Étude de la capacité de discrimination et du patron de consommation de la livrée des forêts (Malacosoma disstria) confrontée à une variation intra et interspécifique de la qualité de ses hôtes alimentaires
100
0
0
Texte intégral
Figure

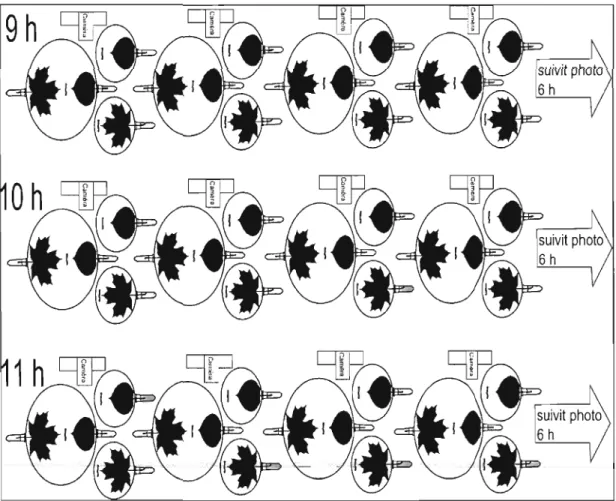
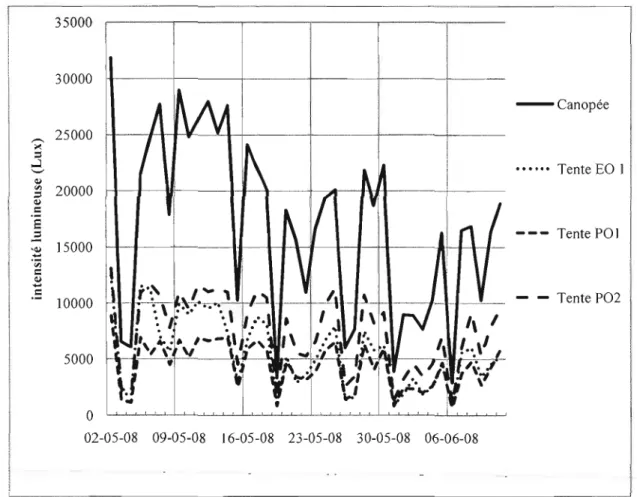
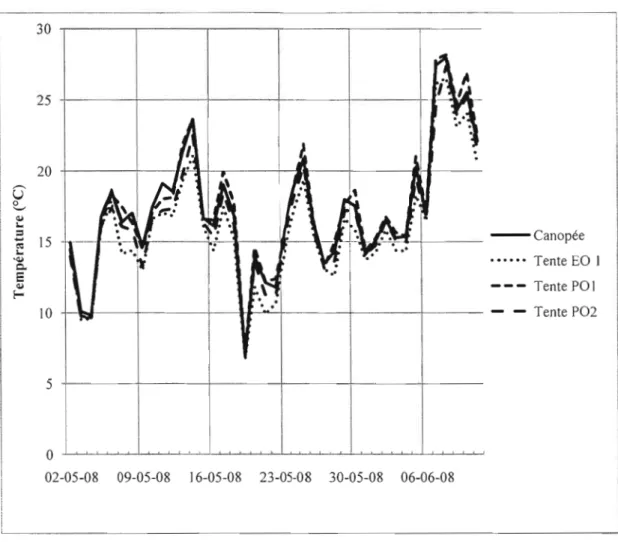
+7
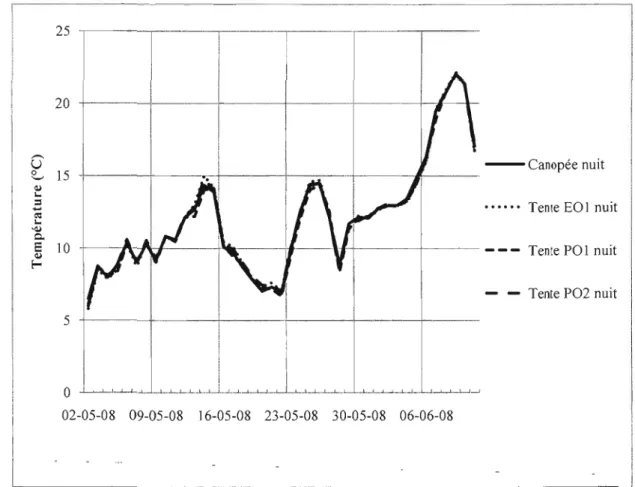


Documents relatifs