation pour publication.
Articles publiés non inclus dans la thèse
Au cours de ce doctorat, j’ai également eu l’opportunité de travailler en collaboration avec d’autres équipes de recherche. Deux de ces collaborations ont déjà donné lieu à des publications :
- Motor hypertonia and lack of locomotor coordination in mutant mice lacking DSCAM.
Lemieux M, Laflamme OD, Thiry L, Boulanger-Piette A, Frenette J, Bretzner F. J Neurophysiol. 2016 Mar; 115(3):1355-71.
- Betacellulin regulates schwann cell proliferation and myelin formation in the injured mouse peripheral nerve.
Vallières N, Barrette B, Wang LX, Bélanger E, Thiry L, Schneider MR, Filali M, Côté D, Bretzner F, Lacroix S. Glia. 2017 Apr; 65(4):657-669.
1
Introduction générale
La locomotion est un comportement, simple en apparence, qui requiert l’activation coordonnée de nombreux muscles et d’un nombre encore plus important de cellules nerveuses. L’organisation précise de ces cellules en réseaux neuronaux fonctionnels est essentielle à la locomotion. De nombreuses études ont décrit l’implication de la protéine DSCAM dans le développement de certains réseaux neuronaux. Cependant, bien que la perte de fonction de cette protéine soit associée à de sévères défauts posturaux et moteurs, son implication dans le développement des circuits spinaux contrôlant la locomotion reste à ce jour inexplorée. Les travaux de recherche présentés dans ma thèse ont pour but d’étudier l’implication de la protéine DSCAM dans le développement et le maintien du circuit locomoteur spinal. Plus particulièrement, j’ai d’abord évalué les effets de la perte de fonction de la protéine DSCAM sur la fonctionnalité du circuit spinal lombaire néonatal et sur la locomotion des souris adultes. J’ai par la suite cherché à identifier et à caractériser la contribution de différentes populations interneuronales du circuit locomoteur spinal affectées par la perte de fonction de DSCAM.
Cette première partie de ma thèse vise à présenter l’état actuel des connaissances sur lesquelles se basent mes travaux de recherche.
1. La locomotion
La locomotion est la capacité de se mouvoir pour se déplacer d’un point donné à un autre. Cette faculté est essentielle à la vie animale et permet notamment de chercher de la nourriture, d’explorer, de fuir un danger, ou d’atteindre un endroit particulier. Selon l’espèce animale, différentes formes de locomotion ont évolué, comme la nage, le vol ou la marche. Ces comportements ont comme caractéristique commune la rythmicité et l’alternance des mouvements du corps ou des membres. Ce caractère rythmique donne à la locomotion l’apparence d’un comportement simple et stéréotypé, impliquant la répétition d’un même mouvement. Cependant, la locomotion se déroule généralement dans des environnements imprévisibles, comportant toutes sortes d’obstacles, et doit donc être constamment modifiée pour adapter les mouvements à l’environnement immédiat.
1.1 Le cycle de marche
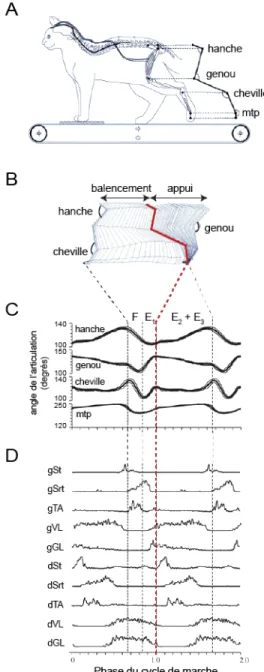
Chez les vertébrés, la locomotion est caractérisée par la succession de cycles de flexion et d’extension des membres permettant une alternance entre les phases de balancement et les phases d’appui, assurant ainsi la propulsion de l’animal (Phillipson, 1905, Engberg and Lundberg, 1969). La phase de balancement débute lorsque le membre quitte le sol et se termine lorsqu’il reprend appui sur le sol, alors que la phase d’appui
2
correspond au moment où le membre est posé au sol (Figure 0.1). Un cycle de marche correspond à la période entre le début de deux phases de balancement successives, et peut être divisé en quatre phases distinctes : flexion (F), première extension (E1), deuxième extension (E2) et troisième extension (E3). La phase de balancement commence par une flexion de la hanche, du genou et de la cheville (phase F). Environ à la moitié de la phase de balancement, la phase E1 débute par une extension du genou et de la cheville alors que la hanche est encore en flexion, permettant ainsi la propulsion du pied en avant du corps. Au début de la phase d’appui (phase E2), le poids du corps est transféré au membre qui se pose au sol. La fin de la phase d’appui (phase E3) est caractérisée par une rapide extension de toutes les articulations, permettant la propulsion du corps vers l’avant. Cette description simplifiée du cycle de marche est basée sur les mouvements d’un seul membre. En plus de la coordination entre flexion et extension des différents muscles d’un membre donné, la locomotion nécessite une coordination étroite entre les différents membres, faisant ainsi intervenir un nombre important de muscles synergistes et antagonistes (Figure 0.1D).
3
Figure 0.1: Les différentes phases du cycle de marche chez le chat. (A) Représentation schématique du modèle expérimental., (B) Diagramme en bâtons représentant les variations des angles articulaires de la hanche, du genou, de la cheville et de l’articulation métatarsophalangienne (mtp) au cours d’une phase de balancement et d’une phase d’appui. (C) Moyenne des variations angulaires observées sur les articulations du membre postérieurs au cours de 10 cycles locomoteurs. Les différentes phases du cycle locomoteur sont représentées par des lignes pointillées : phase de balancement (F et E1) et phase d’appui (E2 et E3). (D) Moyenne des tracés électromyographiques (EMGs) rectifiés
enregistrés dans différents muscles au cours de 10 cycles locomoteurs. g=gauche et d=droit. Les différents muscles enregistrés sont : le Semitendinosus (St, fléchisseur du genou et extenseur de la hanche), le Sartorius (Srt, fléchisseur de la hanche et extenseur du genou), le Tibialis Anterior (TA, fléchisseur de la hanche), le Vastus Lateralis (VL, extenseur du genou), le Gastrocnemius Lateralis (GL, extenseur de la cheville). (Adapté de Rossignol and Bouyer 2004).
4
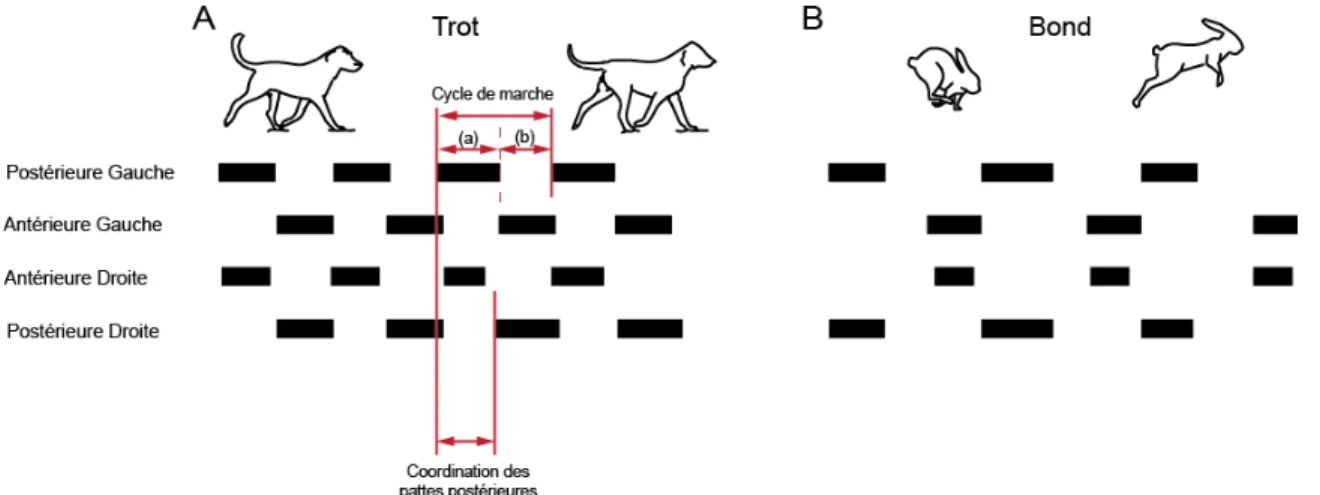
Cette séquence complexe de contractions des muscles résulte de deux propriétés fondamentales, soit le rythme et le patron locomoteur. Le rythme locomoteur correspond à la fréquence du cycle de marche (Grillner, 1975, Pearson, 1976), alors que le patron locomoteur correspond au type de locomotion employé pour se déplacer, comme la marche, la course ou encore le trot et le galop pour les animaux quadrupèdes. Le patron locomoteur dépend de plusieurs facteurs, tels que la durée du cycle de marche, le ratio phase d’appui/phase de balancement ainsi que la coordination entre les membres (Hildebrand, 1976) (Figure 0.2).
Figure 0.2: Le patron locomoteur dépend de la durée du cycle de marche, du ratio phase d’appui/phase de balancement et de la coordination entre les membres. Représentation du trot (A) et du bond (B). Pour chaque membre, chaque trait noir représente une phase d’appui (a) alors que les espaces entre ces traits représentent les phases de balancement (b). Le cycle de marche, ainsi que le couplage entre les membres postérieurs sont représentés par des flèches rouges. (Adapté de Abourachid et al., 2007).
Ainsi, la locomotion nécessite des ajustements posturaux continuels impliquant la coordination précise de l’activité des muscles du tronc et des membres (Gramsbergen, 1998, Ivanenko et al., 2004, Ceccato et al., 2009) et résulte des interactions complexes entre les contraintes biomécaniques des muscles de chaque membres, attachés au squelette, et les circuits neuraux contrôlant ces muscles (Schamhardt, 1998, Biewener, 2006, Maes and Abourachid, 2013). Cette thèse s’attache principalement à l’étude du contrôle locomoteur par le système nerveux, qui soulève deux questions fondamentales. D’une part, comment les réseaux de cellules nerveuses génèrent-ils le rythme locomoteur? D’autre part, comment les patrons locomoteurs sont-ils modifiés pour adapter la locomotion aux perturbations de l’environnement immédiat?
1.2 Organisation centrale et périphérique du contrôle locomoteur
Chez les vertébrés, les structures neurales impliquées dans l’initiation et le contrôle de la locomotion sont relativement bien conservées. Les structures centrales supraspinales initient et contrôlent la locomotion,
5
alors que les structures périphériques contribuent à adapter le mouvement aux contraintes environnementales immédiates. Ces informations sont intégrées au niveau de la moelle épinière qui produit la commande motrice.
1.2.1 Le contrôle supraspinal descendant : initiation et contrôle de la locomotion
L’initiation et la programmation des actes locomoteurs font intervenir plusieurs aires corticales, telles que le cortex moteur primaire, le cortex prémoteur et l’aire motrice supplémentaire. Les informations corticales sont transmises par deux voies de projections descendantes : la voie pyramidale et la voie extrapyramidale. Des études de lésion ont montré l’importance de ces voies descendantes dans le contrôle de la locomotion (Jiang and Drew, 1996, Brustein and Rossignol, 1998). La voie pyramidale est impliquée dans le contrôle des mouvements volontaires distaux fins. La majorité des axones de cette voie projette directement sur les neurones spinaux après avoir décussé au niveau bulbaire. Cependant, une partie des axones forme des collatérales qui projettent au niveau des noyaux moteurs du diencéphale et du tronc cérébral. La voie extrapyramidale inclue les voies rubrospinale, réticulospinale et vestibulospinale. Elle forme un relais au niveau des ganglions de la base avant de projeter sur les centres initiateurs de la locomotion : les régions locomotrices sous-thalamique (Région Locomotrice Sous-thalamique; SLR) et mésencéphalique (Région Locomotrice Mésencéphalique; MLR). La voie extrapyramidale forme ensuite un relais dans la formation réticulée avant de projeter vers la moelle épinière via les funiculi ventro-latéraux et dorso-latéraux.
Un premier aperçu des mécanismes neuronaux du contrôle de la locomotion a été révélé il y a maintenant près d’un siècle, lorsqu’il a été montré que l’ablation des hémisphères cérébraux des chiens et des chats n’abolie pas la marche (Sherrington, 1906, Brown, 1911, Brown, 1914, Miller et al., 1975, Bjursten et al., 1976, MacKay-Lyons, 2002). Ces préparations d’animaux décérébrés correspondent à une section transversale complète du tronc cérébral au niveau du mésencéphale, empêchant ainsi les régions cérébrales plus rostrales telles que le cortex d’influencer le patron locomoteur. Le fait que les animaux décérébrés soient capables de marcher de façon spontanée suggère donc que le cortex cérébral n’est pas indispensable à l’initiation de la locomotion.
Depuis, les préparations d’animaux décérébrés à différents niveaux ont permis d’étudier le rôle des différentes structures supraspinales dans l’initiation et le contrôle de la locomotion (Figure 0.3). Ainsi, lorsque la section est réalisée en avant des colliculi supérieurs (préparation prémammilaire), préservant la SLR, la locomotion spontanée est préservée, bien que fortement stéréotypée et inadaptée aux contraintes environnementales (Mori et al., 1989, Whelan, 1996). Enfin, une section transversale plus caudale, supprimant la SLR (préparation postmammilaire), abolie la locomotion spontanée. Dans ce cas, la
6
stimulation électrique de la MLR permet d’initier différents patrons locomoteurs (de la marche au galop) en fonction de l’intensité de stimulation (Shik et al., 1966).
Dans les deux types de préparations décérébrées (prémammilaire et postmammilaire), la locomotion peut être observée si les pattes de l’animal sont posées sur un tapis roulant motorisé (Shik et al., 1966, Whelan, 1996). Ceci suggère un rôle des structures supraspinales dans le maintien du tonus postural, indispensable à l’équilibre pendant la locomotion. Ce maintien est assuré par un ajustement constant de la contractilité des muscles fléchisseurs et extenseurs. Par exemple, la suppression de l’influence des noyaux rouges par section transversale intercolliculaire induit une rigidité dite de décérébration (hyperextension) (Sherrington, 1906). Ainsi, les noyaux rouges et la formation réticulée pontique interviennent majoritairement dans la régulation de la phase de balancement, en projetant sur les interneurones et les motoneurones fléchisseurs de la moelle épinière (Drew and Rossignol, 1984, Lai and Siegel, 1990, Perreault et al., 1993, Perreault et al., 1994, Rho et al., 1999, Rossignol et al., 2006). À l’inverse, les noyaux vestibulaires de Deiters, qui sont activés phasiquement au cours de la locomotion, projettent sur les neurones extenseurs et augmentent l’activité des muscles antigravitaires lors de la phase d’appui (Orlovsky, 1972).
Figure 0.3: Les préparations décérébrées : rôle des structures supraspinales dans l’initiation et le contrôle de la locomotion.Selon le niveau de section du tronc cérébral, la locomotion est spontanée (section A, réalisée en avant des colliculi supérieurs (préparation prémammilaire)), ou nécessite la stimulation électrique de la région locomotrice mésencéphalique (section B, préparation postmammilaire). Thal=Thalamus, CS=Colliculi Supérieurs, CI=Colliculi Inférieurs, CM=Corps Mammilaire, MLR=Région Locomotrice Mésencéphalique (en rouge). (Adapté de Pearson and Gordon 2000).
1.2.2 Les afférences périphériques: modifications de la locomotion
Durant la réalisation d’un mouvement, les afférences sensorielles proprioceptives et cutanées renseignent le système nerveux sur l’exécution de la commande motrice. Ces informations proviennent de récepteurs sensoriels localisés dans les muscles, les tendons, les articulations et la peau, (Dietz, 2002b). En raison de leurs différentes natures, ces récepteurs répondent à différents stimuli, et permettent une adaptation précise
7
et rapide de la locomotion aux contraintes internes de l’organisme et aux exigences externes de l’environnement. Deux types de préparations expérimentales ont permis d’étudier le rôle des afférences périphériques pendant la locomotion : les dénervations et les préparations paralysés. Dans les modèles de dénervations, la dénervation, réalisée par section de toutes les racines spinales dorsales innervant les membres, permet d’étudier la locomotion en l’absence de toute information sensorielle provenant des membres. L’utilisation d’animaux dits immobilisés, ou paralysés, consiste à empêcher l’exécution du mouvement, permettant ainsi l’étude de l’activité motoneuronale en l’absence de tout réflexe sensoriel. La paralysie est souvent réalisée par injection de d-tubocurare, un inhibiteur de l’acétylcholine qui bloque la transmission synaptique à la jonction neuromusculaire, immobilisant ainsi les muscles. Lorsqu’un épisode de locomotion est évoqué dans une préparation d’animal paralysé, la locomotion est dite fictive car les motoneurones associés aux muscles fléchisseurs et extenseurs déchargent en alternance sans qu’aucun réel mouvement ne soit observé. Les enregistrements intra- et extracellulaires des neurones spinaux des animaux décérébrés et paralysés permettent ainsi d’étudier les évènements synaptiques associés au comportement locomoteur ainsi que l’organisation des différentes voies réflexes impliquées dans la locomotion.
Ainsi, les afférences cutanées provenant des mécanorécepteurs (complexe de Merkel et corpuscules de Ruffini et de Meissner) codent les informations relatives au toucher discriminatif (fibres des groupes II/III). D’autres afférences cutanées, provenant des corpuscules de Pacini et des terminaisons libres, codent respectivement les dépressions phasiques de la surface cutanée et les sensations douloureuses (fibres des groupes III/IV), et sont à l’origine d’un réflexe de défense connu sous le terme de reflexe ipsilatéral de flexion. La stimulation électrique des afférences cutanées augmente l’amplitude des bouffées d’activité enregistrées lors d’un épisode de locomotion fictive chez le chat décérébré et paralysé (Viala and Buser, 1969, Viala and Vidal, 1978, Fleshman et al., 1984), ainsi que chez le chat éveillé (Bretzner and Drew, 2005b). Ces afférences cutanées semblent également être impliquées dans la précision du placement des membres au cours de la locomotion puisque les chats spinaux dénervés sont incapables de réaliser correctement une marche sur les barreaux d’une échelle (Bouyer and Rossignol, 2003).
Les afférences proprioceptives sont organisées en plusieurs classes : les afférences en provenance des fuseaux neuromusculaires (fibres Ia et II), des organes tendineux de Golgi (fibre Ib) et des mécanorécepteurs de la capsule articulaire (fibres I/II). Ces afférences sont notamment impliquées dans les réflexes myotatique (fibre Ia) et myotatique inverse (fibre Ib) et jouent aussi un rôle essentiel dans la modulation de la locomotion. En effet, lors de la locomotion sur tapis roulant d’un animal décérébré, la fréquence locomotrice est corrélée à la vitesse du tapis roulant, suggérant que les informations sensorielles périphériques provenant des membres régulent le cycle de marche (Halbertsma, 1983, Frigon and Gossard,
8
2009). Il est intéressant de noter que lorsque la fréquence locomotrice augmente, la durée de la phase d’appui diminue alors que la durée de la phase de balancement reste constante. Cette observation suggère que les afférences périphériques régulent la durée de la phase d’appui, grâce à des informations sensorielles qui signaleraient la fin de la phase d’appui et initieraient la phase de balancement. Les afférences proprioceptives sont également capables d’initier des séquences locomotrices en activant les réseaux locomoteurs spinaux. Chez le chat, la stimulation des afférences proprioceptives par des mouvements passifs des articulations déclenche un épisode de locomotion (Sherrington, 1910, Grillner and Rossignol, 1978). Un tel entrainement locomoteur peut également être observé lors de la stimulation électrique des afférences périphériques sur une préparation de moelle épinière isolée de rat (Smith et al., 1988, Marchetti et al., 2001, Juvin et al., 2012) et de souris (Whelan et al., 2000) nouveau-nés. Plus récemment, des études d’ablation ou d’inhibition des afférences proprioceptives au moyen de mutations génétiques ont montré le rôle essentiel de ces retours sensoriels dans la production de mouvements locomoteurs coordonnés (Akay et al., 2014, Takeoka et al., 2014).
1.2.3 La moelle épinière : intégration des informations supraspinales et périphériques Une fois générée, la commande centrale supraspinale est adressée aux régions médullaires. À ce niveau, des réseaux de neurones sont capables de transformer la commande tonique descendante en une activité rythmique qu’ils transmettent aux motoneurones. Chez les mammifères, ces réseaux neuronaux sont localisés au sein de la moelle épinière lombaire et cervicale (Figure 0.4). La moelle épinière, également appelée moelle spinale, est aussi la principale porte d’entrée des fibres nerveuses qui apportent les informations sensorielles périphériques, ainsi que la porte de sortie des fibres motrices. La moelle épinière est composée de la matière grise qui regroupe les corps cellulaires des neurones, et de la matière blanche qui est constituée de leurs axones. La matière blanche est le lieu de passage des faisceaux sensoriels ascendants et des faisceaux moteurs descendants des structures supraspinales. Les corps cellulaires des neurones sensoriels sont regroupés dans les ganglions spinaux, et leurs axones forment les racines dorsales et projettent dans la portion dorsale de la moelle épinière. À ce niveau, la matière grise forme les cornes dorsales, qui contiennent principalement des interneurones qui reçoivent et transmettent l’information sensorielle. Les cornes ventrales contiennent des interneurones, ainsi que les corps cellulaires des motoneurones, dont les axones forment les racines ventrales avant d’aller innerver les muscles. En se basant sur les caractéristiques cellulaires des différents neurones observés, Bror Rexed (1914-2002) propose une subdivision de la matière grise en 10 couches, appelées lames de Rexed, de la couche superficielle de la corne dorsale (couche I) jusqu’à la couche X, entourant le canal central.
9
Figure 0.4: Description anatomique de la moelle épinière.(A) Représentation schématique du cerveau et de la moelle épinière, avec les renflements cervical et lombaire indiqués en rouge. (B et C) Représentations schématiques de coupes transversales de la moelle épinière, avec l’entrée des fibres sensorielles et la sortie des fibres motrices (B), ainsi que les dix lames de Rexed (C).
1.3 Le rythme et le patron locomoteurs sont générés au niveau de la moelle épinière
Bien que les structures supraspinales soient nécessaires à l’initiation de la locomotion et que les afférences périphériques soient essentielles à sa modulation, les réseaux neuronaux contenus dans la moelle épinière sont nécessaires et suffisants pour générer le rythme et le patron locomoteurs. En effet, l’utilisation de préparations isolées ou semi-intactes a permis de montrer que des contractions rythmiques et alternées des muscles des pattes postérieures pouvaient être évoquées même après suppression complète des informations sensorielles provenant des membres (Brown, 1911). De plus, bien que la section complète de la moelle épinière au niveau thoracique (préparation spinales) entraine une paralysie des membres postérieurs, cet effet n’est pas permanent (Phillipson, 1905, Brown, 1911, Brown, 1914, Jankowska et al., 1967a, b, Viala and Buser, 1971, Grillner and Zangger, 1979, Forssberg et al., 1980). En particulier, les jeunes chats spinaux sont capables de récupérer spontanément la marche en quelques semaines et de retrouver une activité rythmique et coordonnée des membres postérieurs (Forssberg et al., 1980). La récupération de la locomotion des chats adultes peut être accélérée par un entrainement quotidien sur tapis roulant, associé à une stimulation cutanée de la région périnéale (Barbeau and Rossignol, 1987, Belanger et al., 1996, De Leon et al., 1998a, b). L’utilisation de ces préparations spinalisées permet d’isoler le segment spinal contrôlant la musculature des membres postérieurs du reste du système nerveux central, et d’étudier ainsi le rôle des réseaux spinaux dans la génération du rythme et du patron locomoteur.
À la suite de ses travaux (1911), Graham Brown a suggéré l’existence de réseaux neuronaux, contenus dans la moelle épinière, capables de générer un patron d’activité locomoteur en l’absence de commande descendante provenant des centres supérieurs. Il a alors proposé un modèle, revisité par Anders Lundberg
10
au cours des années 1960, dans lequel l’alternance fléchisseur/extenseur observée durant la marche est contrôlée par deux réseaux de neurones, appelés demi-centres, qui s’inhibent mutuellement. Dans ce modèle, chaque membre est contrôlé par un circuit séparé qui contrôle l’activité des motoneurones fléchisseurs et extenseurs via des groupes d’interneurones excitateurs. Par l’intermédiaire d’interneurones commissuraux, des connexions inhibitrices entre ces demi-centres permettent à l’un d’être actif, pendant que l’autre reste silencieux. Selon ce modèle, les informations sensorielles provenant des membres seraient responsables de la complexité du patron locomoteur. Cependant, à la suite de ses travaux montrant que le patron locomoteur est conservé même après section de toutes les racines dorsales chez le chat spinalisé, Sten Grillner a proposé en 1981 le concept des générateurs de patron centraux (en anglais central pattern generators, CPGs), faisant référence aux réseaux neuronaux spinaux capables de générer un patron d’activité locomoteur en l’absence de retour sensoriel et de commande descendante provenant des centres supérieurs (Grillner, 1981).
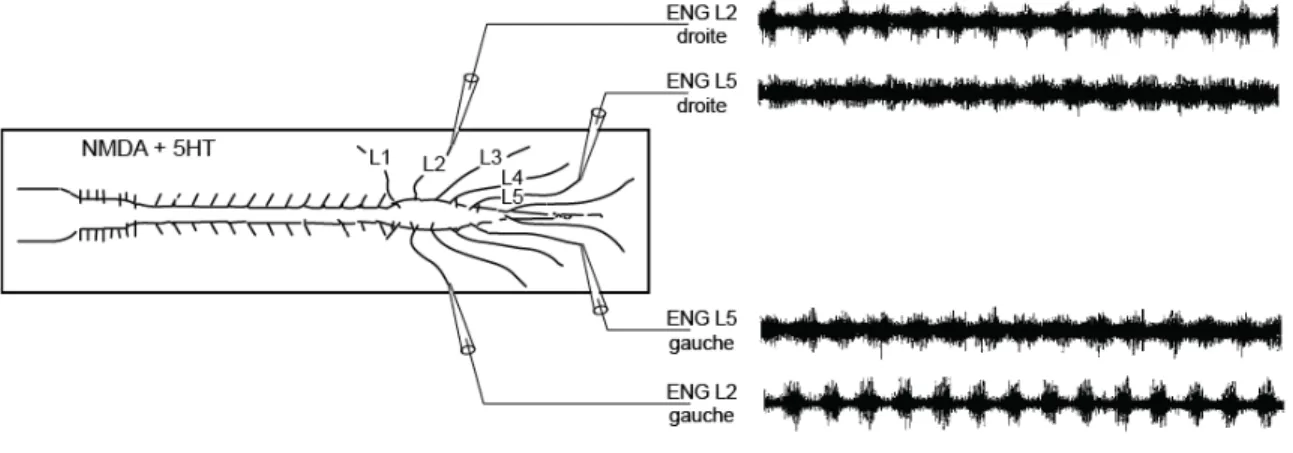
Depuis, il a été découvert qu’une activité rythmique et coordonnée peut être enregistrée à partir des racines ventrales de la moelle épinière de rat ou de souris nouveau-né (0 – 5 jours après la naissance) lorsqu’elle est isolée du corps de l’animal et placée dans une solution de liquide céphalorachidien artificielle contenant les neurotransmetteurs mimant les commandes supraspinales descendantes (NMDA, dopamine et sérotonine) (Cazalets et al., 1992, Cazalets et al., 1995, Kiehn and Kjaerulff, 1996, Cowley and Schmidt, 1997, Whelan et al., 2000) (Figure 0.5). Cette activité électroneurographique (ENG), enregistrée à partir des nerfs, est appelée locomotion fictive car elle ne conduit pas à de réels mouvements puisque la moelle épinière est isolée du corps de l’animal.
La préparation de moelle épinière isolée associée à des techniques d’électrophysiologie, de traçage anatomique et de génétique moléculaire a ainsi permis d’étudier les circuits spinaux qui sous-tendent le comportement locomoteur.
11
Figure 0.5: Capacité de la moelle épinière néonatale isolée à générer une activité rythmique locomotrice. Enregistrements électroneurographiques (ENGs) de l’activité des racines ventrales lombaires L2 et L5 droites et gauches de la moelle épinière isolée d’une souris néonatale (P3). La moelle épinière isolée est placée dans un bain de liquide céphalorachidien artificiel et l’activité rythmique, appelée locomotion fictive, est induite par application de sérotonine (10-4 M) et NMDA (2x10-5 M) dans le bain. (Adapté de Cazalets et al., 1995).
2. La coordination des membres pendant la locomotion
L’efficacité de la locomotion des espèces quadrupèdes et bipèdes, dont l’être humain, dépend de la coordination des membres ainsi que de la stabilisation des muscles du tronc (Wannier et al., 2001, Dietz, 2002a, Dominici et al., 2011). Pour la plupart des espèces animales quadrupèdes, dont les rongeurs, les membres antérieurs et postérieurs présentent des mouvements alternés entre la gauche et la droite pendant la locomotion à basses et à moyennes vitesses, qui deviennent plus synchronisés à hautes vitesses (English and Lennard, 1982, Lemieux et al., 2016a). De plus, la coordination diagonale des mouvements des membres antérieurs et postérieurs varie selon les patrons locomoteurs (Miller et al., 1975, English and Lennard, 1982, Lemieux et al., 2016a).
De manière intéressante, différents types de rythmes peuvent être induits selon la combinaison de neurotransmetteurs ajoutée à la solution saline dans laquelle est maintenue la moelle épinière isolée (Cowley and Schmidt, 1994, Kiehn and Kjaerulff, 1996, Cowley and Schmidt, 1997). Par ailleurs, des comportements moteurs rythmiques différents de la locomotion, tels que le comportement de grattement, peuvent être induits et enregistrés dans les préparations décérébrées ou spinalisées (Arshavsky et al., 1978, Perreault et al., 1999, Frigon and Gossard, 2010, Power et al., 2010), suggérant l’existence de différents circuits spinaux spécialisés (pour revue, Frigon 2012).
12
2.1 Chaque membre est contrôlé par un circuit spinal distinct
L’enregistrement simultané des nerfs moteurs reliés aux membres antérieurs et postérieurs dans les préparations décérébrées, ou des racines ventrales cervicales (innervant les membres antérieurs (McKenna et al., 2000)) et lombaires (innervant les membres postérieurs (Kjaerulff and Kiehn, 1996)) dans les préparations de moelles épinières isolées, montre un rythme de fréquence similaire et coordonné (Viala and Vidal, 1978, Orsal et al., 1990, Ballion et al., 2001, Juvin et al., 2005, Juvin et al., 2012), indiquant une forte interaction entre les réseaux cervicaux et lombaires. Cependant, après section transversale complète de la moelle épinière thoracique, séparant les segments spinaux cervical et lombaire, les activités ENGs cervicales et lombaires sont toujours actives mais ne sont plus coordonnées (Ballion et al., 2001, Juvin et al., 2005), et, selon les préparations expérimentales, le rythme des activités cervicales peut être plus rapide (Juvin et al., 2005) ou plus lent (Ballion et al., 2001, Juvin et al., 2005) que celui des activités lombaires. L’ensemble de ces données indique qu’il existe des réseaux distincts cervical (Ballion et al., 2001, Yamaguchi, 2004) et lombaire (Cazalets et al., 1995, Marcoux and Rossignol, 2000, Langlet et al., 2005), contrôlant l’activité rythmique des membres antérieurs et postérieurs, respectivement.
Par ailleurs, après section spinale réalisée le long de la ligne médiane pour séparer les côtés droit et gauche, les racines ventrales de chaque côté de la moelle épinière continuent de générer une activité rythmique (Kudo and Yamada, 1987, Kato, 1988). De manière similaire, chez le chat dénervé et immobilisé, l’utilisation d’un tapis roulant conçu pour contraindre les membres postérieurs droit et gauche à marcher à différentes vitesses, la fréquence de pas de chaque patte s’adapte indépendamment de l’autre à la vitesse imposée (Forssberg et al., 1980, Frigon et al., 2013), renforçant l’idée que les mouvements locomoteurs des différents membres sont générés par des circuits spinaux distincts.
Si chaque membre est contrôlé par son propre circuit locomoteur spinal, comment ces réseaux communiquent-ils entre eux pour assurer une locomotion à la fois stable et assez flexible pour s’ajuster aux contraintes de l’environnement dans lequel l’animal évolue? De nombreuses études de lésions de la moelle épinière ont permis d’identifier quelles parties du système nerveux participent à la coordination des membres pendant la locomotion. Cependant, avant de discuter du rôle des voies neuronales contenues dans la moelle épinière, il est important de noter que les contraintes biomécaniques des muscles de chaque membres, attachés au squelette, et dont les structures supraspinales sont informées via le rétrocontrôle sensoriel provenant de la périphérie, jouent un rôle important dans la stabilisation des membres pendant la locomotion (Schamhardt, 1998, Biewener, 2006, Maes and Abourachid, 2013).
13
2.2 Le rôle des interneurones propriospinaux
En plus des structures supraspinales et des contraintes biomécaniques reliant les membres, des réseaux de neurones, contenus dans la moelle épinière, participent au contrôle de la coordination des membres pendant la locomotion. Ces interneurones propriospinaux connectent entre eux les différents segments spinaux via des faisceaux ascendants et descendants pouvant projeter sur de courtes (un à six segments) ou de longues (plus de six segments) distances (Skinner et al., 1979, Flynn et al., 2011, Saywell et al., 2011, Ruder et al., 2016). Les faisceaux propriospinaux ascendants et descendants relient les segments cervicaux et lombaires via des projections axonales ipsilatérales et commissurales, soit directement sur les motoneurones, soit sur d’autres interneurones spinaux (Jankowska et al., 1973, Jankowska et al., 1974, Reed et al., 2006, Brockett et al., 2013, Ruder et al., 2016). D’après des études de traçage anatomique (Ruder et al., 2016) et d’enregistrements électrophysiologiques (Alstermark et al., 1987), ces interneurones propriospinaux reçoivent également des projections synaptiques supraspinales.
De nombreuses études de lésions sur des préparations décérébrées, ainsi que sur des préparations de moelles épinières néonatales semi-isolées ou isolées, ont permis de démontrer l’implication de ces interneurones propriospinaux dans le contrôle de la coordination des membres pendant la locomotion. Par exemple, alors qu’une section longitudinale séparant les côtés gauche et droit de la moelle épinière entière inhibe la coordination bilatérale des activités ENG pendant la locomotion fictive, celle-ci n’est pas affectée si la section longitudinale ne touche que les segments sacral et lombaire caudal (jusqu’au niveau L2/L3) (Cazalets et al., 1995). De la même manière, une lésion longitudinale chronique, séparant les moitiés droite et gauche de la moelle épinière entre L1 et S1 n’affecte pas la coordination bilatérale des membres postérieurs du chat chronique (Kato, 1988). La coordination gauche/droite des racines ventrales lombaires est aussi préservée lorsque les segments spinaux thoraciques et cervicaux sont séparés en deux le long de la ligne médiane (Cowley and Schmidt 1997). Ces données indiquent qu’il n’existe pas une région donnée unique de la moelle épinière essentielle pour la coordination bilatérale des membres postérieurs. Au contraire, il semblerait plutôt que les interactions entre la gauche et la droite soient contrôlées par un réseau de projections axonales croisées, distribuées tout le long de la moelle épinière, du segment cervical au segment lombaire et réciproquement (Cowley and Schmidt, 1997, Cowley et al., 2009, Cowley et al., 2010, Juvin et al., 2012). De plus, la coordination des activités ENGs cervicales et lombaires est fortement affectée par l’inhibition ciblée de la transmission synaptique de tous les segments spinaux thoraciques grâce à l’utilisation de bains cloisonnés, permettant de soumettre les différents segments de la moelle épinière isolée à différentes drogues (Juvin et al., 2012). Ces résultats indiquent que les projections directes reliant les segments cervicaux et lombaires via le funicule ventral (Reed et al., 2006, Reed et al., 2009) ne sont pas à elles seules suffisantes pour contrôler la coordination entre les membres antérieurs et
14
postérieurs. De manière similaire, l’inhibition de la transmission synaptique des segments spinaux cervicaux et thoraciques bloque l’activité de locomotion fictive enregistrée au niveau lombaire à la suite de la stimulation électrique ou pharmacologique du tronc cérébral (Zaporozhets et al., 2004, 2006), indiquant le rôle essentiel des relais synaptiques propriospinaux dans la transmission des informations descendants du tronc cérébral (Cowley et al., 2010). Enfin, l’inhibition ciblée des neurones propriospinaux descendants induit des changements dans le comportement locomoteur des souris adultes, incluant des défauts de stabilité posturale, ainsi qu’une diminution de la distance maximale parcourue et de la vitesse maximale atteinte (Ruder et al., 2016). Par ailleurs, en l’absence de ces neurones, les souris contraintes à marcher à haute vitesse de locomotion (40 cm/s) au moyen d’un tapis roulant présentent des défauts de coordination bilatérale des membres postérieurs, caractérisés par une augmentation de la synchronisation gauche-droite, suggérant un rôle des neurones propriospinaux dans le contrôle de l’adaptation de la coordination bilatérale des membres aux changements de vitesses.
Ainsi, la préparation de moelle épinière isolée associée à des techniques d’électrophysiologie et de traçage anatomique a permis d’identifier un réseau de neurones impliqué dans le contrôle de la coordination des membres pendant la locomotion. Par ailleurs, ce type de préparation a également permis d’étudier le rôle de populations spécifiques d’interneurones spinaux dans la génération du rythme et du patron locomoteurs chez le mammifère.
3. Les interneurones spinaux : éléments clés du circuit locomoteur spinal
L’activité locomotrice serait initiée à la suite de l’activation d’interneurones spinaux excitateurs formant des projections axonales ipsilatérales. En effet, une activité rythmique peut être engendrée à la suite de l’augmentation de l’excitabilité du réseau (Bracci et al., 1998, Whelan et al., 2000), ou de l’activation spécifique des neurones glutamatergiques (Hagglund et al., 2010, Talpalar and Kiehn, 2010). De plus, des études pharmacologiques ont montré que l’activité rythmique persiste en absence de la transmission synaptique inhibitrice (Bracci et al., 1996), ainsi qu’à la suite d’une hémi-lésion (Kjaerulff and Kiehn, 1996, Bonnot and Morin, 1998, Whelan et al., 2000). Cependant, la coordination entre la gauche et la droite est maintenue uniquement en présence de la commissure ventrale (Kjaerulff and Kiehn, 1996), confirmant l’implication d’interneurones commissuraux dans la coordination bilatérale. De plus, l’ajout d’antagonistes glycinergiques et GABAergiques dans la solution de liquide céphalorachidien artificielle perfusant la moelle épinière isolée de rat nouveau-né induit une activité rythmique avec une synchronisation bilatérale, confirmant la nature principalement inhibitrice des connexions établies par ces interneurones commissuraux (Bracci et al., 1996).
15
Plusieurs populations d’interneurones spinaux excitateurs et inhibiteurs impliquées dans l’activité locomotrice ont été identifiées sur la base de leurs propriétés membranaires, de leur connectivité et de leur patron d’activité (pour revue, Kiehn 2006). Mais c’est surtout au cours de ces 20 dernières années que de nouvelles approches expérimentales, combinant outils génétiques et moléculaires, ont permis l’identification de différents types d’interneurones spinaux et la caractérisation de leurs rôles au sein des CPGs locomoteurs des mammifères (Jessell, 2000, Lee and Pfaff, 2001, Lanuza et al., 2004, Gosgnach et al., 2006, Zhang et al., 2008, Goulding, 2009, Garcia-Campmany et al., 2010, Gosgnach, 2011, Talpalar et al., 2013, Zhang et al., 2014).
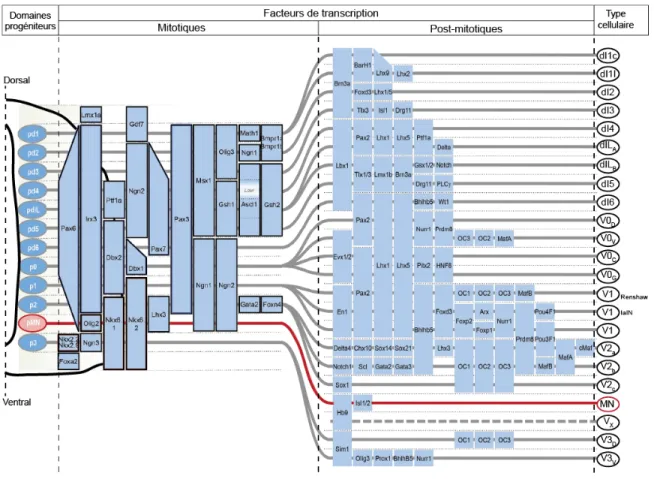
Pendant le développement embryonnaire, différents types de cellules sont générés le long de l’axe dorso-ventral du tube neural., Très tôt au cours du développement, l’action de différentes molécules sécrétées le long de la ligne médiane conduit à la subdivision spatiale dorso-ventrale de domaines progéniteurs qui acquièrent l’expression de différentes combinaisons de facteurs de transcription (pour revue, Jessell 2000). Selon les facteurs de transcriptions qu’elles expriment, les cellules du tube neural se divisent en 11 populations distinctes (dI1-dI6, V0-V3, VMN) qui peuvent être observées dès le 10ème jour embryonnaire
(E10) chez la souris, puis en 23 sous-populations (Figure 0.6). Le profil d’expression de facteurs de transcription détermine l’identité cellulaire (Bang and Goulding, 1996, Goulding and Lamar, 2000, Goulding and Pfaff, 2005), le patron de projection axonale (Betley et al., 2009) et le type de neurotransmetteurs exprimés (Cheng et al., 2005, Mizuguchi et al., 2006, Pillai et al., 2007). Les neurones partageant le même profil d’expression de facteurs de transcription partagent donc potentiellement les mêmes caractéristiques et le même rôle fonctionnel dans la locomotion. Grâce aux outils génétiques modernes, il est possible de générer des souris dans lesquelles une population neuronale spécifique peut être ciblée, grâce aux facteurs de transcription qu’elle exprime, afin de la visualiser par l’expression de protéines fluorescentes, de la stimuler, de l’inhiber ou de la détruire. Ces manipulations génétiques nous permettent ainsi d’étudier l’implication de populations neuronales identifiées génétiquement dans le contrôle moteur et locomoteur. Ainsi, de nombreuses études associant des techniques de génétique moléculaire, d’électrophysiologie et de traçage anatomique ont permis d’identifier 6 populations d’interneurones spinaux (dI6, V0, V1, V2, V3 et Hb9) susceptibles de jouer un rôle clef dans l’activité locomotrice.
16
Figure 0.6 : Détermination de l’identité neuronale des populations neuronales spinales. Représentation schématique des étapes de la détermination génétique de l’identité cellulaire des populations neuronales spinales. La progression développementale, depuis les domaines progéniteurs jusqu’aux neurones post-mitotiques, est représentée de gauche à droite. (Adapté de Alaynick et al., 2011).
3.1 Les interneurones dI6
Les interneurones dl6 expriment les facteurs de transcriptions Lbx1 (Gross et al., 2002) et WT1 (Goulding, 2009) et sont localisées dans la lamina VII/VIII de la moelle épinière (Gross et al., 2002). Cette population neuronale est composée de neurones inhibiteurs ipsilatéraux et commissuraux (Goulding, 2009, Rabe et al., 2009), dont la plupart ont une activité rythmique pendant la locomotion fictive (Dyck et al., 2012). Sur la base de leurs propriétés électrophysiologiques intrinsèques (propriétés membranaires et patrons de décharge), les neurones dI6 peuvent être divisés en deux populations (Dyck et al., 2012). L’une présente des bouffées de potentiels d’action faiblement coordonnées à l’activité ENG des racines ventrales pendant un épisode de locomotion fictive. Cependant, ces neurones montrent des propriétés membranaires intrinsèques similaires à celles des neurones oscillants, suggérant une implication dans l’initiation du rythme locomoteur. La seconde population de neurones présente un patron de décharge rythmique en phase avec
17
l’activité ENG des racines ventrales, mais ne possède aucune des propriétés membranaires caractéristiques des neurones oscillants.
Plus récemment, Dmrt3 a été identifié comme un nouveau marqueur d’une sous-population des interneurones dI6 (Andersson et al., 2012, Vallstedt and Kullander, 2013). Les interneurones Dmrt3 sont inhibiteurs et des expériences de traçage anatomique rétrograde ont révélé des projections directes sur les motoneurones ipsilatéraux et controlatéraux au muscle injecté (Vallstedt and Kullander, 2013). De manière intéressante, certains chevaux portant une mutation spontanée de Dmrt3 présentent un comportement locomoteur spécifique, avec l’apparition du pas amble, type de démarche caractérisé par la synchronisation latérale des membres avants et arrières (Andersson et al., 2012). La mutation Dmrt3-/- affecte aussi la
locomotion fictive néonatale des souris, qui est caractérisée par un rythme irrégulier et une mauvaise coordination des activités ENG des racines ventrales homolatérales reliées aux fléchisseurs et aux extenseurs, ainsi que des racines ventrales gauche et droite d’un même segment spinal (Andersson et al., 2012, Vallstedt and Kullander, 2013). Ces défauts fonctionnels sont accompagnés par une augmentation du nombre de neurones WT1 et une diminution du nombre d’interneurones commissuraux dans la moelle épinière des souris mutantes Dmrt3-/- (Andersson et al., 2012, Vallstedt and Kullander, 2013). Bien que ces
défauts anatomiques et fonctionnels du circuit locomoteur spinal résultent probablement de changements de l’identité développementale des interneurones dI6, l’étendue de la réorganisation du réseau spinal de ces mutants reste inconnue. Ainsi, l’ensemble de ces résultats suggère que les interneurones dl6 pourraient être une composante du circuit locomoteur spinal dont le rôle précis reste à élucider.
3.2 Les interneurones V0
La population d’interneurones V0 est localisée dans la partie ventro-médiane de tous les segments de la moelle épinière (Pierani et al., 1999) et est constituée d’environ deux tiers de neurones inhibiteurs et d’un tiers de neurones excitateurs (Lanuza et al., 2004). Les neurones V0 expriment le facteur de transcription Dbx1 et sont répartis en deux sous-populations d’interneurones commissuraux: les neurones V0V expriment
le facteur de transcription Evx1 et sont localisés plus ventralement, alors que les neurones V0D n’expriment
pas Evx1 et sont plus dorsaux (Moran-Rivard et al., 2001, Pierani et al., 2001). Cependant, un troisième groupe de neurones, exprimant le facteur de transcription Pitx2, a également été mis en évidence. Cette sous-population, appelée V0C/G, est constituée de neurones excitateurs (soit glutamatergiques (V0G) soit
cholinergiques (V0C)) qui projettent ipsilatéralement sur les motoneurones (Zagoraiou et al., 2009). Les
neurones V0C sont la seule source de contacts synaptiques cholinergiques (boutons C) sur les
motoneurones, et ils présentent un patron de décharge rythmique en phase avec l’activité ENG des racines ventrales pendant la locomotion fictive (Zagoraiou et al., 2009). L’absence des trois sous-populations de