Communautés végétales et interactions plante-herbivore :
Comment l’espèce, la qualité nutritive et la répartition spatiale des plantes
environnantes influencent le broutement par les grands herbivores
Thèse
Emilie Champagne
Doctorat en Biologie
Philosophiæ doctor (Ph.D.)
Québec, Canada
© Emilie Champagne, 2017
Communautés végétales et interactions
plante-herbivore
Comment l’espèce, la qualité nutritive et la répartition spatiale
des plantes environnantes influencent le broutement par les
grands herbivores
Thèse
Emilie Champagne
Sous la direction de :
Jean-Pierre Tremblay, directeur de recherche
Steeve D. Côté, codirecteur de recherche
iii
R
ÉSUMÉ
Les interactions trophiques plante-herbivore sont indirectes lorsque la consommation d’une plante par un herbivore est influencée par la présence d’une plante voisine. L’influence des plantes environnantes sur la sélection des herbivores a été décrite par des études sur l’approvisionnement des herbivores, mais aussi par des études sur les effets associatifs entre plantes. À cause de leur évolution parallèle, certains thèmes communs dans les études sur l’approvisionnement des animaux ont eu peu d’échos dans la compréhension des effets associatifs entre les plantes. L’objectif général de cette thèse est de comprendre les effets indirects des communautés végétales sur l’utilisation des plantes par les grands herbivores, dans le contexte des facteurs affectant l’approvisionnement. Premièrement, je me suis intéressée à la distance à laquelle une plante environnante influence la sélection, car l’approvisionnement est un processus spatialement hiérarchique. Grâce à une méta-analyse, je révèle que les plantes environnantes influencent l’utilisation d’autres plantes par les grands herbivores jusqu’à des centaines de mètres carrés. En étudiant les effets associatifs à l’île d’Anticosti (Québec, Canada), je démontre que les plantes environnantes peuvent augmenter ou diminuer le broutement du cerf de Virginie sur le sapin baumier, selon l’échelle considérée. Deuxièmement, j’ai testé l’effet de facteurs susceptibles d’influencer l’approvisionnement des herbivores sur les effets associatifs. Par exemple, la qualité nutritive des plantes environnantes augmentait le broutement sur les sapins. De plus, l’abondance relative des plantes de la communauté pouvait créer des effets associatifs : en Outaouais (Québec, Canada), les pins blancs étaient moins broutés dans les parcelles plus diversifiées où l’abondance relative des espèces préférées était plus faible. Ma thèse met en valeur l’importance d’incorporer une perspective animale, qui tient compte des objectifs et des contraintes qui génèrent les choix des herbivores, à l’étude des effets associatifs. De plus, elle propose des avenues de recherches prometteuses, comme incorporer la variabilité intraspécifique dans les traits fonctionnels des plantes, tel que leur qualité nutritive, et considérer les effets associatifs à plusieurs échelles spatiales. Ces travaux améliorent notre compréhension des interactions trophiques indirectes, en démontrant l’influence de facteurs comme l’abondance des ressources sur ces interactions. De plus, je propose un mécanisme potentiel, soit les comportements d’approvisionnement des herbivores, ce qui ouvre la voie à une généralisation des effets associatifs à différents systèmes.
iv
A
BSTRACT
Plant-herbivore trophic interactions can be indirect, such as when the consumption of a plant by an herbivore is influenced by the presence of a neighbouring plant. The influence of neighbouring plants on herbivores’ selection has been described in studies of foraging but also in studies of associational effects between plants. Because of their parallel evolution, some common theme in the studies of animal foraging has been seldom addressed in the understanding of associational effects among plants. The general objective of this thesis is to understand the indirect effects of vegetation communities on plant use by large herbivores, in the context of factors affecting foraging. First, I investigated at which distance a neighbouring plant can influence herbivores’ selection, because foraging is a spatially hierarchical process. By a meta-analysis, I uncovered that neighbouring plant can influence the use of other plant species by large herbivores up to hundreds square metres. By a study of associational effects on Anticosti Island (Québec, Canada), I also demonstrated that neighbouring plants can increase or decrease browsing by white-tailed deer on balsam fir, depending on the scale considered. Second, I tested the relative contribution on associational effects of factors susceptible to influence foraging by herbivores. For example, the nutritional quality of neighbouring plants increased browsing on firs. Moreover, the relative abundance of plants in the community can generate associational effect: in the Outaouais region (Québec, Canada), white pines were less browsed in diversified plots where the relative abundance of preferred species was lower. My thesis supports the importance of incorporating an animal perspective, which takes into account the objectives and constraints on herbivores foraging choices, in the study of associational effects. Moreover, it offers promising research perspective, like including plant intraspecific variability in functional traits, such as nutritional value, and considering associational effects at multiple spatial scales. This work improves our understanding of indirect trophic interactions, by demonstrating the influence of factors such as resource abundance on these interactions. It also proposes a mechanism, foraging processes of herbivores, which paves the way to a generalization of associational effects in different systems.
v
T
ABLE DES MATIÈRES
Résumé ... iii
Abstract ... iv
Table des matières ... v
Liste des tableaux ... ix
Liste des figures ... xii
Remerciements ... xviii
Avant-propos ... xxii
CHAPITRE 1 Introduction ... 1
Objectifs de la thèse ... 10
Approche méthodologique ... 10
CHAPITRE 2 Spatial extent of neighbouring plants influences the strength of associational effects on mammal herbivory ... 13
Résumé ... 14
Abstract ... 15
Introduction ... 16
Methods ... 19
Literature review ... 19
Data extraction and effect size computation ... 20
Statistical analyses ... 22
Results ... 24
Discussion ... 28
Acknowledgement ... 32
CHAPITRE 3 Neighbouring plants and perception of predation risk modulate winter browsing by white-tailed deer ... 33
Résumé ... 34
Abstract ... 35
Introduction ... 36
Material and Methods ... 37
Experimental Design ... 39
Data Collection ... 41
vi
Statistical Analyses ... 42
Preference Experiment ... 42
Associational Effects and Predation Risk Experiment ... 42
Results ... 43
Preference Experiment ... 43
Associational Effects Experiment ... 47
Predation Risk Experiment ... 47
Discussion ... 49
Acknowledgments ... 52
CHAPITRE 4 Nutritional quality of neighbouring plants and plant density determine ungulate herbivory ... 54
Résumé ... 55
Abstract ... 56
Introduction ... 57
Material and methods ... 59
Study area ... 59
Sampling Design and Data Collection ... 59
Chemical Analyses and Near Infrared Reflectance Spectroscopy Calibration ... 60
Statistical Analyses ... 61
Results ... 64
Browsing on Focal Fir ... 64
Browsing on Fir at the Patch Scale ... 69
Discussion ... 70
Acknowledgements ... 73
CHAPITRE 5 Spatial correlations between browsing on balsam fir by white-tailed deer and the nutritional value of neighbouring winter forage ... 75
Résumé ... 76
Abstract ... 77
Introduction ... 78
Methods ... 80
Study area and data collection ... 80
Spatial analyses... 84
vii
Correlations between browsing and neighbourhood characteristics ... 85
Influence of landscape configuration ... 86
Results ... 87
Distribution of fir browsing ... 87
Correlations between browsing and neighbourhood characteristics ... 87
Influence of landscape configuration ... 91
Discussion ... 91
Acknowledgements ... 95
CHAPITRE 6 Forage type and abundance, not diversity per se, influence winter resource selection by white-tailed deer ... 96
Résumé ... 97
Abstract ... 98
Introduction ... 99
Material and Methods ... 101
Community Diversity and Composition ... 102
Resource Selectivity in Deer ... 102
Statistical Analyses ... 103
Results ... 104
Community composition ... 104
Browsing on All Species ... 106
Selectivity ... 108
Browsing on White Pines ... 108
Discussion ... 114
Acknowledgements ... 116
CHAPITRE 7 Conclusion générale ... 118
Pertinence d’étudier les effets associatifs dans un cadre d’approvisionnement ... 120
Intégrer la variabilité intraspécifique des plantes ... 123
Intégrer un cadre spatial ... 126
Les effets associatifs en forêts boréale et mixte ... 128
Applications des résultats à la gestion ... 129
Applications spécifiques à l’île d’Anticosti ... 129
Applications à la gestion en forêts boréales et mixtes ... 131
viii
BIBLIOGRAPHIE ... 134
ANNEXE 1 Informations supplémentaires concernant le protocole du chapitre 2 ... 150
ANNEXE 2 Tableau des variables extraites au chapitre 2 ... 154
ANNEXE 3 Détails des analyses statistiques pour le biais de publication et la tendance temporelle du chapitre 2 ... 159
ANNEXE 4 Paramètres de calibration des modèles de spectroscopie en proche infrarouge utilisés aux chapitres 4 et 5 ... 166
ANNEXE 5 Résultats de la sélection de modèles au chapitre 4 ... 169
ANNEXE 6 Figures supplémentaires du chapitre 5 ... 180
ix
L
ISTE DES TABLEAUX
Table 3.1 Results of the general linear models (GLM) and the general linear mixed models (GLMM) for the associational effects experiment and the predation risk experiment for white-tailed deer browsing in Outaouais (2014) on white pine and on Anticosti Island (2015) on balsam fir. ... 45 Table 4.1 Impacts of balsam fir nutritional value, neighbouring stem abundance and
neighbouring stem nutritional value on the number of shoots browsed on a) the focal fir and b) on all firs in 4 m2 plots, on Anticosti island (Québec, Canada). In each row, we present the estimates from model averaging with 95%CI for each independent variable in the sets of candidate models. Each column corresponds to a different set of candidate models, including the nutritional value (N: %Nitrogen, IVTD: % In vitro true digestibility on dry matter basis) either of the focal fir (n = 124) or of one of the three neighbouring species (fir: n = 104, birch: n = 78 or white spruce: n = 84) and neighbour abundance variables (number of fir, birch, spruce spp. stems, the total number of stems, and the number of available and browsed birch shoots).We used generalized linear models with a negative binomial distribution, including the log-transformed number of shoots available as an offset. In b), all models also included the distance between the plot and the nearest residual forest patch.The 95% CI (confidence interval) of estimates in bold do not include 0. ... 65 Table 5.1 Descriptive statistics for plot characteristics and variables measured in 4 m2 and
40 m2 concentric sub-plots on Anticosti Island (Québec, Canada). We collected the data for the three species (balsam fir, paper birch and white spruce) either in 2013 or in 2014. We calculated browsing rate (%) by white-tailed deer as the number of shoots browsed/number of shoots available in each 4 m2 plot. We collected all the other variables in the 40 m2 plot. We evaluated fibre content (%, NDF, ADF, ADL), in vitro dry matter digestibility ( IVTDDM) and nitrogen content (% N) using bulk samples collected over 3-10 stems per plot. Distances are expressed in metres. ... 88 Table 6.1 Effect of community diversity and composition on winter browse consumption
(number of shoots browsed) by white-tailed deer. We tallied 102 plots located in a deer wintering area and partitioned in 15 transects in western Québec (Canada). We used generalized linear mixed models with a negative binomial distribution, with the transect as a random variable. All models included resource availability (number of available shoots). We centred all explanatory variables, except the PCA values that were already centred. P-values in bold are significant at α = 0.05 and statistical trends at α = 0.10 are in italics. ... 107 Table 6.2 Effect of community diversity and composition on the selectivity of white-tailed
deer, as estimated by the overall selectivity index (OSI). We tallied 102 plots located in a deer wintering area and partitioned in 15 transects in western Québec (Canada). We used general linear mixed models, with the transect as a random variable and a log transformation on the response variable to reach model assumptions. All models included resource availability (number of available shoots). We centred all explanatory variables, except the PCA values that were already centred. P-values in bold are significant at α = 0.05 and statistical trends at α = 0.10 are in italics. ... 110
x
Table 6.3 Effect of community diversity and composition on winter consumption (number of shoots browsed) of the focal white pine by white-tailed deer. We tallied 102 plots located in a deer wintering area and partitioned in 15 transects in western Québec (Canada). We used generalized linear mixed models with a negative binomial distribution, and the transect as a random variable. All models included resource availability (number of available shoots of all species and number of pine shoots available). We included as an offset the log-transformed number of shoots available on the focal pine. We centred all explanatory variables, except the PCA values that were already centred. P-values in bold are significant at α = 0.05 and statistical trends at α = 0.10 are in italics. ... 111 Table A2.1 Variables extracted from 46 accepted articles reporting results about
damage/survival of a focal plant to herbivory in presence/absence of a neighbouring plant, with their coding categories and number of effect sizes in each category. ... 155 Table A4.1 Summary information of modified partial least squares regression models
predicting fibre content on dry matter basis (NDF, ADF, ADL), in vitro true digestibility on dry matter basis (IVTDDM) and nitrogen content, all expressed as percentages using Near infrared reflectance spectra (NIRS). The r2 value refers to the regression between laboratory and predicted values of the independent validation set. All regression models include values from different tree species: balsam fir (Abies balsamea), white spruce (Picea glauca), black spruce (Picea mariana) and paper birch (Betula papyrifera). The wavelength range used is 1100-2498.2 nm for all samples. ... 167 Table A5.1 Correlation matrix between neighbouring plant abundance and nutritional value
variables used in the sets of candidate models. Values reported are R2 and those above 0.5 are in bold. Nutritional values for different species are never included in the same model or in the same set of candidate models; thus we do not report correlation between those variables. ... 172 Table A5.2 Results of model selection for the effects of neighbour abundance considering
focal fir nutritional value on the number of shoots browsed on the focal fir. ... 173 Table A5.3 Results of model selection for the effects of neighbour abundance considering
neighbouring firs nutritional value on the number of shoots browsed on the focal fir. . 174 Table A5.4 Results of model selection for the effects of neighbour abundance considering
neighbouring birches nutritional value on the number of shoots browsed on the focal fir. ... 175 Table A5.5 Results of model selection for the effects of neighbour abundance considering
neighbouring white spruces nutritional value on the number of shoots browsed on the focal fir. ... 176 Table A5.6 Results of model selection for the effects of neighbour abundance considering
all firs nutritional value on the number of shoots browsed on all firs in the plot. ... 177 Table A5.7 Results of model selection for the effects of neighbour abundance considering
neighbouring birches nutritional value on the number of shoots browsed on all firs in the plot. ... 178
xi
Table A5.8 Results of model selection for the effects of neighbour abundance considering neighbouring white spruces nutritional value on the number of shoots browsed on all firs in the plot. ... 179
xii
L
ISTE DES FIGURES
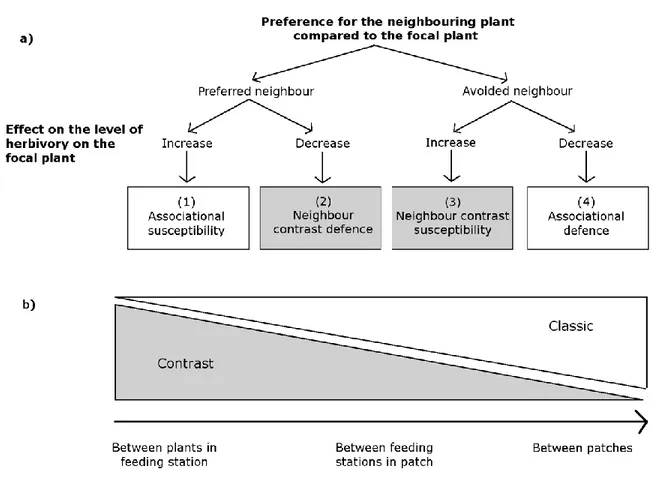
Figure 1.1 Organigramme des quatre types d’effets associatifs. Les effets associatifs se différencient par la préférence de l’herbivore pour la plante environnante, relativement à la plante d’intérêt. Les plantes environnantes, qu’elles soient relativement préférées ou évitées, peuvent augmenter ou diminuer la consommation de la plante d’intérêt. Dans cette thèse, j’utilise la terminologie de Bergvall et al (2006), seule à introduire les effets par contrastes. ... 4 Figure 2.1 (a) Flowchart of the type of associational effects affecting the level of herbivory
on the focal plant based on the preference of the herbivore for the neighbouring plants versus the focal plant (first level of the flowchart) and on the direction of the association (second level). “Classic” types of effects (associational susceptibility and defence) are in white boxes while “contrast” types (neighbour contrast susceptibility and defence) are in grey boxes. (b) Predictions about how the “classic” (white) and “contrast” (grey) associational effects should vary in strength with spatial scale according to Bergvall et al (2006) framework. Scales suggested on the x-axes are suggestions not representing exactly where the type of associational effects are expected to occur. ... 17 Figure 2.2 Summary of difference in damage/survival with and without a neighbouring
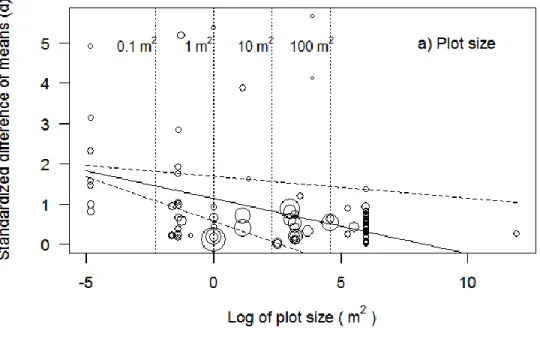
plant (d, standardized difference of means) separated by the independent variable levels tested, with 95% CI and I2, the percentage of total variability due to heterogeneity among d’s. A higher d indicates a higher associational effect of the neighboring plant on the focal plant’s herbivory level. Numbers to the right of the data points are the number of effect sizes in each summary effect. We used a meta-analysis mixed model to test the impact of variables on the standardized difference of means. ... 25 Figure 2.3 Relationship between associational effects and two different indicators of spatial
scale: (a) decrease in the difference in damage/survival with and without a neighbouring plant (standardized difference of means) according to plot size (m2); (b) associational effect limited to the first 10 cm between the focal plant and its neighbours. For each figure, the size of each point indicates the weight of each effect size in the meta-analysis mixed effect model, calculated with the inverse-variance method. Vertical lines and numbers above correspond to untransformed values of plot size (m2). Regression line results from a meta-analysis mixed model and dotted lines represent predicted values with 95% CI. ... 27 Figure 3.1 a) Study areas (black dot: Outaouais, black star: Anticosti Island). Deer densities
were estimated in 2006 in Outaouais (Ministère des Forêts, de la Faune et des Parcs, unpubl. data ) and on Anticosti Island (Rochette and Gingras 2007). Tree species in each bioclimatic domain are reported in Saucier et al. 2009. Species arrangements (view from above) used for b) the preference experiment on white-tailed deer (Odocoileus
virginianus) winter browsing and c) associational effects and predation risk experiments.
Black rectangles are stations, placed on existing deer snow trails. Plots inside the stations are separated by 2 m. Stations are independent, and located at least 200 m from another station. Branches are of similar height. The predation risk experiment has the same design as the associational effect experiment but with half of the stations presenting a scent tag soaked in coyote urine for Outaouais and one third each with
xiii
coyote and fisher urine for Anticosti Island. The scent tag is placed in the middle of the station.. ... 38 Figure 3.2 Winter browsing rates (estimated biomass browsed/available) per species in two-choice preference experiments in the Outaouais region (top panels) and on Anticosti Island (bottom panels). Each panel is one of the species arrangements, offering two plant species to white-tailed deer with each line corresponding to a trial (browsing in one station during 24h). For Outaouais, we tested all combinations of white pine, mountain maple and American beech. For Anticosti Island, we tested all combinations of balsam fir, paper birch and white spruce. Data presented are raw values of browsing. ... 44 Figure 3.3 Winter browsing rates by white-tailed deer on focal species (estimated biomass
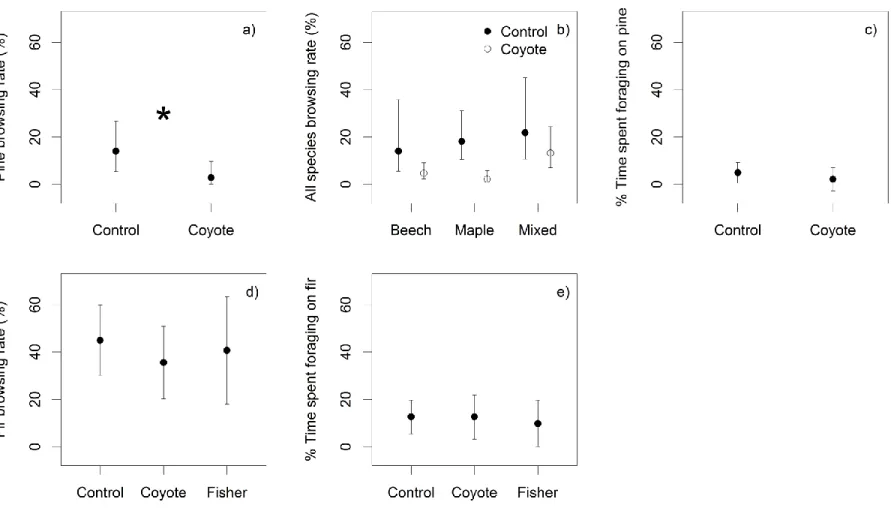
browsed/available; a, c) and % time spent foraging on focal species (b, d) in the associational effects experiment in the Outaouais region (a-b) and on Anticosti Island (c-d). We tested significant differences with GLM (a, c) and GLMM (b, d) and performed a posteriori least square means differences when the treatment was significant (Letters above CI, α = 0.05). Data are model estimates, back transformed when required with 95% CI. ... 48 Figure 3.4 Results of a predation risk experiment in the Outaouais region (a-c) and on
Anticosti Island (d-e) on the focal species browsing rates (estimated biomass browsed/available) by white-tailed deer in winter (a, d), all species browsing rate (b) and % time spent foraging on focal species (c, e) in. We tested significant differences with GLM (a, b, d) and GLMM (c, e) and performed a posteriori least square means differences when the treatment was significant (* above CI, α = 0.05). Data are model estimates, back transformed when required with 95% CI. ... 50 Figure 4.1 Schematic representation of the statistical analyses testing the effects of
neighbour abundance and nutritional values on two responses variables a) the number of shoots browsed on the focal fir and b) the number of shoots browsed on all firs within a plot. Using model selection, we compared 34 candidate models including either neighbour abundance, nutritional value or both. We constructed a different set of candidate models for the nutritional value of each neighbouring species to account for missing values because neighbouring species were not represented in all plots, thus generating distinct data sets. We thus repeated the model selection procedure seven times, for all combination of response variables and neighbours nutritional values, at the exception of shoots browsed on all firs the nutritional value of focal fir. ... 62 Figure 4.2 Variables influencing browsing on focal balsam fir by white-tailed deer in a
management enclosure on Anticosti Island (Québec, Canada), in 2014: a) Interaction between focal fir nitrogen content (minimal, mean and maximal values in dataset represented) with the number of fir stems in the plot; b) nitrogen content of neighbouring firs; c) nitrogen content of neighbouring birches, d) %IVTDDM of neighbouring white spruces. Empty circles are the raw data (not represented for the interaction), the line is the model averaged estimate (generalized linear models) with 95% CI. The number of shoots browsed is the response variable with the log-transformed number of available shoots as an offset, and to present the % of shoots browsed, we divided shoots browsed by available shoots. ... 68
xiv
Figure 4.3 Variables influencing browsing on all balsam firs in plots by white-tailed deer in a management enclosure on Anticosti Island (Québec, Canada), in 2014: a) nitrogen content of neighbouring firs; b) number of fir stems in plot. Empty circles are the raw data, the line is the model averaged estimate (generalized linear models) with 95% CI. For b, estimates from all sets of candidate models were significant and similar, we thus included only the estimate for the set with fir nutritional values. The number of shoots browsed is the response variable with the log-transformed number of available shoots as an offset, and to present the % of shoots browsed, we divided shoots browsed by available shoots. ... 70 Figure 5.1 Enclosed study area on Anticosti Island (Québec, Canada). Grey zones are the
sampling areas located in the cutover patches, with a 15 m buffer from residual forest patches, fences and roads. Plots (n=125) were distributed in the sampling area according to a systematic stratified sampling of non-aligned random points, i.e., four plots were randomly placed in each cell of a 200 m x 200 m grid, with at least 11 m between each to avoid overlap. Each plot consisted of two concentric sub-plots of 4 m2 and 40 m2. Plots located in the sampling area were sampled in 2013 for deer browse (4 m2) and abundance of neighbouring plants (40 m2) and in 2014 for deer browse and nutritional quality (40 m2) ... 81 Figure 5.2 Correlograms for the browsing rate on fir (Abies balsamea, number of shoots
browsed/number of shoots available) in a) 4 m2 plots in 2013 and b) 2014 on Anticosti island (Québec, Canada). Moran’s I was calculated for pairs of plots in distance classes of 50 m. The first bin included distance from 11 m to 50 m, since plots were separated by a minimum of 11 m to prevent overlap. Error bars are 95% confidence intervals. The black dot indicates a statistically significant value with a progressive Bonferonni correction of the α-level, starting with α = 0.05 ... 87 Figure 5.3 Cross-correlograms of the correlation between the browsing rate on balsam fir
(number of shoots browsed/number of shoots available) in 4 m2 plots in 2013 (left column) and 2014 (right column) and a-b) nutritional characteristics of neighbouring fir (Abies balsamea) and c-f) white spruce (Picea glauca). Cross-correlograms between browsing on fir (2013 and 2014) and fir ADL content are almost identical to the relation with fir ADF (g-h) and are presented in Annexe 6. Data were collected on Anticosti island (Québec, Canada). Correlations between each pair of variables were calculated for pairs of plots in distance classes of 50 m and the point is located at the mean value for the class. The first bin included distance from 11 m to 50 m, since plots were separated by a minimum of 11 m to prevent overlap. Black dots indicate statistically significant values with a progressive Bonferonni correction of the α-level, starting with α = 0.05. . 90 Figure 6.1 Ranked Manly’s standardized selection ratios (Bi) for plant species with shoots
available to white-tailed deer. We did not include two species with a number of available shoots under 10, Rhus typhina and Quercus macrocarpa. Ratios add up to 1 and higher values indicate higher selection. The total number of shoots available for a species is indicated between the brackets following the species name. ... 105 Figure 6.2 Interactions between plant community diversity and availability (number of
shoots in the plot) influenced deer consumption of all species present in the plot (a-b) and on the focal pine in the plot (c-d). All data were collected in the sugar
maple-xv
basswood domain, in Western Québec (Canada). Diversity is estimated with the Shannon diversity index (a, d), the Simpson diversity index (b) and the number of species in the plot (richness, c). Explanatory variables are centred and model estimates with 95% CI are presented for relatively low and high shoot availability. ... 109 Figure 6.3 Interaction between species composition and plant availability (number of shoots
in the plot) influenced deer consumption of the focal pine. Positive values of component 3 are associated with the abundance of Dipsacales spp, while negative values are associatied with the abundance of Acer saccharum and Fagus grandifolia. All data were collected in the sugar maple-basswood domain, in Western Québec (Canada). Model estimates with 95% CI are presented for low shoot availability and high shoot availability. ... 113 Figure A1.1 Process followed for the selection of peer-reviewed articles (selection and
exclusion criteria), inspired by the PRISMA flow diagram (Moher et al 2009), and extended with the computation of effect sizes with the number of articles found, rejected and selected. ... 151 Figure A3.1 Funnel plots of the standard error by the standardized difference of means (d)
for the complete model, for each effect size class level (d, r and OR) and for each nature of the experiments (Feeding trials, observational studies, transplantation and removal experiments). Each data point corresponds to one effect size. ... 162 Figure A3.2 Trim and fill funnel plots for the complete model, effect size class d subgroup
and nature of the experiment observational studies subgroup, with the R0 estimator (a, c and e) and the L0 estimator (b, d and f). Each data point corresponds to one effect size. Black dots are real effect sizes and empty dots are the studies missing because of publication bias, as estimated from the trim and fill models. The number of studies potentially missing varies with the estimator used: complete model, R0, n = 7; L0, n = 0; d-class, R0, n = 22; L0, n = 0, observation studies, R0, n = 1, L0 = 1. ... 163 Figure A3.3 Visualization of the cumulative meta-analysis. A mean standardized difference
of means (d, [95% CI]) for each study used in the analysis is added in turn, following its year of publication, to the summary effect. ... 164 Figure A5.1 Abundance of woody plants in a management enclosure on Anticosti Island
(Québec, Canada), in 2014. Each bar represents a different plot (n = 125), with the number of balsam fir (Abies balsamea), spruce (white and black spruce, Picea glauca and P. mariana), paper birch (Betula papyrifera) and other species (Populus tremuloides and P. balsamea) stems. Plots are ordered by the total number of stems in the plot. ... 171 Figure A6.1 Moran’s I correlogram for the abundance and concentration of nutritional
characteristics of balsam fir (first column), paper birch (second) and white spruce (third) in 40 m2 plots on Anticosti island (Québec, Canada). Moran’s I were calculated for pairs of plots in distance classes of 50 m and the point is located at the mean value for the class. The first bin included distance from 11 m to 50 m, since plots were separated by a minimum of 11 m to prevent overlap. Error bars are 95% confidence intervals. Black dots indicate statistically significant values with a progressive Bonferonni correction of the α-level, starting with α = 0.05 ... 181
xvi
Figure A6.2 Cross-correlograms of the correlation between browsing rate on balsam fir (number of shoots browsed/number of shoots available) in 4 m2 plots in 2013 (left column) and 2014 (right column) and nutritional attributes of neighbouring white spruce (a-d) and fir (e-h). Data were collected on Anticosti island (Québec, Canada). Correlations between each pair of variables were calculated for pairs of plots in distance classes of 50 m and the point is located at the mean value for the class. The first bin included distance from 11 m to 50 m, since plots were separated by a minimum of 11 m to prevent overlap. Black dots indicate statistically significant values with a progressive Bonferonni correction of the α-level, starting with α = 0.05. ... 182 Figure A6.3 Cross-correlograms of the correlation between browsing rate on balsam fir
(number of shoots browsed/number of shoots available) in 4 m2 plots in 2013 (left column) and 2014 (right column), and the number of paper birches (a-b) and white spruces (c-d) in 40 m2 plots. Data were collected on Anticosti island (Québec, Canada). Correlations between each pair of variables were calculated for pairs of plots in distance classes of 50 m and the point is located at the mean value for the class. The first bin included distance from 11 m to 50 m, since plots were separated by a minimum of 11 m to prevent overlap. Black dots indicate statistically significant values with a progressive Bonferonni correction of the α-level, starting with α = 0.05. ... 183 Figure A7.1 Rank abundance curve of all the woody plant stems in the plots. Abundance is
recorded as the number of stems > 25 cm in height. To create this figure, we used the
rankabundance and rankabunplot functions of the BiodiversityR package in R 3.3.1
(Kindt and Coe 2005, R Core Team 2016). ... 185 Figure A7.2 Bivariate plot of two significant components of the Principal Component
Analysis (PCA) on the composition of plots including the abundance for 31 woody plant species. We tested significance of each axis using the method described in Peres-Neto et al (2005). Each triangle corresponds to a plot (n = 103). Species names indicate relative values of loadings for each species. Species clustered in the center are represented by open circles. ... 187
xvii
Pour Léonie et Amandine, comme preuve que vous pouvez tout accomplir
« A videorecording of herbivores feeding is not the sort of footage that leads to many Trials of Life-type, glossy documentaries, narrated by important
natural historians with English accents » Jonathan Newman
xviii
R
EMERCIEMENTS
Écrire cette section, c’est le début d’un adieu. Peu importe que je côtoie encore ou non les personnes nommées ci-dessous, elles feront bientôt partie d’une époque révolue, d’un souvenir. Je prends donc aujourd’hui un temps d’arrêt pour immortaliser convenablement ceux et celles qui ont contribué à faire de mon doctorat une période stimulante, amusante, stressante, mais ô combien enrichissante !
Mon projet fait partie de la Chaire de recherche industrielle CRSNG en aménagement intégré des ressources de l’île d’Anticosti, financée par le Conseil de recherche en sciences et génie du Canada (CRSNG), Produits Forestiers Anticosti, le gouvernement du Québec, Sépaq Anticosti, Pétrolia, Gestion Solifor et Safari Anticosti. Merci à ces organismes et compagnies d’avoir contribué à la recherche. J’ai personnellement reçu du financement pour mes études et mon stage à l’étranger du CRSNG, d’EnviroNord, du Centre d’Études Nordiques, du Fonds Richard-Bernard et de la Fédération Canadienne des Femmes Diplomées Universitaires (FCFDU). Merci à tous ces organismes de m’avoir fait confiance et particulièrement à la FCFDU dont la mission me tient particulièrement à cœur.
André Dumont (MFFP) et Daniel Fortin, professeur au département de biologie, ont accepté de faire partie du comité qui m’a encadré tout au long de ma thèse. Vos conseils judicieux ont aidé à orienter ma thèse, la menant dans une direction dont je suis fière. Merci. Merci également à ceux qui ont accepté la tâche de réviser une thèse à sept chapitres : Daniel Fortin, Monique Poulin et Nora Underwood.
Je ne sais pas comment remercier Jean-Pierre Tremblay, mon directeur de recherche. Depuis 2008, Jean-Pierre est un mentor pour moi, une personne inspirante qui a à cœur ma réussite, personnelle et professionnelle. Il m’a poussé à aller plus loin, à repousser les limites que je m’imposais par paresse intellectuelle. Il m’a fait rager, parfois, m’a encouragé, toujours. Jean-Pierre, j’avais de la difficulté à envisager de faire un doctorat sans toi. Le trajet accompli me confirme que j’ai fait un bon choix. Merci pour tout. Merci à Steeve, mon codirecteur, de m’avoir apporté un soutien de grande valeur, des orientations différentes et des conseils toujours pertinents.
xix
À cette équipe de direction s’est ajouté un collaborateur imprévu, Ben Moore. Ben, I hope I haven’t traumatized you by imposing myself into your lab. Our conversations contributed to this thesis and improved it. You gave me confidence and inspired me to go further. Thank you so much.
J’ai eu la chance d’être appuyée dans une de mes expériences par Lucie Perroud, une stagiaire tombée du ciel. Devenue analyste de vidéo et coauteure, Lucie a effectué un travail exceptionnel, avec le sourire et ses expressions bien françaises. Merci de m’avoir sauvé des heures d’analyses vidéos et d’avoir mis un peu de toi dans ma thèse. Aussi, merci pour le fromage.
Après plus de 10 ans dans le même département, je ne peux passer sous silence ceux qui, parfois par leur simple présence, ont contribué à rendre mon passage agréable. Pour leur aide et les mots échangés, merci à Josée et Jocelyne. Pour leur soutien au long de mon parcours universitaire et les conversations positives et constructives, merci à Julie Turgeon, Nadia Aubin-Horth, Gilles Gauthier, Line Lapointe et Stéphane Boudreau. Merci à Hélène Crépeau pour le soutien statistique et surtout pour les réponses simples et rapides. Pour le plaisir au travail, merci à Benoît Dumas. Sans oublier les collègues étudiants, avec qui j’ai ri, discuté… merci de votre présence, particulièrement à Pascale Ropars, ma compagne de bureau des derniers mois.
Merci aux piliers du labo, notre soutien technique à tous, Caroline Hins, Julien Hénault-Richard et Sonia de Bellefeuille. Sonia, ta confiance en moi m’a fait un bien immense, merci. Marie-Claude Martin et Michaël Bonin ont également été d’une aide inestimable en labo, comblant mes lacunes techniques et pratiques. Merci à vous deux. J’ai eu droit à une armée d’assistants de terrain et de laboratoire : Aimée Benoist-Chénier, Jonathan Bonin-Bourgault, Mathieu Boudreau, Katherine Charland, Milène Courchesne, Janie Dufresne, Félix Larochelle, Lorraine Lessard, Guillaume Otis-Lagrange, Félix Parent-Paquette, Brenda Salmon Rivera et Damien. Merci pour votre aide et surtout pour les mesures de broutement. Parmi cette armée, je ne peux passer sous silence le plaisir que j’ai eu à travailler avec Milène et Aimée. Vous êtes de belles personnes, lumineuses, joyeuses. Vous m’avez soutenue, bien au-delà de la collecte de données. Merci !
xx
Après des années de travail dans le Nord, dans une bulle particulière, le terrain effectué à Anticosti et en Outaouais m’a rappelé le côté social important de la science. Merci à l’équipe du Ministère des forêts, de la faune et des parcs en Outaouais de m’avoir appuyée dans mes projets : André Dumont, Bruno Beaudoin, qui m’a notamment permis de collecter quelques branches d’arbres sur sa terre et Stéphan Grondin, qui a sauvé les parcelles de l’Outaouais en me fournissant des informations précieuses. À l’île, j’ai redécouvert une communauté extraordinaire. Merci à Johanne Labonté et à Alex pour les données sur l’île. Merci à Joël, pour son aide technique donnée avec intelligence et humour. Merci à Aimée et Stefan, de m’avoir fait sentir chez moi, notamment par vos services de fournisseurs d’internet. Merci à Stéphanie, de m’avoir inclus à tes activités. Merci à Jean-François Boudreault de m’avoir enseigné la conduite en motoneige. Merci à Alex et Danièle, pour la bonne nourriture et les belles conversations. À tous ces gens et à Sébastien, Bianca, les pilotes ainsi que ceux que j’oublie, merci pour les soirées arrosées et festives.
Mon labo… comment vous rendre justice ? J’ai tellement peur d’oublier quelqu’un et de passer sous silence votre contribution. Vous êtes une source de soutien moral et j’ai utilisé sans limites ce soutien. Vous êtes une source de connaissances, variées et pertinentes, et je vous ai posé des questions sans arrêt. Dans le désordre, anciens et nouveaux : Amélie, Michaël, Sabrina, Julien H., Édouard, Dalie, Clara, Nicolas, Laurent, Mathieu, Glenn, Karine, Andrea, Alice-Anne, Alexandre, Valérie, Antoine, Maxime, Karina, Jérémie, Patrick, Pascale, Julien B., Mael, Barbara, Bob, Béatrice, Marianne, Myriam, Fred, Flo. Dalie, Clara, Maxime, vous m’avez particulièrement apporté du plaisir et de bons conseils. Merci pour votre dynamisme et votre intelligence. Julien Beguin, tu m’as amené à repousser mes limites. Sabrina, ta détermination et ton intelligence m’impressionnent. Michaël, tu es une force tranquille et inébranlable. Mael, sans toi le trajet n’aurait pas été le même. Merci pour ton soutien, les confidences et les conseils.
J’ai eu la chance de pouvoir compter au long de mon doctorat sur des amitiés durables, certaines datant de l’école primaire. Ces gens, rencontrés au fil de mes études et des amitiés communes, sont aujourd’hui connus sous le nom du club de tricot et du continent de carton. Merci à Anne, pour sa confiance et son côté pragmatique. Merci à François, Élizabeth, Perras, Émilie, David, Anne, Louis-Gab, Andréanne, Didier, Jeff, Amparo, Sophie et tous
xxi
les autres pour les soirées partagées. Votre perspective différente, intelligente, informée du monde me stimule. Merci également à mon club de lecture pour leur encouragement et les belles rencontres. Merci aux amies avec qui il fait toujours bon discuter, Aimée, Marianne et aussi Morgane et Victoria, rencontrées en cours de route.
J’ai une famille aimante, qui m’a soutenue dans tous mes projets, depuis de longues années. Comme le dit Bilbon, « je ne connais pas la moitié d’entre vous à moitié autant que je le voudrais ; et j’aime moins que la moitié d’entre vous à moitié aussi bien que vous le méritez ». Parmi ces gens, et en espérant ne pas faire de jaloux, je veux souligner l’apport de certains. Ma marraine, Johanne, a toujours été une présence réconfortante et encourageante. Jocelyne, merci pour ton enthousiasme devant mes projets et pour ta contribution à mes études. Merci à ma belle-famille, Sylvain, Johanne, Catherine et Bernard pour les beaux soupers et les chasses mémorables. Olivier et Jacinthe, je n’ai pas toujours été la grande sœur que j’aurais voulu être ou que vous méritez. Merci des beaux moments, magnifiés par la présence d’une belle-sœur souriante, Catherine et d’un beau-frère amusant, Benoît. Maman, nous partageons trop de qualités et de défauts pour que je puisse t’apprécier toujours à ta juste valeur. Merci pour ton soutien et l’exemple que tu donnes. Papa, ta passion pour ton travail est un modèle pour moi, ainsi que ton assurance tranquille. Je conclus ces remerciements avec ma principale source de soutien, de nourriture et d’amour sur une base journalière, Marc Antoine. Mon amoureux est une personne intelligente, drôle, affectueuse, avec qui l’on est bien, tout simplement. C’est une chance que tu sois dans ma vie et que tu sois resté tout au long de ce projet. Je sais que ça n’a pas toujours été facile et je t’en suis infiniment reconnaissante.
xxii
A
VANT
-
PROPOS
Cette thèse s’intéresse à l’impact des communautés végétales sur la consommation des plantes par les grands herbivores, avec comme sujet d’étude principal le cerf de Virginie (Odocoileus virginianus). La thèse comporte sept chapitres, soit une introduction et une conclusion en français ainsi que cinq chapitres sous forme d’articles scientifiques rédigés en anglais. Je suis l’auteure principale de tous les chapitres et la maître d’œuvre de ce projet, soit autant dans la conception, la prise de données, les analyses statistiques et l’interprétation des résultats. Les paragraphes suivants résumeront l’état actuel des chapitres rédigés sous forme d’articles scientifiques ainsi que la contribution respective des coauteurs.
Le chapitre 2 est une méta-analyse, publiée en juillet 2016 dans la revue Ecosphere. La version présentée ici est identique à celle publiée, à l’exception de quelques phrases dont la clarté a été améliorée. Ce chapitre a été réalisé en collaboration avec mon directeur de thèse, Jean-Pierre Tremblay, et mon codirecteur, Steeve D. Côté, professeurs au département de Biologie de l’Université Laval. Ces derniers ont participé à la conception du projet, à l’interprétation des résultats ainsi qu’à la révision et à l’approbation du manuscrit. Ils ont contribué de la même façon aux chapitres 3 à 6 et sont coauteurs de ces chapitres.
Les chapitres 3 et 6 ont été réalisés en collaboration avec André Dumont, biologiste responsable de la grande faune à la Direction de la gestion de la faune de l’Outaouais, Ministère des Forêts, de la Faune et des Parcs. Ce dernier a participé à la conception de l’expérience du chapitre 3, a contribué aux décisions reliées aux travaux de terrains des deux chapitres et révisé les deux manuscrits. Lucie Perroud, stagiaire en Master II à l’Université de Bourgogne est également coauteure du chapitre 3. Cette dernière a récolté des données vidéos, participé à l’analyse et à l’interprétation des données ainsi qu’à la révision du manuscrit. Le chapitre 3 a été soumis au Canadian Journal of Zoology en mars 2017. Le chapitre 6 sera soumis à un journal scientifique après le dépôt final de la thèse. Les chapitres 4 et 5 ont été partiellement rédigés lors d’un séjour à la Western Sydney University (Sydney, Australie), en collaboration avec Ben D. Moore, senior lecturer de
xxiii
cette institution et chercheur au Hawkesbury Institute for the Environment. Ce dernier m’a conseillé dans l’analyse des données, a participé à l’interprétation des résultats ainsi qu’à la révision du manuscrit. Ces chapitres seront soumis à un journal scientifique après le dépôt final de la thèse.
1
CHAPITRE 1
Introduction
2
Les interactions trophiques entre les producteurs primaires, les plantes, et les herbivores peuvent modifier la structure, la composition et le fonctionnement des écosystèmes (Hester et al 2006). Ces interactions sont complexes, parce qu’elles impliquent souvent des communautés végétales, où les herbivores ont une large abondance de ressources de qualité variable à leur disposition (Hairston et al 1960). Cette complexité végétale ouvre la porte à des interactions trophiques indirectes, plus dures à identifier, où l’interaction directe entre une plante et un herbivore est influencée par la présence d’une plante voisine (Abrams 1987; Morin 2009).
Dans une interaction directe entre un herbivore et une plante, la décision de consommer ou non la plante est principalement fonction de la profitabilité de cette plante pour l’herbivore. La profitabilité est la valeur en énergie ou en nutriments d’une plante, divisée par le temps requis par l’herbivore pour trouver et consommer cette plante (Pyke et al 1977). Cette profitabilité, variable entre les espèces, génère une hiérarchisation des espèces au long d’un gradient de préférence. La préférence est l’utilisation d’un item lorsqu’il est offert en quantité égale avec d’autres items (Johnson 1980). Le gradient de préférence est influencé par la présence de composés attractifs ou répulsifs dans les plantes (Molyneux et Ralphs 1992). Par exemple, les herbivores préfèrent et sélectionnent généralement les espèces riches en azote, élément essentiel dans les processus métaboliques et dans la structure cellulaire (Mattson 1980). À l’inverse, certains herbivores comme les ruminants évitent les espèces riches en fibres qui réduisent la digestibilité (Danell et al 1994b; Dumont et al 2005; Forsyth et al 2005) et les composés de défense comme les tannins, qui peuvent réduire la digestibilité et la disponibilité des protéines (Robbins et al 1987a; Robbins et al 1987b).
La consommation d’une plante n’est toutefois pas uniquement basée sur les préférences de l’herbivore et dépend, entre autres choses, de l’effet indirect des plantes environnantes. L’influence des ressources alternatives sur le comportement d’approvisionnement des herbivores est notamment décrite par la théorie de l’approvisionnement optimal. Élaborée dans les années 60 et 70, cette théorie décrit mathématiquement ce comportement comme une série de choix qui visent à maximiser la quantité d’énergie consommée ou à minimiser le temps d’alimentation (MacArthur et Pianka 1966; Charnov 1976; Pyke et al 1977). Dans
3
certains systèmes et pour certains herbivores, c’est plutôt l’acquisition de nutriments spécifiques qui est optimisée (Raubenheimer et al 2009) ou la capacité digestive de l’herbivore (Verlinden et Wiley 1989). En vertu de la théorie classique de l’approvisionnement optimal, il est possible de déterminer le régime alimentaire d’un animal en fonction de la profitabilité des plantes disponibles. La présence d’un item dans ce régime alimentaire optimal ne dépend pas de sa disponibilité, mais plutôt de la disponibilité d’items relativement plus profitables (Pyke et al 1977).
Expérimentalement, on a remarqué que le comportement d’approvisionnement des animaux diffère fréquemment des prédictions mathématiques de l’approvisionnement optimal (Schluter 1981). Les premières études sur l’approvisionnement optimal étaient effectuées dans des milieux simples contrôlés alors que les environnements naturels sont complexes et proposent une large gamme d’items à consommer (Schluter 1981). Par exemple, le comportement d’échantillonnage de la mésange charbonnière (Parus major) crée une alimentation sous-optimale, car les mésanges consomment des proies de faible profitabilité, même lorsque l’abondance des proies plus profitables est élevée (Krebs et al 1977). Ce comportement leur permet toutefois de déterminer la profitabilité et l’abondance des proies disponibles. De plus, des considérations non alimentaires peuvent influencer l’approvisionnement, comme le besoin pour l’herbivore de se protéger des conditions climatiques (Mysterud et Østbye 1999), de diminuer les coûts de locomotion (White et Yousef 1978) ou de se protéger d’un prédateur (Brown et al 1999). Toutefois, la théorie de l’approvisionnement optimal et ses dérivées (voir Raubenheimer et al 2009) procure un cadre théorique pertinent à l’étude de l’approvisionnement des herbivores, tenant compte à la fois des effets directs de la qualité des plantes et des effets indirects des plantes environnantes.
Parallèlement aux études sur l’approvisionnement des herbivores, des chercheurs ont abordé les interactions trophiques indirectes selon une perspective végétale. Les hypothèses d’effets associatifs (Milchunas et Noy-Meir 2002; Barbosa et al 2009), décrivent comment la présence d’une plante environnante module la consommation par les herbivores d’une plante d’intérêt. Quatre types d’effets associatifs ont été décrits, définis notamment par la préférence de l’herbivore entre la plante d’intérêt et la plante environnante (Figure 1.1). La
4
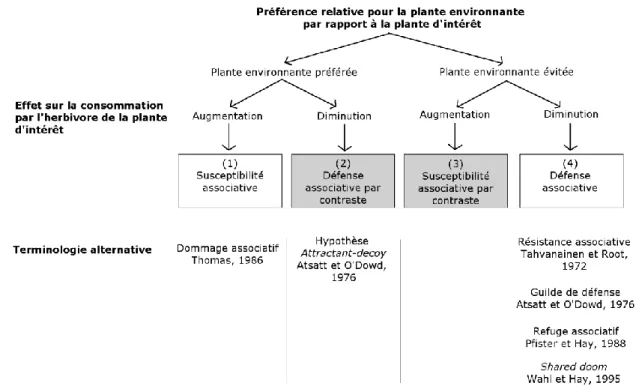
présence d’une plante environnante relativement préférée peut augmenter ou diminuer la sélection pour la plante d’intérêt ce qui correspond respectivement à 1) la susceptibilité associative et 2) la défense associative par contraste (Figure 1.1; Thomas 1986; Hjältén et al 1993; Bergvall et al 2006). De même, la présence d’une plante environnante relativement évitée peut augmenter ou diminuer la sélection pour la plante d’intérêt, ce qui correspond respectivement à 3) la susceptibilité associative par contraste et 4) la défense associative (Figure 1.1; Tahvanainen et Root 1972; Atsatt et O'Dowd 1976; Bergvall et al 2006).
Figure 1.1 Organigramme des quatre types d’effets associatifs. Les effets associatifs se différencient par la préférence de l’herbivore pour la plante environnante, relativement à la plante d’intérêt. Les plantes environnantes, qu’elles soient relativement préférées ou évitées, peuvent augmenter ou diminuer la consommation de la plante d’intérêt. Dans cette thèse, j’utilise la terminologie de Bergvall et al (2006), seule à introduire les effets par contrastes.
Ces deux groupes d’hypothèses, l’approvisionnement optimal basé sur une perspective animale et les effets associatifs basés sur une perspective végétale peuvent converger dans une vision commune des effets indirects des plantes sur les interactions plante-herbivore. Notamment, des chercheurs ont proposé que le mécanisme des effets associatifs puisse être l’approvisionnement optimal. Par exemple, la maximisation de l’acquisition d’énergie chez le bison (Bison bison) crée des défenses associatives par contraste et des susceptibilités
5
associatives (Courant et Fortin 2010). Dans le premier cas, des plantes ne sont pas consommées en présence de Carex atherodes, car ces plantes sont plus petites. Les consommer, en même temps que C. atherodes, augmente le temps de manipulation pour l’herbivore et diminue donc la profitabilité de la période d’alimentation. À l’inverse, les plantes de même taille que C. atherodes subissent des susceptibilités associatives, car les consommer ne change pas le temps de manipulation. Ce test explicite des principes d’optimalités pour expliquer les effets associatifs reste rare dans les études avec une perspective végétale. Plus fréquemment, les comportements d’approvisionnement sont invoqués qualitativement pour expliquer les effets associatifs (pour exemples, voir Miller et al 2006; 2007; Miller et al 2009; Emerson et al 2012). Bergvall et al (2006) propose un mécanisme se basant sur les niveaux de sélectivité variable des herbivores pour expliquer la différence entre effets associatifs « sans contraste » (Figure 1.1, boîtes blanches) et avec contraste (boîtes grises). Lorsqu’un herbivore est sélectif entre les parcelles, il sélectionne les parcelles contenant des espèces préférées, générant des susceptibilités associatives à l’échelle des parcelles. Inversement, l’évitement d’une parcelle contenant les espèces les moins préférées suscitera des défenses associatives. Lorsque l’herbivore effectue une sélection dans une parcelle, il choisit l’espèce la plus profitable de celles qui sont présentes. Ce comportement génèrera alors des effets associatifs par contraste pour les plantes dans les parcelles. Dans l’étude de Bergvall et al (2006), les daims européens (Dama dama) étaient peu sélectifs entre des parcelles (7 m2), mais sélectifs à l’intérieur des parcelles, ce qui a effectivement produit des défenses associatives par contraste et des susceptibilités associatives par contraste. Par exemple, la consommation de moulée faible en tannins (des composés peu digestibles) était plus grande dans les parcelles où il y avait de la moulée riche en tannins (Bergvall et al 2006). Dans une expérience d’alimentation en milieu naturel, des écureuils (Sciurus niger et S. carolinensis) ont démontré une forte sélectivité entre des parcelles qui a mené à des susceptibilités associatives et des défenses associatives (Emerson et al 2012).
Même lorsqu’elles ne sont pas étudiées de concert, les hypothèses sur l’approvisionnement et les effets associatifs concordent en ce qui concerne le rôle central de la qualité nutritive des plantes (c.-à-d. profitabilité et composés attirants ou répulsifs) dans la sélection des ressources alimentaires. Selon la théorie d’approvisionnement optimal, c’est la profitabilité
6
relative des plantes qui détermine leur utilisation, soit leur contenu en énergie ou en nutriments (Pyke et al 1977; Raubenheimer et al 2009). Dans le contexte des effets associatifs, on invoque plutôt la préférence relative (Miranda et al 2011), qui peut être liée à la profitabilité, mais tiendrait compte de la présence de composés attirants ou répulsifs, n’influençant pas la profitabilité (Molyneux et Ralphs 1992). Autant dans les études d’approvisionnement que dans celles d’effets associatifs, les différences de qualité nutritive considérées sont généralement interspécifiques. Pourtant certains herbivores, comme les cervidés (Hanley 1982), ajustent également leur choix alimentaire en fonction de la variation intraspécifique en profitabilité et en composition chimique. Par exemple, le daim européen (Dama dama) préfère la nourriture artificielle faible en tannins à une nourriture identique, mais riche en tannins, avec un impact sur les effets associatifs (Bergvall et al 2006). En milieu naturel, Moore et al (2010) ont démontré que les koalas (Phascolarctos
cinereus) sélectionnent plus fortement les Eucalyptus entourés de grands arbres de forte
qualité nutritive (susceptibilité associative) ou de petits arbres de faible qualité (susceptibilité associative par contraste). Le tétras des armoises (Centrocercus
urophasianus) pourrait créer des effets associatifs, car il effectue une sélection entre des
parcelles basée sur leur composition chimique (Frye et al 2013). Ces résultats récents suggèrent que les études des deux domaines, approvisionnement des herbivores et effets associatifs, bénéficieraient d’incorporer les variations intraspécifiques en qualité nutritive. À cause de leur évolution parallèle, certains thèmes communs dans les études avec une perspective animale ont eu peu d’échos dans les tests d’effets associatifs. Par exemple, une avenue de recherche répandue associée à la théorie de l’approvisionnement optimal concerne l’effet du risque de prédation par les carnivores. Cet effet est une autre interaction trophique indirecte, où l’impact des herbivores sur les plantes est modulé par un carnivore. En présence d’un carnivore ou lorsqu’ils perçoivent un risque de prédation, les herbivores peuvent modifier leur comportement pour réduire le risque d’être consommé, potentiellement aux dépens de la maximisation de l’acquisition d’énergie (théorie de l'écologie de la peur; Brown et al 1999). Par exemple, le temps passé à s’alimenter diminue en milieu risqué, car un herbivore consacrera plus de temps à des activités antiprédatrices (Brown et al 1999; Lima et Bednekoff 1999). Les changements de comportement peuvent également affecter le choix du lieu d’alimentation : les herbivores de la forêt de Białowieża
7
utilisent moins les plantes situées dans le centre du territoire des meutes de loups (Canis
lupus) que dans les zones moins utilisées de ces territoires (Kuijper et al 2013). Cette
différence d’utilisation se traduisait en une diminution de 8% de l’intensité du broutement sur les arbres au centre dy territoire des loups. Le risque de prédation pourrait pareillement modifier les espèces consommées par un herbivore. Un herbivore s’alimentant dans un milieu risqué se concentrerait sur les items les plus profitables, serait donc plus sélectif, pour tirer parti du court temps passé à s’alimenter (Brown et Kotler 2004). Si le risque de prédation rend l’herbivore plus sélectif, il pourrait y avoir une diminution des susceptibilités associatives, car certaines susceptibilités associatives sont le résultat d’une consommation peu sélective (Wang et al 2010; Milligan et Koricheva 2013). Dans un des rares tests de l’effet du risque de prédation sur la sélectivité, Morrison et al (2004) ont démontré que les pikas (Ochotona collaris) réduisaient leur alimentation dans les environnements plus risqués, sans changer leur sélectivité. Aucune étude ne semble avoir exploré la relation entre le risque de prédation des grands herbivores et les effets associatifs.
Une autre thématique répandue dans les études avec une perspective animale, mais qui a été peu explorée dans les études d’effets associatifs est l’importance de l’échelle spatiale, soit la distance à laquelle les plantes environnantes influenceront l’utilisation des herbivores. La notion d’échelle spatiale est utilisée de façon variable par les écologistes (Fortin et Dale 2005). Dans cette thèse, je définis l’échelle spatiale comme une combinaison de l’espace physique où se produit un processus (étendue) et de la taille des unités d’observations (grain) (Wiens 1989). Cette échelle est parfois définie en mesures fixes (mètres) ou relatives (parcelle, domaine vital, paysage). La sélection des ressources serait en fait un processus hiérarchique, dépendant de décisions prises à plusieurs échelles spatiales (Johnson 1980; Senft et al 1987; Bailey et al 1996). Johnson (1980) a proposé les niveaux de sélection suivants : distribution géographique de l’espèce, domaine vital de l’individu, habitat dans le domaine vital, site d’alimentation. Ces niveaux grossiers peuvent être divisés plus finement, mais ils illustrent efficacement ce concept de sélection hiérarchique. Le choix d’un site d’alimentation sera contraint par le choix du domaine vital, par exemple. En réalité, la sélection des ressources est probablement continue, limitée uniquement par la perception et la mémoire des herbivores et par des barrières physiques bien définies. Aux
8
plus larges échelles, Rettie et Messier (2000) ont proposé que les facteurs avec le plus grand impact sur la valeur adaptative, par exemple le risque de prédation, déterminent la sélection d’un domaine vital. La sélection des ressources alimentaires se déroulerait à des échelles plus fines, comme au niveau du site d’alimentation et de la plante (Rettie et Messier 2000).
Les études des effets associatifs sont souvent effectuées à un grain très fin avec des parcelles de quelques mètres carrés, un problème commun dans les études végétales (Vellend 2016). Ceux qui ont intégré une composante spatiale à l’étude des effets associatifs ont souvent comparé l’utilisation à l’intérieur d’une parcelle de végétation vs entre des parcelles (Hjältén et al 1993; Bergvall et al 2006; Huang et al 2012; Stutz et al 2015b). C’est notamment le cas du cadre proposé par Bergvall et al (2006) décrit plus haut. Par contre, Hester et Baillie (1998) ont étudié la susceptibilité associative avec une approche spatiale continue qui leur a permis d’en déterminer l’étendue. Dans cette étude, les effets des parcelles de graminées sur la consommation de Calluna vulgaris par le cerf élaphe et le mouton (Ovis aries) disparaissaient lorsque la distance entre les graminées et C.
vulgaris atteignait 1 à 3 m. Si les effets associatifs sont générés par un mécanisme
d’approvisionnement optimal, on peut supposer qu’ils seront hiérarchiques et soumis aux mêmes compromis que la sélection des ressources. Underwood et al (2014) ont proposé une relation entre effets associatifs et échelle spatiale, allant des relations entre plantes à l’intérieur d’une parcelle jusqu’aux différences d’utilisation entre des parcelles, soit des distances appropriées pour des insectes. Selon ce cadre, à fine échelle, la plante environnante influence l’interaction directe entre la plante d’intérêt et l’herbivore, sans qu’il y ait d’interaction directe entre la plante environnante et l’herbivore. Par exemple, la compétition avec la plante environnante diminue les ressources disponibles pour la plante d’intérêt qui ne peut plus produire de composés de défenses. À l’échelle d’une parcelle, le taux de rencontre de l’herbivore avec les différentes espèces de plantes influence sa consommation de la plante d’intérêt. Il n’y a actuellement aucune hypothèse généralisant les effets associatifs à travers l’échelle spatiale de sélection des ressources par les grands herbivores.
9
Autant pour les études à partir de l’angle de l’approvisionnement des herbivores que de celui des effets associatifs entre les plantes, les effets étudiés présentent généralement un nombre limité de ressources alternatives ou plantes environnantes. L’interaction trophique indirecte à trois composantes, deux plantes et un herbivore constitue d’ailleurs la majorité des études sur les effets associatifs (Barbosa et al 2009). Dans un milieu où une seule espèce de plante permet un approvisionnement optimal, ce raccourci peut être suffisant. Par exemple, le bouleau verruqueux (Betula pendula) génère des effets associatifs plus importants que d’autres espèces ligneuses, car il est fortement sélectionné par l’orignal (Alces alces) et fortement évité par les campagnols (Microtus spp. et Clethrionomus spp.; Vehviläinen et Koricheva 2006). Dans une communauté végétale naturelle, il y a probablement plusieurs effets indirects à l’œuvre, simultanément. Cette complexité est habituellement considérée en utilisant l’une des propriétés des communautés végétales, soit la richesse spécifique ou la diversité. La relation entre diversité et effets associatifs a été très étudiée chez les insectes. Généralement, le niveau de dommage sur la communauté par les insectes est plus faible dans les parcelles plus riches et diversifiées (défense associative; Brown et Ewel 1987; Castagneyrol et al 2014). Cet effet dépend toutefois du niveau de spécialisation de l’herbivore et la relation entre dommage et diversité est plus forte lorsque ces insectes sont des spécialistes (Castagneyrol et al 2014). Pour un insecte monophage, une forte diversité diminue la densité relative de la plante préférée et donc la quantité relative de ressources disponible. Comme cette défense associative est liée à la spécialisation de l’herbivore, il est douteux qu’elle s’applique aux grands herbivores (Vehviläinen et Koricheva 2006), car ils sont majoritairement des généralistes (voir les données récoltées par Barbosa et al 2009). Avec l’orignal, on observe plutôt des susceptibilités associatives en lien avec la richesse. En Finlande, l’orignal consommait plus de biomasse dans les parcelles plus riches en espèces (Vehviläinen et Koricheva 2006; Milligan et Koricheva 2013). Cette susceptibilité associative peut s’expliquer par la présence de plus d’espèces préférées par l’orignal dans ces parcelles riches (Vehviläinen et Koricheva 2006) ou par une diminution de la sélectivité (Wang et al 2010; Milligan et Koricheva 2013). En effet, les herbivores seraient moins sélectifs dans les milieux multispécifiques où les espèces sont réparties de façon hétérogène parce que la sélectivité y serait plus complexe et donc plus coûteuse (Wang et al 2010). En se basant sur la théorie de