ACTIVITE SAISONNIERE DE TROIS ESPECES DE LONGICORNES ET SUIVI DE LA PROGRESSION DES DÉGÂTS CAUSÉS PAR LE LONGICORNE NOIR
APRÈS LE PASSAGE DU FEU EN FORÊT BORÉALE
Mémoire présenté
à la Faculté des études supérieures et postdoctorales de l'Université Laval dans le cadre du programme de maîtrise en sciences forestières
pour l'obtention du grade de Maître es sciences (M. Se.)
DEPARTEMENT DES SCIENCES DU BOIS ET DE LA FORET FACULTÉ DE FORESTERIE ET DE GÉOMATIQUE
UNIVERSITÉ LAVAL QUÉBEC
2013
La qualité du bois récupéré après le passage des feux de forêt se détériore rapidement sous l'action de plusieurs espèces de longicornes qui profitent de cette soudaine disponibilité pour leur reproduction, nutrition et croissance. Ce projet visait à fournir des outils à l'industrie du sciage pour réduire les pertes économiques causées par les longicornes. Dans un premier temps, la colonisation et la ponte du longicorne noir {Monochamus scutellatus scutellatus (Say)) et autres longicornes ont été caractérisées dans cinq brûlis récents. L'abondance des adultes capturés dans des pièges à interception et des larves retrouvées dans les bûches variait selon les brûlis. Lorsque les feux ont fait rage tôt en mai, la colonisation des longicornes s'est effectuée moins intensément. La ponte saisonnière du longicorne noir suivait de près l'activité des adultes, contrairement à celle de Acmaeops proteus proteus (Kirby) où cette dernière était décalée. Ensuite, grâce à la technologie de la tomodensitométrie axiale (CT-Scan), l'estimation de la progression des dommages à plusieurs régimes de température différents selon le type d'essence a été possible. Aux températures les plus élevées (24°C et 28°C), les larves se sont développées rapidement passant ainsi moins de temps dans les tissus sous-corticaux que celles évoluant dans les bûches soumises à de basses températures. Les taux de développement sous-cortical et de progression de la profondeur des galeries mesurées étaient légèrement plus élevés chez l'épinette noire {Picea mariana Mill.) que chez le pin gris {Pinus banksiana Lamb.). Aucun oeuf n'a éclos et aucune larve ne s'est développée sous l'écorce à 12°C La progression des galeries dans les bûches soumises à 20°C ont montré des patrons de progression différents indiquant que les larves peuvent effectuer une diapause sous l'écorce ou dans le bois. À la lumière de ces résultats, des dommages importants peuvent survenir l'année même suivant le passage des feux démontrant la nécessité de débuter la récupération du bois brûlé le plus rapidement possible afin de limiter les pertes économiques causées par les larves de longicorne noir.
Abstract
The quality of post-fire wood harvested rapidly decreases under the influence of several species of longhorned beetles benefiting from this sudden availability of dead wood for reproduction, feeding and growing. This study aimed to reduce economic losses caused by longhorned beetles for sawmill industry. The first step was to characterize colonization and egg laying behavior of the whitespotted sawyer {Monochamus scutellatus scutellatus (Say)) and other longhorned beetles in five recent bums. Adults caught in flight-intercept traps and larvae found in logs varied in abundance among burns. Results show that colonization was slower when fires happened in early May. Egg laying followed adult activity for the whitespotted sawyer while it was delayed in time for Acmaeops proteus proteus (Kirby). The second step was to estimate damage progression under different temperature regimes and on different tree species, using the axial tomograph technology (CT-Scan). Results show that subcortical development rate and gallery depth progression rate were higher on black spruce {Picea mariana Mill.) than on jack pine {Pinus banksiana Lamb.). At 12°C there has been no hatching nor larval development under bark, while at 24°C and 28°C larvae rapidly penetrated sapwood. Patterns of gallery progression in logs reared at 20°C revealed that longhorned beetle larvae can enter diapause under bark or into sapwood. The study shows that significant insect-induced wood damage can appear during the first year following a forest fire. It is therefore necessary to start salvage logging as soon as possible after a fire in order to minimize the economic losses caused by whitespotted sawyer larvae.
Avant propos
Ce mémoire de maîtrise a été rédigé en anglais sous forme de deux articles scientifiques destinés à être soumis à une revue scientifique internationale au cours de l'année 2012. Ces articles représentent chacun un chapitre et s'insèrent entre une introduction et une conclusion générale écrite en français. Le premier chapitre, portant sur l'arrivée et l'oviposition des longicornes dans les brûlis récents après le passage des feux, a fait l'objet d'une récolte de données sur le terrain dans cinq brûlis récents. À l'aide de mon équipe, une véritable chasse aux feux de forêt s'est effectuée durant les étés 2009-2010. Ensuite, j'ai traité les échantillons en laboratoire, identifié les larves et les insectes capturés pour finalement compiler et analyser les données. L'expérience constituant le deuxième chapitre a demandé davantage de logistique puisqu'une technologie innovatrice et basée sur les rayons X a été utilisée pour suivre la progression des dommages effectués par les larves du longicorne noir. Dans ce but, avec l'aide d'un spécialiste du logiciel Matlab, j'ai participé au développement d'une programmation composée d'algorithmes afin de déterminer automatiquement la profondeur de chacune des galeries contenues dans les échantillons. Ensuite, j'ai effectué le traitement des données puis l'analyse en utilisant une modélisation non linéaire de Chapman-Richards, qui m'était alors totalement inconnue. La combinaison de ces deux chapitres permettra de développer prochainement le premier modèle de prédictions des dommages effectués par les larves du longicorne noir après le passage des feux et sera fort utile pour les gestionnaires de l'industrie forestière afin de planifier la récupération des bois brûlés de façon à limiter les pertes économiques. Encadré et soutenu par mon directeur et mes codirecteurs, j'ai entièrement rédigé ce mémoire. Sans le support financier du Consortium de recherche iFor, du Conseil de recherches en sciences naturelles et en génie du Canada, du Ministère des Ressources Naturelles et de la Faune du Québec, du Conseil de l'Industrie Forestière du Québec, du Service canadien des Forêts et de la Société de Protection des Forêts contre les Insectes et les Maladies du Québec, ce projet de recherche n'aurait pu être réalisé avec autant de succès.
Remerciements
Je voudrais exprimer toute ma gratitude envers mon directeur de recherche, le Dr. Éric Bauce pour son appui et la confiance portée à mon égard tout au long de ma maîtrise. Ses connaissances, ses conseils et ses anecdotes m'ont permis de réaliser ce beau projet de recherche et de me surpasser sans cesse! Je voudrais remercier mon codirecteur de recherche, le Dr. Christian Hébert du Service canadien des forêts (SCF-RNCan) avec qui j'ai eu beaucoup de plaisir à travailler et qui m'a toujours épaulé et guidé dans les moments les plus critiques. L'assistance technique et les ressources auxquelles j'ai pu avoir accès dans son laboratoire m'ont vraiment permis de mener à terme ce projet avec succès. Merci spécialement au Dr. Richard Berthiaume, coordonnateur du Consortium iFor, pour son encadrement, son appui et les nombreuses discussions dans le domaine forestier bien sûr, mais aussi dans plusieurs autres domaines qui ont contribué à mon développement personnel et professionnel. J'adresse aussi mes remerciements au Dr. Bernard Long, mon codirecteur de recherche de l'Institut de la Recherche Scientifique (INRS), pour son aide et toutes les ressources technologiques qu'il a mis à ma disposition pour atteindre mes objectifs.
Je voudrais également remercier Yves Dubuc du SCF-RNCan pour son dévouement et sa patience à toute épreuve envers ma personnalité parfois déroutante. Également, merci à Martin Charest pour son support et les nombreuses missions spéciales qui ont sûrement amélioré le sort du monde. Pour sa bonne humeur, son encouragement et sa collaboration, je remercie Marcel Trudel de la SOPFEU. Pour leur aide et assistance technique concernant le tomographe à rayons X et Matlab, je vous remercie au plus haut point Louis-Frédéric Daigle et Jacques Labrie de l'INRS. J'aimerais remercier Louis Morneau et les collègues du Ministère des Ressources Naturelles et de la Faune du Québec pour leur support.
J'exprime aussi toute ma gratitude envers les étudiants de l'Université Laval Jean-François Bourdon (le viking), Mathieu Couture (le ninja), Patrick Descôtes (l'homme des hommes), Jean-Philippe Gendron-Bouchard (JP la brute) et Etienne Godbout, qui composaient l'équipe de rêve de l'été 2009 avec qui il a été possible de réaliser l'impossible. Merci aussi
Meunier (ma meilleure aussi), Alessandro Dieni (mon meilleur aussi) et Jean-Philippe Boyer ainsi que Yan Bouchard, Maxime Couette et Sandra Losier du Cégep de Ste-Foy qui se sont greffés à l'équipe par la suite. Aussi, Mélanie Couture, Louis-Philippe Rodier, Francis Grenier, Louis-Philippe Ménard, David Saint-Pierre, Maxime Tanguay, Martin Pelletier, Gabriel Roy, Claudie Lachance, Julien Dessureault, Albert Paquet, Cyrille Forchelet, Laurence Saucier et Mathilde Maïsano, Paule Huron, je vous remercie pour votre aide.
Une attention particulière aux membres de mon laboratoire d'ÉcoDIF, soit Olivier Norvez (D.G.G.B.H.), Jonathan Boucher pour son aide précieuse (John Bouc), Yannick Cadorette-Breton (Ti-Yannic), Olivier Jeffrey (Ti-cœur), Francis Desjardins (Franki), Ermias T. Azeria (Erbi), Jean-Philippe Légarê (JP), Pierre-Marc Brousseau (Broue), Éric Domaine (Le Rick), Mayerline Murillas pour leur aide et les bons moments passés ensemble, merci! Merci aussi à Nathalie Delvas, Roberto Quezada, Alvaro Sebastian Fuentealba Morales, Lukas Seehausen et Rosemarie Vallières du laboratoire d'entomologie forestière de l'Université Laval.
Finalement, j'aimerais vraiment remercier mes parents, mes frères et mes bons amis, spécialement Oli, Maxime, Amélie, Cédric, Jérémy, Orphé, Léo, Rémy, Valérie, Steph, Karine, Andrée-Anne, David, Hubert et les « Sarah-Maude » pour leur support durant ma maîtrise.
La force d'un homme ne se mesure pas au nombre défais où il tombe mais à sa capacité de se relever
Table des matières
Résumé i Abstract ii Avant propos iii Remerciements iv Table des matières vii Liste des tableaux ix Liste des figures x Liste des annexes xii Introduction générale 1
Forêt boréale, une dynamique changeante 1
Récupération du bois brûlé 2
Problématique 3 Mécanismes d'attraction et variables environnementales 4
Les longicornes les plus abondants au Québec dans les habitats récemment brûlés 5
Le cycle biologique du longicorne noir 6
Tomographe à rayon X 7
Objectifs 8 Adult seasonal activity and egg deposition of three longhorned beetles (Coleoptera:
Cerambycidae) shortly after fire in boreal black spruce forest 9
Résumé 10 Abstract 11 Introduction 12 Materials and methods 13
Study areas 13 Insect sampling 14 Statistical analysis 15
Results 16 Longhorned beetle diversity and abundance 16
Adult seasonality and egg deposition 16 Relation between M. s. scutellatus adult abundance and egg density 17
Discussion 17 Acknowledgement 21
References cited 22 Effect of temperature and tree species on damage progression of whitespotted sawyer,
Monochamus scutellatus scutellatus (Say), larvae in recently burned logs 32
Résumé 33 Abstract 34 Introduction 35 Materials and methods 37
Origin of logs used in the experiment 37 Whitespotted sawyer breeding 38 Rearing and CT scanning 38
Data analysis 39 Statistical analysis 40
Results 41 Model fitting 41
Chapman-Richards models 41 Subcortical development and gallery depth progression rates 43
Discussion 43 Acknowledgements 48
Reference cited 49 Conclusion générale 59
Arrivée et oviposition des longicornes 59 La vitesse de progression des larves de longicorne noir dans le bois 60
Effet de l'essence 61 Effet de la température 61 Implication pour l'aménagement des brûlis et perspectives d'avenir 62
Nouvelle approche et aménagement écosystémique 64
Références 65 Annexes 71
Liste des tableaux
Tableau 1-1: Abundance of adult longhorned beetles caught in trunk-window traps and overall egg density (per m2 of bark) in five burned black spruce forests the same year as
the fire, in Quebec, Canada 26 Tableau 2-1 : Number of logs and galleries used in analysis as a function of temperature and
tree species 53 Tableau 2-2: Subcortical development and gallery depth progression rates as a function of
Figure 1-1: Location of the five burns studied in Québec, Canada A) Chapais B) Assinica
C) Rivière Serpent D) Lac Mondonac E) Lac Smokey 25 Figure 1-2: Relationship between the mean abundances of Monochamus s. scutellatus and
Acmaeops p. proteus in trunk-window traps among five burns studied in Québec in
2009 and 2010 27 Figure 1-3: Adult seasonal abundance and seasonal egg deposition for three longhorned
beetle species in three 2010 burns. Solid lines fitted for adult seasonality and dotted lines for egg deposition. To facilitate comparisons, adult abundance of the three longhorned beetle species was multiplied by 3 for Lac Mondonac and Lac Smokey burns. The period during which fire burned is represented by the black line which becomes dotted for the Lac Smokey. Graphics at the bottom show the mean daily temperature (C°) for each burn collected by Hobo data loggers throughout the sampling
season 28 Figure 1-4: Adult seasonal abundance and seasonal egg deposition for three longhorned
beetle species in two 2009 burns. Solid lines fitted for adult seasonality and dotted lines for egg deposition. The period during at which fire burned is represented by the black line. Graphics at the bottom show the mean temperature (C°) for each burn collected by
Hobo data loggers throughout the sampling season 29 Figure 1-5: Relationship between mean adult abundance in trunk-window traps and egg
density of Monochamus s. scutellatus in five burns according to the 1st week of captures (A), the first 2 weeks of captures (B) and for the entire season of captures (C) after the end of fire. The empty circle represents the Assinica burn larval density measured the year after the fire. This measure was corrected for taking into account the average mortality (83,7%) that occurs between egg deposition and larval entrance into
the wood based on Rose (1957) 30 Figure 1-6: Seasonal propagation of the fire in the Lac Smokey bum in 2010. The star
represents our sampled plot 31 Figure 2-1 : Larval gallery depth of Monochamus scutellatus scutellatus as a function of
time at different temperatures. Solid lines fitted for black spruce (BSP) and dotted lines for jack pine (JPI). Figures: A) BSP: rM.99, JPI: r^O.99 B) BSP: r^O.99, JPI:
1^=0.97 C) BSP: 1^=0.98, JPI: 1^=0.99 D) BSP: r^O.99, JPI: r^O.96 54 Figure 2-2: Larval gallery depth of Monochamus scutellatus scutellatus as a function of
time at 20°C Galleries were classified into 2 categories according to the time of
Figure 2- 3: Larval gallery depth of Monochamus scutellatus scutellatus as a function of time at 20°C Solid lines fitted for black spruce and dotted lines for jack pine. Galleries were classified into 2 groups according to the time of larval entrance: early and late. The long dashed at 20°C represents the second galleries progression fitted by
Chapman-Richards model 56 Figure 2- 4: Subcortical development A) and gallery progression rates B) of Monochamus
scutellatus scutellatus as a function of temperature. Solid lines fitted for black spruce
and dotted lines for jack pine 57 Figure 2-5: Gallery progression of a Monochamus scutellatus scutellatus larva in the same
Liste des annexes
Annexe 1. Photographie d'un piège à interception (Trunk window trap (TWT) utilisé pour
capturer les longicornes adultes, © Jean-François Bourdon 71 Annexe 2. Photographie d'un arbre protégé de l'attaque des longicornes par un papier noir
en construction. Les arbres ont été par la suite dénudés de leur protection pour être exposés à la ponte des longicornes pendant des périodes pré-déterminées, ©
Jean-François Bourdon ...72 Annexe 3. Photographie illustrant la méthode d'écorçage d'une bûche provenant d'un arbre
brûlé ayant été exposé aux longicornes, © Catherine Dion 73 Annexe 4. Photographies des différents niveaux de sévérité de brûlage retrouvés dans les
brûlis lors de ce projet de recherche, © Sébastien Bélanger (gauche et milieu), Jonathan
Boucher (droite) 74 Annexe 5. Photographie d'une cage contenant une bûche sur laquelle les longicornes noirs
femelles ont pondu leurs œufs dans l'écorce (haut). Photographie de la serre où les cages ont été placées pendant trois jours à une température et humidité contrôlée au
centre de foresterie des Laurentides, Québec, Québec (bas), © Sébastien Bélanger 75 Annexe 6. Photographies représentant des galeries creusées dans le bois par les larves du
longicorne noir, © Sébastien Bélanger, © Produits forestiers Résolu 76 Annexe 7. Photographie du tomographe à rayons X localisé à l'Institut National de la
Recherche Scientifique, Centre Eau, Terre et Environnement, Québec, Québec, ©
Sébastien Bélanger 77 Annexe 8. Image numérique de 0.6 mm d'épaisseur d'une résolution de 0.4 x 0.4 mm par
pixel obtenue lors de la numérisation d'une bûche d'épinette par le tomographe à
Forêt boréale, une dynamique changeante
Au cours des dernières décennies, suite à l'augmentation de la concentration des gaz à effet de serre (dioxyde de carbone, méthane, etc.) causée en grande partie par l'intensification des activités humaines, le réchauffement climatique n'a cessé de s'accentuer (Cox et al., 2000; Flannigan et al., 2005; Kerr, 2007; Esenhauer et al., 2012). Ce réchauffement climatique influence la dynamique des forêts par l'augmentation de la fréquence, de la durée et de l'intensification des catastrophes naturelles (Overpeck et al., 1990; Serreze et al., 2000; Soja et al., 2007). Dans la forêt boréale, le feu est la principale perturbation naturelle (Johnson, 1992; Payette, 1992; Stocks et al., 2002; Flannigan et al., 2005). De par ses caractéristiques, le feu permet de diversifier et de structurer la forêt en créant une mosaïque forestière (Bergeron et al, 2010) et une grande variété d'écosystèmes (Dajoz,
1998) permettant de maintenir une grande diversité de communautés végétales et animales (Pastor et Mladenoff, 1992). Or, cette perturbation devient de plus en plus importante économiquement puisque la fréquence des incendies et les superficies brûlées ont augmenté au cours des 4 dernières décennies et que cette tendance à la hausse devrait se poursuivre (Gillett et al., 2004). L'intérêt de mieux comprendre la dynamique des feux de forêt pour ainsi l'intégrer aux nouvelles approches d'aménagement de la forêt s'est rapidement développé au cours de la dernière décennie (Bergeron et al., 2004). Comme l'économie de plusieurs régions du Canada dépend en grande partie de l'exploitation forestière, les incendies forestiers ont des conséquences importantes pour ces communautés. C'est ainsi qu'une nouvelle génération de modèles de prédiction pour les forêts ont vu le jour afin d'estimer par exemple, la croissance et la succession forestière après une perturbation ou une intervention majeure (Fortin et Langevin, 2010; Pothier et Auger, 2011), la régénération forestière naturelle après feu (Boulfroy, 1996) et le comportement des incendies de forêts (Wotton et al., 2009).
Depuis le début des années 90, le secteur forestier québécois a traversé une importante crise (Beauregard et Bouthillier, 1993; MRNF, 2008; Mockler et al., 2011). La diminution de la demande pour les produits fabriqués à partir du bois canadien était en grande partie responsable des problèmes du secteur forestier (Mockler et al., 2011). La réduction structurelle de la demande en papier journal, la concurrence des économies en développement et la diminution cyclique de la demande de l'industrie de la construction sont les principales causes qui ont mené à la crise (Madore et Bourdages, 1992; Mockler et al., 2011). D'autres facteurs ont aggravé la problématique tels que la force de la devise canadienne, la hausse des coûts de l'énergie et le manque d'accessibilité au crédit (Madore et Bourdages, 1992; Mockler et al., 2011). De plus, les usines déjà existantes et devenant de plus en plus désuètes se retrouvaient éloignées des forêts économiquement exploitables et la ressource ligneuse susceptible d'être économiquement exploitable se raréfiait (Madore et Bourdages, 1992). Le rapport de la commission d'étude sur la gestion des forêts publiques québécoises (aussi connu sous le nom de rapport Coulombe) comportait une série de recommandations visant à assurer la pérennité de la ressource (Coulombe et al., 2004). Entre autre, la diminution de la possibilité forestière annuelle de 20% était fortement recommandée dans les peuplements conifériens. En 2006, suivant ces recommandations, le Forestier en chef dévoilait une baisse moyenne de 21,9% de la possibilité annuelle de coupe des forêts publiques du Québec pour les années 2008 à 2013 (Paillé et al., 2007). Ainsi, dans les régions les plus affectées par la réduction de la possibilité forestière (plus de 20%), l'impact économique du feu pouvait être d'autant plus important, en termes de perte de ressources ligneuses, si aucune récupération n'était effectuée dans les brûlis. Depuis 1986, le gouvernement du Québec a dû consentir beaucoup d'efforts afin d'obliger les compagnies forestières à prioriser la récolte du bois brûlé au lieu du bois vert sous peine de réductions des quotas alloués. L'article 79 de la Loi sur les forêts (L.R.Q., c. F-4.1) a eu pour effet d'accroître le volume des coupes de récupération de 3 millions de m de bois brûlé par hectare en 1991 à plus de 10 millions de m3 en 1996 (Purdon et al., 2002). En fait, les bénéficiaires d'un contrat d'approvisionnement et d'aménagement forestier (CAAF) doivent se conformer au plan spécial d'aménagement élaboré par le Ministère des
forestières où ont eu lieu des perturbations naturelles (Purdon et al., 2002; Nappi et al., 2004). Ainsi, la récupération du bois a pu s'effectuer davantage lors des dernières années où de nombreux feux de forêt ont sévi et que d'immenses superficies ont été brûlées. Dans la région du Saguenay-Lac Saint Jean en 2005, 95% du bois traité dans les usines de sciage provenait de la récupération du bois dans les brûlis (Saint-Germain et Greene, 2009). Dans cette optique, la récolte du bois provenant des brûlis récents est devenue, au Québec ainsi qu'au Canada, une stratégie pouvant contribuer à réduire l'impact économique des feux et à maintenir un fonctionnement adéquat dans l'industrie forestière (Saint-Germain et Greene, 2009; Nappi et al., 2011).
Problématique
La récupération du bois dans les brûlis ne se fait pas sans difficulté puisqu'elle dépend et varie selon l'accessibilité du territoire, la nature des peuplements touchés et la demande des produits sur le marché du bois (Nappi et al., 2011). De plus, une vraie course contre la montre s'effectue afin de récupérer la ressource forestière avant de subir des pertes économiques importantes de la valeur des produits du bois causés par les insectes xylophages lorsque ce bois est destiné aux usines pour la production de bois de sciage. Actuellement, il n'existe aucun moyen fiable de prédire les dommages causés par ces insectes dans les brûlis récents. Autrefois, on laissait sous-entendre que les dommages de longicornes, les principaux insectes xylophages causant les dommages aux arbres après feu, devenaient importants à partir de la deuxième année suivant le passage du feu. Or, des observations récentes montrent que les dommages peuvent apparaître dès la première année suite au passage du feu (communications personnelles : Michel Deshaies, Barrette-Chapais ltée et Serge Bergeron, Ministère des Ressources naturelles et de la Faune, Chibougamau). De plus, il semble exister des variations dans la vitesse de progression des dommages en fonction des essences forestières, de la température, des régions, etc. En l'absence de modèle de prédiction efficace afin d'optimiser la récupération, les gestionnaires ont peu d'outils pour les éclairer à prendre de bonnes décisions afin d'optimiser la récupération du bois brûlé. Pour développer un tel modèle, trois étapes fondamentales doivent être
progression des larves dans le bois en fonction de la température et des essences forestières.
Mécanismes d'attraction et variables environnementales
Dans cet habitat nouvellement créé que représente un brûlis (Wikars, 2002), certains insectes xylophages tirent avantage de la grande abondance de bois mort ou moribond brûlé soudainement disponible pour se nourrir et se reproduire (Wikars, 1994; Boulanger et Sirois, 2007). Plusieurs espèces opportunistes de Cerambycidae et de Buprestidae sont attirés vers cette ressource par l'odeur de la fumée (Wikars, 1992), les composés volatils émis (Chénier et Philogène, 1989; Peddle, 2000) et la chaleur (Dajoz, 1998). En fait, afin de profiter de cette ressource, plusieurs espèces d'insectes xylophages auraient développé des adaptations telles que des récepteurs antennaires et organes sensoriels thoraciques sensibles à la fumée (Schûtz et al., 1999; Suckling et al., 2001) et au rayonnement infrarouge (Evans, 1966; Schmitz et al. 2002), leur permettant de localiser et de converger rapidement vers les territoires récemment brûlés (Hosking et Bain, 1977). Concernant les longicornes du genre Monochamus, il a été démontré qu'ils sont attirés par certains composés phénoliques volatils tels que les monoterpènes et l'éthanol (Phillips et al., 1988; Chénier et Philogène, 1989; de Groot et Nott, 2001). Cependant, ces mécanismes d'attraction sont directement influencés par des facteurs environnementaux. En effet, la superficie et la sévérité du feu (Richmond et Lejeune, 1945; Ross, 1960; Raske, 1972; Saint-Germain et al., 2004), la distance de la forêt verte, les caractéristiques des différentes espèces d'arbres (Saint-Germain et al., 2004) ainsi que le moment de l'émergence des adultes (Morley, 1939) sont des facteurs pouvant jouer un rôle dans la colonisation, l'abondance/occurrence et l'oviposition des longicornes dans les brûlis.
Au Québec, trois espèces de longicornes sont souvent abondants dans les brûlis soit Acmaeops proteus proteus (Kirby), Arhopalus foveicollis (Haldeman) et Monochamus scutellatus scutellatus (Say) (Saint-Germain et al., 2004b-c; Boulanger et Sirois, 2007; Berthiaume et al., 2010). Gardiner (1957, 1958) a démontré que les larves de Acmaeops p. proteus n'effectuent que des dommages dans les tissus sous-corticaux en se nourrissant du
phloème et du cambium et ne causent aucune perte économique puisqu'elles ne creusent aucune galerie dans le xylème. Pour sa part, Arhopalus foveicollis (Haldeman) est une espèce qui a un rôle écologique important comme source de nourriture pour les pics à dos noirs (Nappi et al., 2010). Bien que cette espèce semble effectuer des galeries dans le xylème, peu de pertes économiques seraient anticipées puisque ses larves se retrouveraient principalement au niveau de la souche et des racines des épinettes et des pins morts (Yanega, 1996). Cependant, une espèce très proche {Arhopalus férus (Mulsant)) présente à travers l'Europe, l'Afrique ainsi que dans la région orientale (Hosking, 1970), cause des pertes économiques importantes au niveau de la tige et de la souche sur une grande variété de pins récoltés dans les chablis et les brûlis (Hosking et Bain, 1977).
En ce qui a trait à Monochamus s. scutellatus, les larves creusent des galeries dans le xylème causant d'importantes pertes économiques (Raske, 1972; Peddle, 2000). Bien que la solidité du bois n'est pas appréciablement réduite (Kennedy, 1957), le bois affecté est déclassé d'un point de vue esthétique sur le marché et peut même être totalement rejeté dans certains cas (Mcintosh et al., 2001). De plus, les exportations peuvent être mises en quarantaine ou retournées à leur port d'origine à cause de la visibilité des dommages (trous) (Shore, 1985). Dans l'ouest canadien, Prebble et Gardiner (1958) ont estimé que les pertes économiques sur le pin blanc {Pinus strobus L.), engendrées par les larves du genre Monochamus, étaient approximativement de 14% pour le bois de faible qualité et au-delà de 33% pour le bois de qualité. Raske et Safranyik (1970), ont démontré que les pertes pouvaient atteindre plus de 30% pour le pin tordu {Pinus contorta Dougl.). Des enquêtes effectuées dans les scieries de la Colombie-Britanique ont permis d'estimer des pertes économiques annuelles de l'ordre de 43 millions SUS causées par les longicornes
important de bien connaître le cycle biologique du longicorne noir pour ainsi être en mesure de prédire ses dommages.
Le cycle biologique du longicorne noir
Le longicorne noir est omniprésent en Amérique du Nord où la forêt est composée de conifères (Peddle, 2000). La majorité des adultes émergent à la fin mai et durant le mois de juin pour ensuite se diriger vers les arbres morts, sénescents ou affaiblis soit par l'âge, un chablis, un feu, une infestation d'insectes ou par des maladies (Peddle, 2000). Les débris de coupes forestières ainsi que les empilements de bois sont également susceptibles d'être attaqués par cette espèce (Martineau, 1985). Les femelles utilisent ces hôtes afin de pondre leurs œufs dans les fissures naturelles de l'écorce, à la fourche de grosses branches (Rose, 1957; Walsh et Linit, 1985), mais également dans des fentes qu'elles creusent elles-mêmes avec leurs mandibules.
L'éclosion des œufs a lieu après une incubation de 9 à 14 jours dépendamment de la température ambiante (Rose, 1957; Peddle et al., 2002). Les jeunes larves creusent ensuite une galerie à travers le phloème pour atteindre le cambium en deux ou trois jours (Wilson, 1962). Pendant deux à trois semaines, les larves des deux premiers stades creusent à la surface du cambium pour s'alimenter (Rose, 1957). Ensuite, au début de l'automne, lorsqu'elles atteignent le troisième stade larvaire, elles commencent à pénétrer le bois d'aubier (Rose, 1957). A la fin de septembre, devenant léthargique avec l'avènement du froid, les larves se réfugient au fond de leur galerie pour passer leur premier hiver (Martineau, 1985). Le printemps suivant, les larves continuent à creuser dans le bois en effectuant souvent dans leur galerie, des changements de direction pour expulser la sciure (Raske, 1972). A la fin de septembre, les larves qui ont atteint le quatrième stade larvaire, se trouvent près de la surface sous l'écorce pour construire une loge pupale dans laquelle elles passeront un deuxième hiver au stade prépupal (Rose, 1957). Avec le retour de la chaleur du mois de mai, la pupaison a lieu dans la loge pupale et le développement des
développement larvaire du longicorne noir peut s'effectuer sur une période de un ou deux ans (Peddle, 2000). Bien que cela est actuellement inconnu chez le longicorne noir, la plupart des espèces du genre Monochamus entrent en diapause obligatoire durant les premiers stades larvaires et également lors du dernier stade larvaire (Rose, 1957; Togashi,
1991b). Par exemple, Monochamus alternatus Hope, réalise un cycle de vie de deux ans en effectuant une première diapause au 1er ou au 2ieme stade larvaire lorsque les conditions naturelles sont défavorables (Togashi, 1991a-b). Par ailleurs, la durée du cycle biologique peut varier en fonction de la date d'oviposition du longicorne noir qui s'échelonne sur quelques semaines durant les mois de juin et de juillet (Wilson, 1962). Certaines larves pourraient également passer le premier hiver au 4ieme stade larvaire; le cycle se déroulerait alors sur une seule année (Peddle, 2000) et pourrait caractériser les individus se trouvant davantage au sud de l'aire de répartition (Rose, 1957) ou exposé à des conditions estivales particulièrement chaudes.
Tomographe à rayon X
Plusieurs études ont jadis estimé les pertes économiques causées par les longicornes dans les brûlis (Richmond et Lejeune, 1945; Prebble et Gardiner, 1958), mais aucune n'a jamais tenté de prédire la progression temporelle des larves et/ou des galeries afin de limiter les pertes économiques. La tomodensitométrie axiale ou scanographie axiale (CT-scan) est une technologie qui a été utilisée lors de ce projet de recherche. Il s'agit d'une technique non-destructive qui permet de visualiser l'intérieur d'un objet en 3 dimensions à l'aide d'images digitalisées avec une grande précision. Davantage axée sur le domaine médical au début, cette technique à été utilisée dans d'autres champs d'application tel que dans le domaine pétrolier (Withjack, 1988; Coles et al., 1991), en minéralogie (Tully et al., 1988; Grevers et al., 1989) et plus récemment en archéologie (Jansen et al., 2006) et sédimentologie (de Montety et al., 2003; Michaud et al., 2003; Gagnoud et al., 2009). Cette approche est utilisée ici pour suivre la progression des galeries du longicorne noir afin de prédire l'apparition des dommages.
Le premier chapitre de ce mémoire de maîtrise vise à déterminer la période d'arrivée et d'oviposition du longicorne noir après le passage de feux de forêt. Ces phénomènes biologiques ont été déterminés dans cinq brûlis récents afin de servir de point de départ pour la création d'un modèle permettant de prédire les dommages de cette espèce. De plus, cette étude vise également à vérifier s'il existe une relation entre le nombre de longicornes adultes capturés à l'aide de pièges à impact fixés au tronc des arbres brûlés et la densité des œufs pondus sur ces derniers afin de prédire rapidement l'intensité des dommages dans le bois brûlé. Dans le deuxième chapitre de ce mémoire de maîtrise, nos objectifs sont de déterminer le taux de développement sous-cortical et la progression de la profondeur des galeries du longicorne noir dans le bois. Ces variables ont été estimées en fonction de la température pour l'épinette noire et le pin gris. Ces résultats serviront à l'élaboration d'un modèle de prédiction des dommages du longicorne noir tenant compte de différentes variables environnementales (date du feu, température, temps entre le passage du feu et le début de la récupération). Il sera ainsi possible d'optimiser la récolte de bois suite au passage du feu.
beetles (Coleoptera: Cerambycidae) shortly after fire in boreal
black spruce forest
1 7 * 1 ' 1
Sébastien Bélanger , Christian Hébert , Richard Berthiaume and Eric Bauce
1 Département des sciences du bois et de la forêt, Faculté de foresterie, de géographie et de géomatique, Pavillon Abitibi-Price, Université Laval, Québec, Canada
2 Natural Resources Canada, Canadian Forest Service, Laurentian Forestry Centre, Quebec, Canada
Résumé
Au Québec, la récupération du bois après le passage des feux de forêt est devenue une pratique courante afin de maintenir un approvisionnement en bois adéquat pour l'industrie forestière. Toutefois, les galeries creusées dans le bois par les insectes xylophages réduisent la valeur économique du bois pour l'industrie du sciage. Parmi les insectes xylophages, le longicorne noir, Monochamus scutellatus scutellatus (Say), est celui qui occasionne le plus de dommage dans la forêt boréale récemment brûlée. Cette étude vise à caractériser la colonisation des arbres par le longicorne noir et les autres espèces de longicorne après le passage des feux de forêt, et de vérifier si l'abondance des longicornes adultes capturés dans des pièges à interception fixés au tronc pourraient être utilisés pour prédire l'oviposition des longicornes. L'abondance des adultes capturés dans les pièges à interception et les larves retrouvées dans les arbres variait selon les brûlis et était plus élevée dans les brûlis se situant davantage au nord. La colonisation des longicornes s'est effectuée rapidement dans les brûlis ayant eu lieu près de la période où les longicornes sont actifs pour la reproduction. La ponte saisonnière du longicorne noir suivait de près l'activité des adultes, mais celle de Acmaeops proteus proteus (Kirby), dont les larves se nourrissent dans les tissus sous-corticaux, était décalée. Les relations entre l'abondance des longicornes noirs adultes et la densité d'oeufs pondus dans les arbres étaient significatives pendant les deux premières semaines de capture ainsi que pour l'ensemble de la saison. Ce résultat suggère que la prévision du niveau d'attaque des arbres pourrait être possible en mesurant l'activité des adultes du longicorne noir au début de la colonisation des brûlis. Cela montre un fort potentiel pour la gestion de la récupération du bois brûlé car un tel outil serait utile pour améliorer la prédiction des dommages.
Abstract
In Quebec, Canada, salvage logging after fire has become a management practice for maintaining timber supply required by the forest industry. However, the galleries burrowed into the wood by woodborers reduce wood value for the sawmill industry. The whitespotted sawyer, Monochamus scutellatus scutellatus (Say) is the most damaging wood-boring insect in recently burned boreal forest. This study aimed to characterize bum colonization by the whitespotted sawyer and other longhorned beetles after wildfires, and to verify if adult beetles caught in traps could be used to forecast woodborer larval density. The abundance of adults caught in traps and of larvae found in trees varied among burns and was greater in northern burns. Longhorned beetle colonization was rapid in burns that occurred near their seasonal period of activity. The seasonal egg deposition of whitespotted sawyer, closely followed adult activity but that of Acmaeops proteus proteus (Kirby), which feed on subcortical tissues, was delayed. Relationships between whitespotted sawyer adult abundance and overall egg density were significant when considering the first two weeks of capture as well as the overall season. This suggests that forecasting woodborer attack level might be possible to some extent by measuring the early colonization of the whitespotted sawyer aduts. This shows a high potential for management because such tool would be useful to improve damage prediction.
Introduction
Wildfire is one of the most important natural disturbances driving forest succession in the boreal forest (Bergeron et al., 2001). The variations in fire frequency, intensity and area affected generate a mosaic of various stand ages which structure the forest landscape and the regional biodiversity (Payette et al., 1989; Johnson, 1992; Pastor and Mladenoff, 1992; Nappi et al., 2004). This disturbance generates tremendous amounts of dead and dying trees which represent food resources and reproduction habitats for several xylophagous insects (Wikars, 1992; de Groot and Nott, 2004; Saint-Germain et al., 2004c). Some beetles have developed sensory organs to detect smoke and heat for locating bums (Evans, 1966; Schutz et al., 1999; Schmitz et al., 2000). Beetles may also use attractive volatiles (e.g. ethanol, monoterpenes and terpenoids) released from stressed or dying trees in locating potential hosts (Parmelee, 1941; Dyer and Seabrook, 1978; Chénier and Philogène, 1989). Many studies in boreal forest have already shown that several xylophagous species are abundant after fire (Saint-Germain et al., 2004b; Boulanger and Sirois, 2007). The longhorned beetles (Coleoptera: Cerambycidae) Acmaeops proteus proteus (Kirby), which feed on subcortical tissues (Morley, 1939; Gardiner, 1958), and Arhophalus foveicollis (Haldeman), which feed at the base of trees (Yanega, 1995), are among the most abundant species after fire in the boreal forest (Boulanger and Sirois, 2007; Boulanger et al., 2010). However, the most damaging woodborer after fire for forest industries is definitely the whitespotted sawyer, Monochamus scutellatus scutellatus (Say), because it digs deep galleries into trees (Gardiner, 1957; Peddle, 2000; Purdon et al., 2002; Nappi et al., 2004) thus reducing lumber value.
The biology and the life cycle of the first two longhorned beetles are poorly known, but the whitespotted sawyer biology and ecology are better known. Its life cycle lasts usually two years in the northern part of its distribution but only one year in the southern part (Cerezke, 1977). However, in Québec, we estimated that up to 10 % of individuals of a boreal forest population completed their life cycle in 1-yr (unpublished data) while in Alberta, 25 % of the individuals went through a 1-yr life cycle (Cerezke, 1977). After completing development of the first two larval instars under bark while feeding on
subcortical tissues, most larvae enter the third instar by early September and begin tunnelling into the wood. By late September, larval tunnels may extend deeply into the wood (Rose, 1957). Indeed, larval damage can be high even few months after fire (Richmond and Lejeune, 1945). The maximum depth of galleries can reach over 10 cm one year after fire (Richmond and Lejeune, 1945; Ross, 1960). After winter diapause, whitespotted sawyer larva resumes digging towards the heartwood and then turn back and continue digging but towards outer bark thus creating a characteristic U-shaped gallery (Wilson, 1962). Thus, damage continues all over the summer of the year following fire.
Over the last 20 years in Québec and Canada, salvage logging after wildfire has become a management practice for maintaining the timber harvest volumes necessary for sustaining forest industry production (Lowell et al., 1992; Saint-Germain and Greene, 2009). However, larval galleries dug into the wood by the whitespotted sawyer cause important economic losses to the sawmill industry (Cerezke and Volney, 1995; Saint-Germain and Greene, 2009; Gervais, 2010). Predicting the seasonal progression of larval damage would help forest managers to develop salvage logging plans minimizing economic losses. However, even if the life cycle of the whitespotted sawyer is relatively well known, its seasonal oviposition period has never been determined. Such knowledge is an essential prerequisite for developing a predictive model of the seasonal progression of larval damage into the wood. It would be the starting point of such model and therefore, our first objective was to determine the seasonality of egg deposition of the whitespotted sawyer and other longhorned beetles also present in different wildfires. We were also interested to improve forecasting of woodborer damage in new burns. Thus, our second objective was to determine if the abundance of adult beetles captured in trunk-window traps could be used to forecast longhorned beetle larval density and thus of potential damage.
Materials and methods
Study areas. We studied five recently (<1 week after the burn area was made accessible by the chief of security) and severely burned sites in Québec, Canada (Fig. 1-1). In each bum, we selected one mature black spruce stand; trees were 15 to 20 cm in
diameter at breast height (DBH). In 2009, we studied the Chapais (49°55'N; 75°14'W) and Assinica (50°8'N; 75°7'W) bums located in the Chibougamau region. Theses bums covered respectively 78 and 665 ha, were ignited on June 18 and 24 respectively and burned until June 23 and July 9. In 2010, three bums were studied. The first one, named the Rivière Serpent burn (49°42'N, 71°20'W) covered 1,654 ha and burned during three weeks, from May 16 to June 5. The second one, the Lac Mondonac bum (47°21' N, 74°00' W), covered 5,865 ha and burned between May 24 and June 9. The last one, the Lac Smokey burn (48°15' N, 73°31' W), was ignited on May 26 and burned until the beginning of September, affecting over 107,000 ha. All these bums were ignited by lightning.
Insect sampling. We studied the adult seasonal activity and seasonal egg deposition of longhorned beetles on severely burned black spruce trees (bark completely carbonized and needles/twigs no longer present) in all sites except in the Assinica bum in which we sampled adults but only estimated the post-winter survival of larvae. Fifty black spruce trees were selected in each bum. Trees were protected from insect egg deposition by surrounding the trunk with a thick black tarpaper between 0.5 and 2 m above ground. Afterwards, at weekly intervals, the tarpaper was removed from five trees randomly selected in each bum to expose these trees to longhorned beetle egg deposition. At each visit, tarpaper of five new trees was removed and the five trees previously exposed were cut to collect a 1-m bole section. These bole sections were numbered and brought back to the Laurentian Forestry Centre, Natural Resources Canada in Québec City, where they were kept outside in a screened protected shelter. Before applying the tarpaper protection, we cut five control trees and collected bole sections in the same way, to ensure that no longhorned beetles had already laid eggs on trees. The detection of longhorned beetle eggs in bark is a difficult and tedious task while larvae are much easier to find under bark. Therefore, we waited at least 3 weeks to allow eggs to hatch and larvae to establish under bark. Larvae were thus used as a proxy of egg deposition. To facilitate bole handling, each 1-m log was cut in two 50-cm logs before processing in the lab. Afterwards, logs were debarked and peeled and longhorned beetle larvae were collected and preserved in 70 % ethyl alcohol until identification. In the Assinica bum, we collected 4 severely burned logs in late May 2010, one year after fire. These logs were debarked and peeled to collect larvae in order to
estimate egg density resulting from cumulative egg deposition done during summer 2009. To monitor the flying activity of adult longhorned beetles, 5 trunk-window traps (TWTs) (Kaila, 1993) were placed in each bum. These traps were made with a 10 x 40 cm plexiglas panel attached to the trunk of burned black spruce trees at DBH. A funnel (diameter: 16 cm, height: 10 cm) placed below the panel directed insects into a collecting recipient (plastic bottle of 500 ml) containing 200 ml of a 40 % ethyl alcohol solution + 5 % vinegar (to keep specimens soft) for killing and preserving insects.
Statistical analysis. Longhorned beetle abundance, and resulting damage, may vary between bums according to several environmental variables (Saint-Germain, 2004a). We pooled weekly abundances of longhorned beetle adults for each trap and used one-way ANOVAs in order to compare their overall abundances between bums. We also used one-way ANOVAs to compare their abundance at the species level. Analyses were made using the GLM procedure of SAS (SAS Institute version 9.2) and the least significant difference (LSD) test was used to compare differences between bums for each significant ANOVA. Linear regressions were used to verify if the abundances of the three most common longhorned beetle species were linked, which would indicate that they respond similarly in various bums. Analyses were performed using the PROC REG procedure of SAS (SAS Institute version 9.2).
In order to determine if the number of whitespotted sawyer adults caught in trunk-window traps in burned sites could be used to predict larval density in trees and thus potential damage, linear regressions were performed while considering different subsets of captures: the first week after completing trap installation, the first two weeks, the first four weeks, or the entire season. For the Assinica bum, since we collected larvae in logs only the following spring, we applied an average mortality of 83.7 % obtained from Rose (1957) for the period ranging between egg deposition and larval entrance into the wood. The PROC REG procedure of SAS was also used in these analyses. Data were transformed (log x+1) to normalize distributions and stabilize variances when necessary.
Results
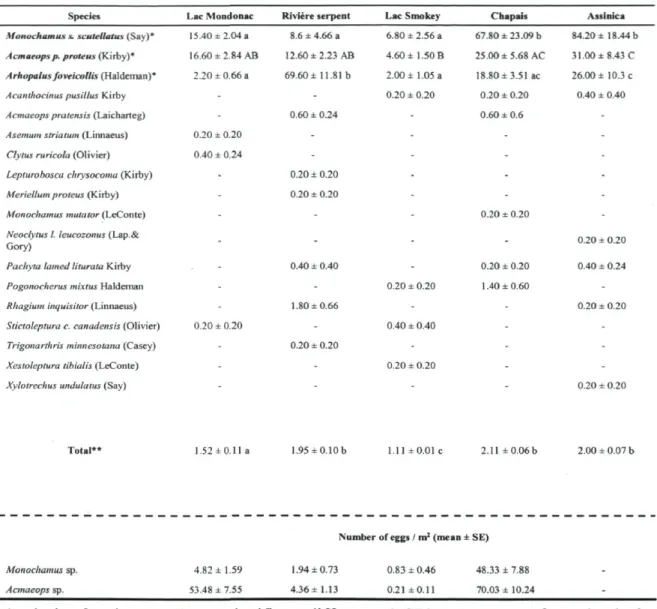
Longhorned beetle diversity and abundance. A total of 2003 adult longhorned beetles distributed among 18 species were captured in the five burns, among which M. s. scutellatus, A. p. proteus and A. foveicollis were the most abundant species (Table 1-1). Overall, a significant difference in longhorned beetle's abundance was observed between bums (F4)2o= 21.01, p < 0.0001) (Table 1-1). The northern bums (Rivière Serpent, Chapais and Assinica) showed a higher abundance of longhorned beetles than those located further south (Lac Mondonac and Lac Smokey), but strong regional differences were noticed in the relative importance of species. The abundance of M s. scutellatus was significantly higher in Chapais and Assinica (F4> 2o = 15.32, p< 0.0001) than in the three other bums while the abundance of A. foveicollis was higher in the Rivière Serpent bum (F4j 20 = 25.92, p< 0.0001). No difference in abundance of A. p. proteus was observed between Assinica and Chapais bums but in the last case, the abundance was similar with Lac Mondonac and Rivière Serpent bums (F4i 20= 7.02, p = 0.0011). The total abundance of M s. scutellatus and A. p. proteus among the five bums followed a similar pattern being most abundant in the Assinica and Chapais bums followed by the Lac Mondonac, Riviere Serpent and Lac Smokey bums. Indeed, a significant relationship was found between their respective abundances in the five bums (r2 = 0.87, p = 0.0197) (Figure 1-2). No relationship was found between M. s. scutellatus or A. p. proteus and A. foveicollis, but higher abundance of this last species was observed in northern bums (Table 1-1). We found a total of 3,563 longhorned beetle larvae with the debarking/peeling method used as a proxy of egg deposition. Larvae belonging to M. s. scutellatus and A. p. proteus were the dominant species found in logs, accounting for 33.8 and 61.2 % respectively of the overall abundance (Table 1-1). The proportion of the remaining larvae (5 %) could not be identified at the sepecies level.
Adult seasonality and egg deposition. Seasonal egg deposition of M s. scutellatus was closely linked to adult activity patterns (Figures 1-3 and 1-4). This differs from A. p. proteus which deposited eggs several weeks after the peak of adult seasonal activity (Figures 1-3 and 1-4). In southern burns (Lac Mondonac and Lac Smokey), M. s.
scutellatus adults were captured up to the end of July while in the north (the three other bums), they were found throughout August and even in early September. However, even if adult abundance increased in late summer 2009 (Figure 1-4), eggs were then deposited at a lower rate than expected on the basis of adult captures. Egg deposition of both M. s. scutellatus and A. p. proteus lasted about two months and was particularly high in the Chapais bum (2009) and at Lac Mondonac (2010), particularly for A. p. proteus. Adult abundance reached the highest levels when bums were colonized rapidly after the end of the fire. Few beetles of both species were caught and few eggs were deposited in the Rivière Serpent and the Lac Smokey bums (Figure 1-3).
Seasonal activity patterns of A. foveicollis were clearly defined only in northern bums; Chapais and Assinica in 2009 (Figure 1-4) and Rivière Serpent in 2010 (Figure 1-3). This last bum was the earliest studied and the first A. foveicollis adults were captured only three weeks after the end of the fire (June 20). Adult activity peaked by July 10 in that bum and its activity continued until August 20, thus covering two months. In the Chapais and Assinica bums, which occurred in late June 2009, the first adults were caught rapidly after the fire, increased until late July and peaked in mid-August. This seasonal pattern doesn't seem to be linked with seasonal temperatures. In southern bums (Lac Mondonac and Lac Smokey), A. foveicollis was uncommon and scattered between mid-June and late-July.
Relation between M. s. scutellatus adult abundance and larval density. A significant relationship was found between adult abundance of M s. scutellatus in trunk-window traps and resulting larval density, while considering short term colonization (one or two weeks after trap installation) (1 week: 1^=0.90, p=0.0134; 2 weeks: r2=0.93, p=0.0074), as well as for a longer period (4 week: 1^=0.68, p=0.0856) or the entire season (r2=0.88, p=0.0173) (Figure 1-5).
Discussion
Our results suggest that a short delay between the fire event and the seasonal occurrence of longhorned beetles increase the risks of heavy colonization that may result in severe
woodborer damage which compromise the benefits of salvage logging for lumber products. The two 2009 bums occurred in late June, when most longhorned beetle adults were already active as shown by their rapid colonization of these bums. Few data exist on adult emergence of longhorned beetles in field conditions but Simpson (1951) reported that most M. s. scutellatus adults emerged from infested logs or killed trees between June 20 to July 10 in central New Brunswick. Gardiner (1956) observed that A. p. proteus adults were active from mid-June to mid-September. This is consistent with our measurements of adult activity in these bums. Moreover, these two burns were the most heavily colonized by both M. s. scutellatus and A. p. proteus. This adds accuracy to the statement of Cerezke and Volney (1995) that timber burned during spring or early summer were more attractive to M. s. scutellatus adults than timber burned in late summer or fall. Our results strongly suggest that the most heavily affected stands would be those that burned in early summer.
In addition to the seasonal period of the bum, its size has been suggested to influence colonization and resulting population levels of M. s. scutellatus (Cerezke and Volney, 1995). The 2009 burns were also the smallest ones in our study and occurred during a season where burns were not abundant in Quebec with only 17,948 ha affected compared with an annual average of 116,424 ha (2002 to 2011 ; SOPFEU). Different cues can be used by longhorned beetles for locating bums (Saint-Germain et al., 2004a; Boulanger, 2010). Burned trees are charred by fire and it has been reported that they release high amounts of ethanol (Kelsey and Joseph, 2003). Monochamus s. scutellatus and A. p. proteus are known to respond to host volatiles such as ethanol and monoterpenes emitted by dead trees (Chénier and Philogène, 1989; Allison et al., 2004; de Groot and Nott, 2004). Therefore, the 2009 conditions might have been optimal for longhorned beetle colonization as volatiles were released during short periods of time and over small areas, thus providing important cues for beetles. Moreover, as these fires occurred in late June, beetles were already active in surrounding forests which are also better habitat sources for longhorned beetles because of the less intensive management of northern forests.
Other factors may have influenced longhorned beetle populations. For instance, the higher abundance of A. foveicollis in northern bums might be linked to the greater abundance of
jack pine as this insect is closely associated with this tree species (Berthiaume et al., 2010). Arhopalus foveicollis has also been reported in other bums in northwestern Quebec
(Boulanger and Sirois, 2007; Boulanger et al., 2010) but was absent from the Parc des Grands-Jardins bum, which was located much southern (Saint-Germain, 2004a). Thus, it might be linked to the colder climate in the north, to the less intensive forest management, to the higher recurrence of forest fires, to the greater abundance of jack pine or to a combination of these factors. Also, since higher amounts of volatiles could result in a greater attraction to severely burned habitats (Kelsey and Joseph, 1999; Pureswaran et al., 2004), differences in bum severity between bums could also influence abundance of longhorned beetles.
The 2010 bums occurred earlier during the season and the abundance of longhorned beetles was lower (except for A. foveicollis in the Rivière Serpent bum) and the delay between fire events and beetle colonization was longer than for the 2009 bums. A decreasing concentration of volatile compounds as time elapses could explain the lower colonization in the 2010 bums. For instance, only few adults were captured and larvae were nearly absent from logs at the Lac Smokey bum. This was the largest bum of our study and it burned from late May to early September. The map showing the seasonal bum progression indicates that the wind changed direction from a northwest-southeast axis at the beginning of the fire to a southwest-northeast axis after 3 weeks (Figure 1-6). Our stand was located in the western part of the bum area and winds thus transported smoke and volatiles far to the northeast. As this area burned all over the season, it is possible that these cues may have not been of any use to longhorned beetles in locating the area under study.
Even if A. foveicollis adults were abundant in traps, it was not possible to determine oviposition pattern for this species because no larva were found when peeling logs. Yanega (1996) noticed that larvae of A. foveicollis feed at the base of dead pine and spruce trees. According to Gardiner (1970), species belonging to the subfamily Aseminae like A. foveicollis are known to feed on stumps and roots. Egg deposition by M. s. scutellatus closely followed the pattern of adult activity, except near the end of summer. However, A. p. proteus egg deposition was maintained for several weeks after the adult peak activity.
This suggests that A. p. proteus males are probably very active when they colonize stands, looking for mates. Females would be much less active flyers and probably walk on boles depositing eggs (Cerezke, 1977) during following weeks. Spreading egg deposition over several weeks might be a strategy to lower risks of direct competition with other larger longhorned beetles or to have longer time for finding suitable hosts in the green forest. This spread of egg deposition could be interesting on a conservation basis as it may provide a continuous access to food for species such as the black-backed woodpecker {Picoides arcticus Swainson), an abundant bird in burned forests (Nappi et al., 2004, 2010).
Recently burned forests constitute important volumes of timber that are economically valuable for harvesting if woodborer damage remains low. Our results support those of Richmond and Lejeune (1945) that wood burned in early summer (late June) would be more heavily attacked by woodborers during the season it burned. In this respect, our attempt to predict longhorned beetle larva; density, and thus potential damage, on the basis of adult abundance in trunk-window traps provide interesting potential. These traps can be rapidly set up and their potential for predicting longhorned beetle damage should be validated in future years and if so, a predictive model of longhorned beetle damage could then be developed. The egg deposition curves will also serve as a starting point in modeling the progression of longhorned beetle damage into trees. Such tools would help managers to better determine areas that should be salvaged or not as well as when they should be salvaged.
Acknowledgements
We thank Y. Dubuc, M. Charest, J-F. Bourdon, M. Couture, P. Descôtes, J.-P. Gendron-Bouchard and E. Godbout for technical assistance. We are grateful to Marcel Trudel (Société de protection des forêts contre le feu (SOPFEU)) for his support and information sharing about wildfires. Financial support was provided to the iFor research Consortium by the Natural Science and Engineering Research Council of Canada, the Ministère des Ressources Naturelles et de la Faune du Québec, the Conseil de l'Industrie Forestière du Québec, the Canadian Forest Service, and the Société de Protection des Forêts contre les Insectes et les Maladies du Québec.
References cited
Allison, J. D., Borden, J. H. and Seybold, S. J. 2004. A review of the chemical ecology of the Cerambycidae (Coleoptera). Chemoecology 14, 123-150.
Bergeron, Y., Gauthier, S., Kafka, V., Lefort, P. and Lessieur, D. 2001. Natural fire frequency for the eastern Canadian boreal forest: consequences for sustainable forestry. Canadian Journal of Forest Research 31, 384-391.
Berthiaume, R., Hébert, C , Boucher, J., Bélanger, S., Légarê, J. P. and Bauce, É. 2010. Colonisation et performances des longicornes dans les brûlis. Rapport interne. Ministère des Ressources naturelles du Québec, Canada. 28 p.
Boulanger, Y. 2010. Colonisation initiale des coléoptères saproxyliques et décomposition des débris ligneux grossiers d'épinette noire après feu en milieu boréal. Thèse de doctorat, Université du Québec à Montréal, 247 p.
Boulanger, Y. and Sirois, L. 2007. Postfire succession of saproxylic arthropods, with emphasis on Coleoptera, in the north boreal forest of Quebec. Environmental Entomology 36, 128-141.
Cerezke, H. F. 1977. Characteristics of damage in tree-length white spruce logs caused by the white-spotted sawyer, Monochamus scutellatus. Canadian Journal of Forest
Research 7, 232-240.
Cerezke, H. F. and Volney, W. J. A. 1995. Forest insect pests in the northwest region. In.
Armstrong, J.A., Ives, W.G.H. (Eds.) Natural Resources Canada, Canadian Forest Service, Science and Sustainable. Development Directorate, Ottawa, 59-72.
Chénier, J. V. R. and Philogène, B. J. R. 1989. Field responses of certain forest Coleoptera to conifer monoterpenes and ethanol. Journal of Chemical Ecology 15, 1729-1745. de Groot, P. and Nott, R. W. 2004. Response of the whitespotted sawyer beetle,
Monochamus s. scutellatus, and associated woodborers to pheromones of some Ips
and Dendroctonus bark beetles. Journal of Applied Entomology 128,483-487. Dyer, L. J. and Seabrook, W. D. 1978. Some aspects of oviposition site selection in
Monochamus notatus and M. scutellatus (Coleoptera: Cerambycidae). Journal of
Chemical Ecology 4, 199-210.
Evans, W. G. 1966. Morphology of the infrared sense organs of Melanophila acuminata
(Buprestidae: Coleoptera). Annals of the Entomological Society of America 59, 873-877.
Gardiner, L. M. 1956. Studies of wood-boring beetles (Cerambycidae). Int. Rept. 1956-4, Forest Insect Lab. Sault Ste Marie.
Gardiner, L. M. 1957. Deterioration of fire-killed pine in Ontario and the causal wood-boring beetles. Canadian Entomologist 89,241-263.
Gardiner, L. M. 1958. Studies of wood-boring beetles (Cerambycidae). Int. Rept. 1956-4, Forest Insect Lab. Sault Ste Marie.
Gardiner, L. M. 1970. Immature stages and habits of Spondylis upiformis Mannerheim (Coleoptera: Cerambycidae). Pan-Pacific Entomology 46, 33-36.
Gervais, D. J. 2010. Effects of post-fire salvage logging on the biodiversity of wood-boring beetles and the distribution of wood-boring beetle damage in fire-killed black
spruce {Picea mariana). Master thesis. Department of Biology. University of Concordia. 81 p.
Johnson, E. A. 1992. Fire and vegetation dynamics: studies from the North American boreal forest. Cambridge University Presse, Cambridge, U.K.
Kaila, L. 1993. A new method for collecting quantitative samples of insects associated with decaying wood or wood fungi. Entomologica Fennica 4, 21-23.
Kelsey, R. G. and Joseph, G. 1999. Ethanol and ambrosia beetles in Douglas fir logs exposed or protected from rain. Journal of Chemical Ecology 25, 2793-2809.
Kelsey, R. G. and Joseph, G. 2003. Ethanol in ponderosa pine as an indicator of physiological injury from fire and its relationship to secondary beetles. Canadian Journal of Forest Research 33, 870-884.
Lowell, E. C , Willits, S. A. and Krahmer, R. L. 1992. Deterioration of killed and fire-damaged timber in the Western United States. Department of Agriculture, Forest Service, Pacific Northwest Research Station Gen. Tech. Rep. PNW-GTR-292. Portland.
Morley, P. M. 1939. Time of cut as a factor influencing infestation of coniferous logs. Canadian Entomologist 71, 243-248.
Nappi, A., Drapeau, P. and Savard, J. P. L. 2004. Salvage logging after wildfire in the boreal forest - Is it becomming a hot issue for wildlife? Forestry Chronicle 80, 67-74.
Nappi, A., Drapeau, P., Saint-Germain, M. and Angers, V. A. 2010. Effect of fire severity on long-term occupancy of burned boreal conifer forests by saproxylic insects and wood-foraging birds. International Journal of Wildland Fire 19, 500-511.
Parmelee, F. T. 1941. Long-homed and flat-headed borers attacking fire-killed coniferous timber in Michigan. Journal of Economic Entomology 34, 377-380.
Pastor, J. and Mladenoff, D. J. 1992. The southern boreal-northern hardwood forest border. In: H. H. Shugart, R.L.e.G.B.B. (Ed.), A system analysis of the global boreal forest, Cambridge, 144-169.
Payette, S., Morneau, C , Sirois, L. and Mireille, D. 1989. Recent fire history of the Northern Quebec Biomes. Ecology 70, 656-673.
Peddle, S. M. 2000. Host selection, oviposition behaviour, and inter- and intra-specific competition in the white-spotted pin sawyer beetle, Monochamus scutellatus (Say) (Coleoptera:Cerambycidae). Master thesis. Faculty of Forestry. University of Toronto, 95 p.
Purdon, M., Noël, J., Nappi, A., Drapeau, P., Harvey, B., Brais, S., Bergeron, Y., Gauthier, S. and Greene, D. 2002. The impact of salvage-logging after wildfire in the boreal forest: lessons from the Abitibi NSERC-UQAT-UQAM Industrial Chair in Sustainable Forest Management, SFM Chair, 4th Research Note. 8 p.
Pureswaran, D. S., Gries, R. and Borden, J. H. 2004. Quantitative variation in monoterpenes in four species of conifers. Biochemical Systematics and Ecology 32,
1109-1136.
Richmond, H. A. and Lejeune, R. R. 1945. The deterioration of fire-killed white spruce by wood-boring insects in northern Saskatchewan. Forestry Chronicle 21 168-192. Rose, A. H. 1957. Some notes on the biology of Monochamus sacutellatus (Say)
(Coleoptera: Cerambycidae). Canadian Entomologist 89, 547-553.
Ross, D. A. 1960. Damage by long-homed wood borers in fire-killed spruce, central British Columbia. Forestry Chronicle 36, 355-361.
Saint-Germain, M., Drapeau, P. and Hebert, C. 2004a. Landscape-scale habitat selection patterns of Monochamus scutellatus (Coleoptera: Cerambycidae) in a recently burned black spruce forest. Environmental Entomology 33,1703-1710.
Saint-Germain, M., Drapeau, P. and Hebert, C. 2004b. Xylophagous insect species composition and patterns of substratum use on fire-killed black spruce in central Quebec. Canadian Journal of Forest Research 34, 677-685.
Saint-Germain, M., Drapeau, P. and Hébert, C. 2004c. Comparaison of Coleoptera assemblages from a recently burned and unbumed black cpruce forests of northeasten North America. Biological Conservation 118, 583-592.
Saint-Germain, M. and Greene, D. F. 2009. Salvage logging in the boreal and cordilleran forests of Canada: Integrating industrial and ecological concerns in management plans. Forestry Chronicle 85, 120-134.
Schmitz, H., Schmitz, A. and Bleckmann, H. 2000. A new type of infrared organ in the Australian "fire-beetle" Merimna atrata (Coleoptera: Buprestidae). Naturwissenschaften 87, 542-545.
Schutz, S., Weissbecker, B., Hummel, H. E., Apel, K.-H., Schmitz, H. and Bleckmann, H. 1999. Insect antenna as a smoke detector. Nature 398, 298-299.
Simpson, L. J. 1951. Prevention of damage by borers in softwood logs or fire killed timber. Canadian Department of Agriculture. Divison Forest and Biology. 8 p.
Wikars, L.-O. 1992. Forest fires and insects. Entomologisk Tidskrift 113, 1-11.
Wilson, L. F. 1962. White-spotted sawyer. White-spotted sawyer. U. S. Department of Agriculture. Forest Pest Leaflet. 7 p.
Yanega, D. 1996. Field guide to northeastern longhorned beetles (Coleoptera: Cerambycidae). Illinois Natural History Survey Manual 6, 184 p.
75'VOIN 74'0'0-W
T
73*0'O"W 72'0'0'W
Figure 1-1: Location of the five bums studied in Québec, Canada. A) Chapais B) Assinica C) Rivière Serpent D) Lac Mondonac E) Lac Smokey
Tableau 1-1: Abundance of adult longhorned beetles caught in trunk-window traps and overall egg density (per m2 of bark) in five burned black spruce forests the same year as the fire, in Quebec, Canada.
Number of adults / trap (mean ± SE)
Species Lac Mondonac Rivière serpent Lac Smokey Chapais Assinica Monochamus s. scutellatus (Say)* 15.40 ±2.04 a 8.6 _ 4.66 a 6.80 ± 2.56 a 67.80 ± 23.09 b 84.20 ± 18.44 b Acmaeops p. proteus (Kirby)* 16.60 ± 2.84 AB 12.60 ±2.23 AB 4.60 ± 1.50 B 25.00±5.68AC 31.00 ± 8.43 C Arhopalus foveicollis (Haldeman)* 2.20 ± 0.66 a 69.60 ± 11.81b 2.00 ±1.05 a 18.80 ± 3.51 ac 26.00 ± 10.3 c Acanthocinus pusillus Kirby - - 0.20 ± 0.20 0.20 ± 0.20 0.40 ± 0.40 Acmaeops pratensis (Laicharteg) - 0.60 ± 0.24 - 0.60 ± 0.6
-Asemum striatum (Linnaeus) 0.20 ± 0.20 - - -
-Clytus ruricola (Olivier) 0.40 ± 0.24 - - -
-Lepturobosca chrysocoma (Kirby) - 0.20 ± 0.20 - -
-Meriellum proteus (Kirby) - 0.20 ± 0.20 - -
-Monochamus mutator (LeConte) - - - 0.20 ± 0.20
-Neoclytus I. leucozonus (Lap.&
Gory) - - - - 0.20 ± 0.20
Pachyta lamed liturata Kiiby Pogonocherus mixtus Haldeman Rhagium inquisitor (Linnaeus) Stictoleptura c. canadensis (Olivier) Trigonarthris minnesotana (Casey) Xestoleptura tibialis (LeConte) Xylotrechus undulatus (Say)
0.20 ± 0.20 0.40 ± 0.40 1.80 ±0.66 0.20 ± 0.20 0.20 ± 0.20 0.40 ± 0.40 0.20 ±0.20 0.20 ± 0.20 1.40 ±0.60 0.40 ± 0.24 0.20 ± 0.20 Total** 1.52 ±0.11 a 1.95±0.10b 1.11 ±0.01 c 2.11 ± 0.06b 2.00 ± 0.07 b Monochamus sp. Acmaeops sp. 4.82 ± 1.59 53.48 ± 7.55
Number of eggs / m2 (mean ± SE)
1.94 ±0.73 0.83 ±0.46 48.33 ± 7.88 4.36 ±1.13 0.21 ±0.11 70.03 ± 10.24
* Analysis of variance and least significant difference (LSD) tests were performed only for M. s. scutellatus, A. p. proteus and A. foveicollis and for the overall abundance of longhorned beetles. For each species, means followed by the same letter did not differ significantly (a = 0.05)
o J3 E o. im -■m* a la «S,
I
35 30 25 20 15 10 y = 0.2644x-8.2933 r2 = 0.87 p = 0.0197 10 20 30 40 50 60 70 80 90Number of Monochamus s. scutellatus/trap
Figure 1-2: Relationship between the mean abundances of Monochamus s. scutellatus and Acmaeops p. proteus in trunk-window traps among five bums studied in Québec in 2009 and 2010.