A superfast muscle in the complex sonic apparatus of Ophidion rochei (Ophidiiformes): histological and physiological approaches
Texte intégral
Figure


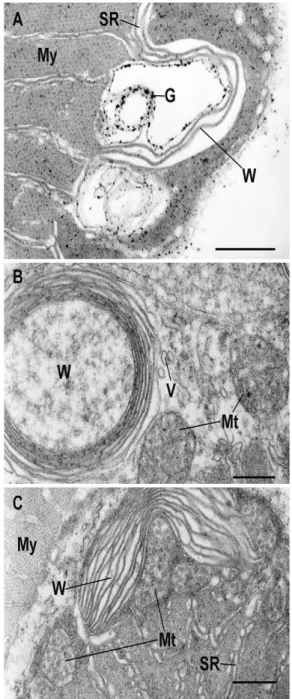
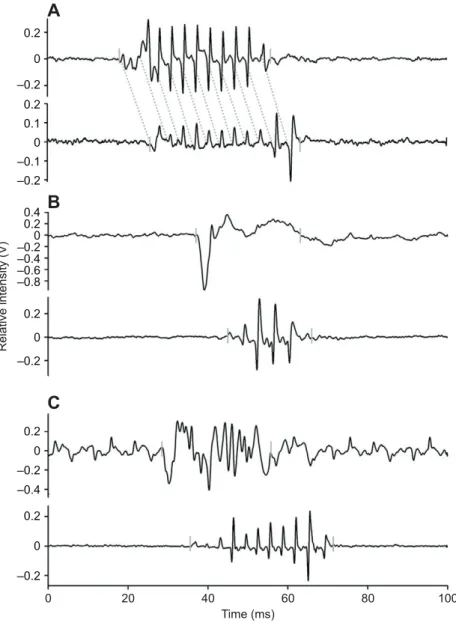

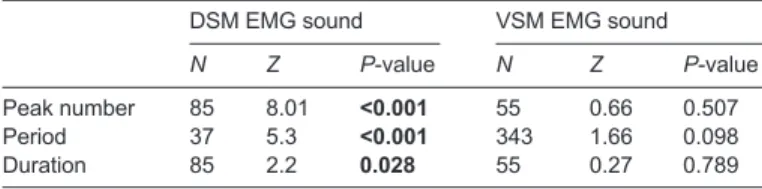
Documents relatifs
Dans ce sens, la formation d’alliances, de coalitions, de consortiums, de groupements de firmes, et plus récemment de réseaux d’alliances, contribue de manière
Keywords Horizontal collaborative networks · Business share · Trust · Product realisation graph · Partner selection Introduction.. The enterprises today are aware of the
The track is updated as space track adding the 3 hits; the Z position of the R hits is adjusted as we know now x and y; the track is fitted, and the highest χ 2 is computed and has
Therefore, the aim of the present study was (i) to characterise BMI and cardiovascular function (heart rate and blood pressure) in children and adolescents of this
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des
a) Dans une heure, Jean-Pierre ira consulter les archives.. « Nous ne pouvons tout de même pas être réduits à appeler la force armée contre nos ouvriers.. a) Le professeur demande
While colour pattern has been shown to be the most reliable character for species identification in several Entomobryidae genera, it might not be the case in Iso- tomurus
Keywords: phraseological pattern; productivity; semantic classification; open slot; corpus linguistics; dictionary of
