Analysis of radiation induced DNA damage by LC-MS/MS in isolated and cellular DNA
181
0
0
Texte intégral
Figure
+7
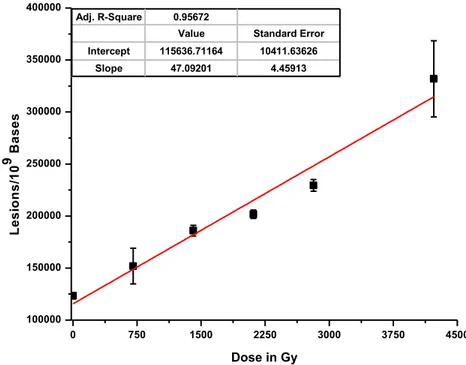
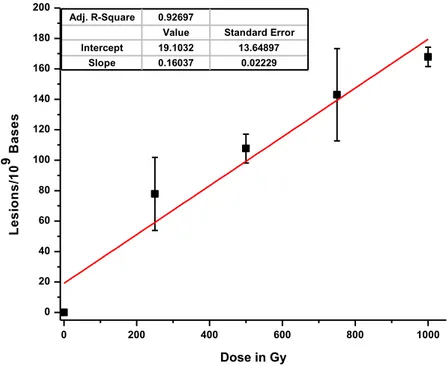
Documents relatifs