Association of milk traits with SSCP polymorphisms at the
growth hormone gene in the serrana goat
P. X. Marques
P. X. Marques
aa
; M. Pereira
; M. Pereira
bb; M. R. Marques
; M. R. Marques
a, ba, b; I. C. Santos
; I. C. Santos
a,ba,b; C. Belo
; C. Belo
bb; R.
; R.
Renaville
Renaville
ccand A. Cravador
and A. Cravador
a*a*aFaculdade de Engenharia de Recursos Naturais, Universidade do Algarve, Campus de Gambelas,
8000-117 Faro, Portugal. E-mail: mrmarques@ualg.pt; icsantos@ualg.pt; acravad@ualg.pt
bEstação Zootécnica Nacional, Departamento de sistemas e técnicas de produção animal, Fonte Boa,
2000-763 Vale de Santarém, Portugal. E-mail: dovi.ezn@mail.telepac.pt
cAnimal and microbial Unit, Gembloux Agricultural University, 13 Avenue Maréchal Juin, 5030
Gembloux, Belgium
*Corresponding author. Tel.: +351 289 800935; fax: +351 289 818914; E-mail address: acravad@ualg.pt
Abstract
The present study supports the existence of an association between milk
production traits and genetic polymorphisms at the GH gene in the Portuguese
indigenous Serrana goat. The DNA from 229 animals of two ecotypes (Jarmelista and
Ribatejano) was analysed by single-strand conformation polymorphism (SSCP)
technique revealing a high degree of genetic polymorphisms at the goat GH (gGH)
gene. Two conformational patterns were detected in exons 1 and 2, 6 in exon 3, 10 in
exon 4 and 5 in exon 5. The evaluation of an association effect between these SSCP
patterns with milk, fat and protein yields and fat and protein percentage revealed a
positive effect for pattern A/B of exon 4 in Ribatejano ecotype and for pattern A/B of
exon 2 with milk yield (P < 0.05) in Jarmelista ecotype and for pattern A/B of exon 1
and pattern B/B of exon 2 with protein percentage (P < 0.05) in Ribatejano ecotype.
The results provide support for the hints suggested by previous studies at the importance
of the gGH gene as a candidate gene for marker-assisted selection in goat breeds and
suggest that exon 4 is a preferential target for further investigation on mutations that
influence milk yield variation.
Keywords
:
Polymorphism; Single-strand conformation polymorphism; Genetic markers; Growth hormone, Goat, Milk traits.1.
INTRODUCTION
Since its finding in the 1920s, it is well documented that growth hormone (GH)
influences animal process as growth, lactation, reproduction and metabolism (Bauman,
1999, Baldi, 1999, , Breier et al., 1999, Scaramuzzi et al., 1999)).
Less controversial than the exogenous administration of GH to be exploited as a
production stimulator, is the strategy based on the selection of animals with an
endogenous GH secretion resulting in a superior quantitative trait. This productive
advantage can be the result of a more active GH variant or of a more favourable
upstream regulation directing its synthesis. The establishment of significant correlations
between quantitative traits and molecular variation at the level of GH or of factors
governing its secretion and action could lead to the identification of markers useful for
assisting selection programmes.
Genetic polymorphisms of GH have been reported in various domestic livestock,
mainly e in cattle and several studies have related association effects between bovine
GH (bGH) polymorphisms and milk yield traits. Due to an insertion of a T at position
+837 and a C-G transition at position +838, a MspI RFLP is present in the third intron
of the bovine GH gene (Zhang et al., 1993). Høj et al. (1993) observed a difference in
allele frequencies at the GH loci between two Danish and Norwegian lines, the MspI (-)
haplotype being more frequent in the line selected for milk fat. Lee et al. (1993, 1994)
found also a significant association of MspI (-) with higher fat yield in Holstein cows. In
a recent study, Lagziel et al. (1999) found a significant increasing effect of MspI (-)
allele on protein percentage and kg of protein per year.
With the PCR-RFLP technique using the AluI restriction enzyme, Lucy et al. (1993)
revealed two alleles responsible for alternative forms of bovine GH with a Leu or Val
residue at position 127. Injection of recombinant bovine GH with a valine residue
corresponding at the Leu/Val polymorphic locus to lactating cows has been shown to
improve fat corrected milk production more than GH-Leu
127administration (Eppard et
al., 1992). This observation suggests that the Val allele genotype is better for milk
production. On the other hand, Lucy et al. (1993) reported a significant higher predicted
transmitting ability (PTA) for milk yield for Val/Val genotype Jersey cows, but no
significant effect was found in their samples of Holstein, Guernsey, Ayrshire or Jersey
bulls. However, Furu et al. (1998), Schlee et al. (1994a) and van der Werf et al. (1998)
observed no significant effect of this Leu/Val polymorphism on milk trait breeding
values of Simmental, Holstein or Jersey sires. On the other hand, Lee et al. (1996) and
Lucy et al. (1993) reported a decreased milk yield associated with the valine variant of
the GH gene in Holstein cows. Further studies by Schlee et al. (1994b) in German Black
and White bulls revealed that animals homozygous for the leucine variant had higher
plasma levels of GH than their heterozygous counterparts.
More recently, Grochowska et al. (2001) observed an association between this
polymorphism and stimulated GH release by TRH, IGF-1 and also milk and protein
yields in Polish Friesen cattle
Other polymorphic sites were described by Zhang et al. (1993) and by Unanian et
al. (1994) who reported PCR-RFLP polymorphisms in exon 3 at position 1547 by MspI
digestion and in the 3' region at position 2637 of bGH by HaeIII digestion respectively.
An investigation of the association between bGH loci corresponding to these GH-MspI,
GH-HaeIII polymorphisms and to the GH-AluI polymorphism and milk yield traits in
Holstein cattle by means of pedigree analysis concluded that the bGH complex is linked
to one trait locus affecting protein percentage and that it may be linked to a quantitative
trait loci (QTL) affecting other milk traits (Vukasinovic et al, 1999).
Genetic polymorphisms have also been detected by SSCP in the GH gene
(Kirkpatrick, 1992, Lagziel et al., 1996, Yao et al, 1996) and in other genes (Neibergs
et al., 1993) in cattle and in the goat GH gene (Malveiro et al., 2001)..
Most significant
studies using the SSCP approach were accomplished on bovines in linkage analysis
(Lagziel et al., 1996) and to define intragenic haplotypes for GH in Israeli Holstein
cattle. Lagziel
et al. (1996) analysed nine fragments of the bovine GH gene, identified
eight SSCP haplotypes, and associated one of them with a high milk protein percentage.
The SSCP haplotypes were later sequenced disclosing 12 sequence variants (Lagziel et
al., 1999). Using the SSCP approach Yao et al. (1996) have reported two
polymorphisms (TC in intron 3 and A C in exon 5), which were associated with
milk production traits, in the bGH gene of Holstein cattle. In goats, the only study on
polymorphism at goat GH gene (gGH) was described by Malveiro et al. (2001) who
reported several SSCP patterns in the Algarvia goat. Two of these patterns were
positively associated with milk traits.
The Serrana goat is a Portuguese autochthonous breed of unknown origin with
high productive potentialities, high rusticity, and good productive and reproductive
indexes in extensive systems and good expansion possibilities. This breed, the most
numerous in Portugal is reared mainly in a region covering the centre and the north of
Portugal. In particular, Ribatejano ecotype is reared in areas around the river Tejo
(Ribatejo region) and Jarmelista ecotype in a climatically and topographically distinct
region covering an interior area southern of river Douro (Beira Alta region). Milk, used
in cheese manufacture, and meat are two important economic resources provided by the
Serrana goat to rural populations. Well adapted to its natural environment showing good
milk performances in poor feeding conditions its preservation is nevertheless threatened
by the introduction of foreign breeds characterized by higher milk production but
unadapted to traditional Portuguese grazing conditions. Genetic selection within this
breed aimed at the improvement of production traits may be the best strategy to assure
its preservation and contribute to biodiversity conservation. As a first step towards these
objectives we have analysed the five exons of the gGH gene in the Serrana breed by
polymerase chain reaction (PCR)-SSCP and present in this paper the results obtained as
well as the results of the analysis aimed at the establishment of an association of the
identified single-strand polymorphisms with milk, fat and protein production and fat
and protein content.
2.
Materials and methods
2.1
Animals
Two hundred and twenty-nine Serrana goats (2-11 years old) were used in the present study were selected from farmers of Ribatejo (111 females and 11 males, Ribatejano ecotype)) and Beira Alta (100 females and 7 males, Jarmelista ecotype) regions of Portugal. The animals were randomly chosen from herds provided by the breeders associations "Associação de Criadores de Reprodutores de Gado Oeste" (ACRO), Associação de Criadores de Caprinos e Ovinos do Ribatejo Oeste" (ACORO) and "Associação de Criadores da Guarda" (AGRIGUARDA).The mean productive values were as follows: milk 250 L; fat: 10 kg; protein: 9 kg; fat yield: 3,5%; protein yield: 3,3%.
DNA was extracted from peripheral blood leukocytes using a DNA Isolation Kit from Puregene.
2.2
PCR amplification
The 5 exons of the gGH gene were amplified by PCR using the five prime pairs shown in Table 1. The resulting five amplification fragments ranged from 112 to 287 bp. PCR reactions were performed in a UNOII thermocycler (Biometra) using Ready-to-Go PCR Beads (Amersham Biosciences) according to the following conditions: 25-50 ng of genomic DNA; 4 to 16 pmol of each primer (Table 1); 1,5 U Taq DNA polymerase, 10 mM Tris-HCl, pH 9; 50 mM KCl; 1,5 mM MgCl2; stabilisers, including BSA, for a final volume of 25 l. The
amplification included a initial denaturation at 95ºC for 5 min, 30 cycles of denaturation at 95ºC for 30 s; annealing at 57-62C (Table 1) for 30 s and extension at 72C for 30 s followed by a final extension at 72ºC for 5 min. The product of each amplification was analysed by electrophoresis on 2% agarose gel (5 V/cm), using ethidium bromide staining.
Table 1.
SSCP fragments at the gGH gene and PCR analysis parameters
Exon Fragment length andlocalisationa (bp)* Primer sequence
b Annealing
temperature (ºC) Primers(pmol) DNA(ng)
1 112 360-471 5'-CAGAGACCAATTCCAGGATC-3' 5'-TAATGGAGGGGATTTTTGTG-3' 57 16 25 2 202 673-874 5'-GACACATCTCTGGGGAGCTT-3'5'-CCTCCTGGTCTCTCCCTAGG-3' 62 16 50 3 157 1063-1219 5'-GTGTGTTCTCCCCCCAGGAG-3'5'-CTCGGTCCTAGGTGGCCACT-3' 60 4 25 4 200 1416-1615 5'-GAAGGGACCCAACAACGCCA-3'5'-CTGCCAGCAGGACTTGGAGC-3' 60 8 25 5 287 1856-2142 5'-AAAGGACAGTGGGCACTGGA-3'5'-CTTGGCAGGAGCTGGAAGAT-3' 62 16 50
a According to published gGH gene sequence (Kioka, et al., 1989).
b Primers (Life Technologies) were designed according to published gGH gene sequence (Kioka, et al., 1989);
*bp: base pairs.
2.3.
SSCP analysis
For SSCP analysis, 5 L (or 10 L) of each amplification product was added to 10 µl (or 15 l) of stop solution (95% formamide, 10 mM NaOH, 0,05% xylene cyanol and 0,05% bromophenol blue). The samples were heat-denatured at 95C for 5 min, chilled at 0C, and loaded (20 L or 25 L of each) onto an 8-12% polyacrylamide/TBE gel, with a 2,5% crosslinking. Gels were run at 25 W for 4-8 h, at 15-20ºC, in a DCode Universal Mutation Detection System, (BIO-RAD), coupled to a refrigeration system. The SSCP analysis parameters were optimised for each fragment (Table 2). After the run, the gel was removed from the apparatus and silver stained. The observed patterns and frequencies are shown in Table 3.
Table 2. Optimised conditions of SSCP analysis for the 5 exons of the gGH gene.
Exon DNA (l) Stop Solution (l)
%Ta Temp(C)b Bufferc Time
(h)d 1 10 10 12 15 1x 8 2 5 15 10 15 0,5x 4:30 3 5 15 12 15 1x 7:30 4 5 15 10 15 0,5x 6:15 5 10 10 8 20 0,5x 4
a Polyacrylamide percentage; b electrophoretic temperature; c buffer concentration; d electrophoretic running time
Table 3: SSCP analysis parameters and results for the indicated exons Ribatejano ecotype (n = 122)
Exon Number of patterns Frequencies of patterns (%)
1 2 A/B (82,8); A/A (17,2)
2 2 A/B (90,2); B/B (9,8)
3 5 A/B (74,6); C/D (2,5); A/B/D (15,6); A/B/C/D (5,7); A/B/D/E (1,6) 4 5 A/B (68,9); A/C (9,8); A/B/C (13,1); A/B/D/E (1,6); A/B/C/F (6,6) 5 5 A/B (70,5); A/C/D (12,3); A/B/C/D (13,1); A/BC/E (2,5); A/B/C/F (1,6) Jarmelista ecotype (n = 107)
Exon Number of patterns Frequencies of patterns (%)
1 2 A/B (84,1); A/A (15,9)
2 2 A/B (93,5); B/B (6,5)
3 6 A/B (72); C/D (0,9); A/B/D (6,5); A/B/C/D (9,4); A/B/D/E (5,6); A/C/D (5,6) 4 9 A/B (54,2); A/C (0,9); A/B/C (15,9); A/B/C/F (16,8); A/C/F (1,9); A/B/E (5,6); C/F
(2,8); A/A (0,9); A/B/F (0,9)
5 4 A/B (57); A/C/D (2,8); A/B/C/D (34,6); A/B/C/F (5,6)
Using this method, animal genotype that showing more than 4 bands for several patterns can be interpreted as the result of GH gene duplication in goats. Indeed RFLP studies have demonstrated that there are two alleles at the GH gene locus in sheep and goats (Valinsky et al., 1990). In sheep, the GH1 allele presents a single copy of the gene (GH1) while the GH2 allele presents two GH copies in tandem (GH2-N and GH2-Z). Individual animals homozygous for GH1 or GH2 alleles have two or four GH-like genes, respectively; while heterozygous animals with one copy of GH1 and one of GH2 (GH1/GH2), have three GH-like genes (Wallis et al., 1998). Moreover, sequence differences between the sheep GH2-N and GH2-Z genes have been demonstrated(Ofir and Gootwine, 1997). In goats, a similar allelic polymorphism was observed revealing an allele, CgGH, with a single GH copy, gGH1, and the EgGH allele with the duplicated gene presenting also a tandem arrangement of the gGH2 and gGH3 genes (Yamano
et al., 1991). The presence of this allelic polymorphism makes the identification of sequence
polymorphism more difficult. One can thus expect to observe from 2 to 6 bands in SSCP analysis. Animals homozygous for GH1 can theoretically not give rise to more than 4 bands for each of the 5 exons of gGH. In the sample of 229 Serrana goats studied individual animals exhibiting 2 and/or 4 bands in each of the 5 exons could be homozygous for the GH1 allele. However, the possibility that they are heterozygous, i. e. GH1/GH2, or homozygous for the same sequence variant of the GH2 allele (for instance gGH2 and gGH3 are each homozygous considered separately but are heterozygous between each other) can not be excluded. The letters used below to designate the different patterns refer to sequence variants of any of the GH-like genes as it is impossible with the present analysis to establish which of them is homozygous or heterozygous.
2.4.
Statistical analyses
From the 229 animals analysed by PCR-SSCP, 18 are males and one female had insufficient data, so only 210 animals were statistically analysed.
The data collected included cumulative milk yield, total milk production (yield), total milk yield adjusted for 150 days and mean of daily milk yield for both ecotypes- Jarmelista and Ribatejano and fat and protein yield, fat and protein percentage adjusted to 150 days only for Ribatejano ecotype. These variables were measured in seven consecutive years (1993-1999). Data have been considered as repeated measures and the correlations between the measures in the same individual were considered in the statistical model.
Linear mixed models, which included the fixed effects of ecotype, farmer (ecotype), type of parturition, linear and quadratic effect of the covariable age, exons 1-5 and considering repeated measures on the animal were used to determine associations between SSCP and milk traits (SAS System).
Four models were tested: a classical mixed model;
a fixed effects model with power process (time as discrete variable); a fixed effects model with power process and measurement of error; a fixed effects model with an unspecified covariance structure
The models were compared using Akaike's Information Criterion and Schwarz's Bayesian Criterion, in order to choose the model with the best fit. The nearest zero values were obtained with model d (unspecified covariance structure) for all milk traits when considering both ecotypes in the model and model b (model with fixed effects and power process) when ecotypes were analysed separately.
The F-values for fixed effects of exons 1 to 5 were not significant for p<0.05 for all milk traits. Since not all patterns were observed for some exons in the two ecotypes (Jarmelista's had no A/B/D/E pattern in exon 4 and A/B/C/E pattern in exon 5 and Ribatejano's lacks A/C/D pattern in exon 3 and A/C/F, A/B/E, C/F, A/A, A/B/F patterns in exon 4), we analysed milk traits for each ecotype separately.
To determine associations between milk traits for both ecotypes and fat and protein traits on ecotype Ribatejano we also used the four linear mixed models already described but only with the fixed effects of flock, type of parturition, linear and quadratic effect of the covariable age and exons 1-5 and considering repeated measures on the animal.
The effect of type of parturition wasn’t significant for a p-level of 0.05 for all traits so it was removed from the models
When F-test for the fixed effects were significant (p<0.05) we use multiple comparison tests (LSD) of LSMEANS.
3.
Results and discussion
SSCP polymorphisms
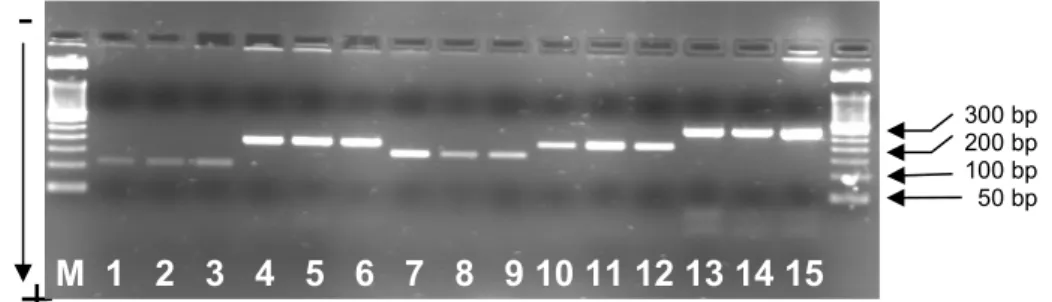
SSCP analysis of exons 1 to 5 of the gGH were performed on the fragments amplified by PCR using the primers described in Table 1. which showed expected lengths (Figure 1)
Figure 1. Electrophoretic separation of the 5 gGH exons amplified by PCR in a 2% (w/v) agarose gel.
M: 50 bp ladder, lanes 1-3: exon 1; lanes 4-6: exon 2; lanes 7-9: exon 3; lanes 10-12: exon 4; lanes 13-15: exon 5.
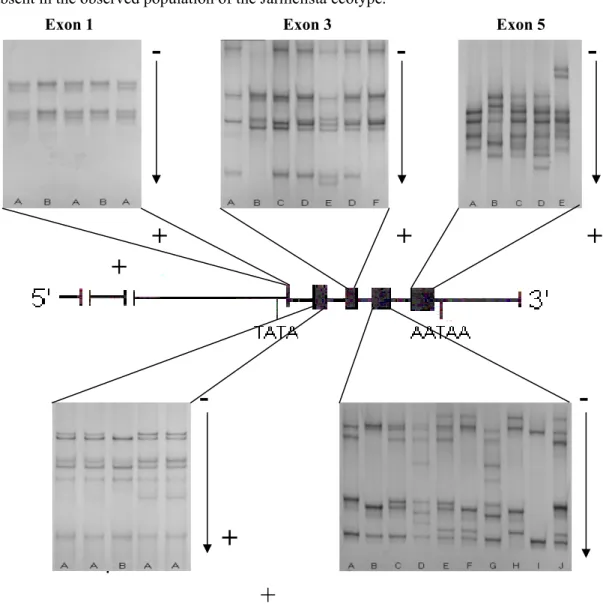
The analysis of the 229 Serrana goats shows several conformational patterns inside each amplified fragment (Figs. 2 and 3, Table 3).
One homozygous (A/A) and one heterozygous (A/B) conformational pattern were observed in exon 1 in both ecotypes. One homozygous (B/B) and one heterozygous (A/B) conformational pattern were observed in exon 2 in both ecotypes.
Exon 3 exhibits a high level of polymorphism. Six conformational patterns were observed in Jarmelista ecotype: patterns A/B and C/D have each 4 bands; 2 patterns show 6 bands -A/B/D and A/C/D; 2 patterns exhibited 8 bands - A/B/C/D and -A/B/D/E. The A/C/D pattern was not present in the observed population of Ribatejano ecotype.
In exon 4, the most polymorphic one, 10 patterns were observed in the aggregate, one homozygous (A/A) and 9 heterozygous exhibiting 4 (A/B, A/C, C/F), 6 (A/B/C, A/C/F, A/B/E, A/B/F) and 8 (A/B/D/E, A/B/C/F) bands. Jarmelista ecotype lacks pattern A/B/D/E and in the Ribatejano ecotype only five patterns were observed: A/B, A/C, A/B/C, A/B/D/E and A/B/C/F. Lastly, in exon 5, five patterns were observed with the number of bands varying from 4 to 8: pattern A/B (4 bands), pattern A/C/D (The five exons and the 5' and 3'-untranslated regions (5'-UTR and 3'-(5'-UTR) of the oGH gene were screened for mutations using PCR-single strand conformation polymorphism (PCR-SSCP) procedures in 523 Serra da Estrela ewes and were found to be highly polymorphic. The region extending across and between the GH2-N and GH2-Z copies was sequenced allowing the design of primers for the specific PCR amplification of each copy. These were cloned and sequenced in 20 animals representative of all SSCP patterns. The corresponding genotypes were established for each copy following nucleotide sequencing of SSCP alleles. Twenty-four polymorphic sites were found at the GH2-N (or GH1) and fourteen at the GH2-Z copies. Eight amino acid substitutions were predicted at the GH2-N
+
M
M 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
300 bp 200 bp 100 bp 50 bpand six at the GH2-Z copies. Milk yield adjusted to 150 lactation days was analysed for the genotype of each oGH gene copy taken separately or together (associated genotypes) by restricted maximum likelihood (REML) through a univariate best linear unbiased prediction (BLUP) animal model with repeated measures. Significant associations between genotypes and milk yield were observed. Within GH2-N genotypes there was a milk yield differential of 21.4+/-0.2 l/150 d between the most (N7) and the least (N5) productive ones. Within GH2-Z genotypes there was a differential of 21.6+/-0.2 l/150 d between the most (Z8) and the least (Z1) productive ones. The effect of associated GH2-N and GH2-Z genotypes revealed a differential of 39.6+/-0.3 l/150 d between the most (N1+Z7) and the least (N3+Z2) productive associated genotypes. The results show that GH2-N and GH2-Z genotypes significantly affect milk yield in Serra da Estrela ewes. Moreover, the apparent joint effect of GH2-N and GH2-Z genotype could improve milk yield in 25% as compared with the mean milk production of the analysed
population.
6 bands), patterns A/B/C/D, A/B/C/E and A/B/F/G (8 bands). Pattern A/B/C/E was absent in the observed population of the Jarmelista ecotype.
Figure 2. SSCP patterns for exons 1 to5 of Serrana goat GH gene separated by PAGE under
non-denaturing conditions are shown above and below the schematic representation of goat GH gene. The exons are represented by black boxes. For simplification a single letter designation was assigned to the patterns, whose correspondence with the adopted designation is shown in figure 3.
In spite of the duplication of the gGH gene that increases the complexity of the SSCP analysis, namely in the more polymorphic exons it was possible to clearly assign distinct SSCP
+
+
+
+
Exon 2 Exon 4Exon 1 Exon 3 Exon 5
- - -
+ + +
+
patterns to each exon of the gGH gene that could be used to look for associations with production parameters (see below).
A low level of polymorphism was observed in exons 1 and 2 attesting nucleotidic conservation in these regions of gGH1, gGH2 e gGH3 genes in Serrana goat. Exon 1 and its adjacent upstream region were shown to be highly conserved in ovine GH1, GH2-N e GH2-Z genes (Ofir e Gootwine 1997). Previous SSCP analysis of the gGH gene carried out on 108 Algarvia Portuguese goat (two patterns in exons 1 and 2, four in exon 3, six in exon 4 and five in exon 5 (Malveiro et al., 2001)) are comparable with the present observations performed in Serrana goat on twice the number of animals (229). Thus, the GH gene is highly polymorphic in goats and exons 4 and 5 tend to be the most polymorphic while exons 1 and 2 are more conserved. SSCP pattern A B C D E F G H I J A/B A/A Exon 1 191(83.4) 38(16.6) A/B B/B Exon 2 210(91.7) 19(8.3)
A/B C/D A/B/D A/B/C/D A/B/D/E A/C/D
Exon 3
168(73.4) 4(1.8) 26(11.4) 17(7.4) 8(3.5) 6(2.6)
A/B A/C A/B/C A/B/D/E A/B/C/F A/C/F A/B/E C/F A/A A/B/F
Exon 4
142(62.0) 13(5.7) 33(14.4) 2(0.9) 26(11.4) 2(0.9) 6(2.6) 3(1.3) 1(0.4) 1(0.4)
A/B A/C/D A/B/C/D A/B/C/E A/B/F/G
Exon 5 147(64.2) 18(7.9) 53(23.1) 3(1.3) 8(3.5) a b a b b b a b a b a b d a d b c d c d a c b d c a d b a b d e a d b e a c d c a d a b b a c a c a c a b bc a d a b e b e d a f c a b bc f a f c a c f a a b e b e a f c c f a a f a b b f a a a b b c d a a d c f g a g a f b b c d a a b d b c c e a a b b c e a b a b a a
Figure 3: Schematic representation of SSCP and proposed patterns for each exon, number of animals in
which the pattern was found and respective frequencies (in brackets).
Statistical analyses
The analysis of fat and protein yield and fat percentage reveals no differences between production traits of animals with different patterns in all studied exons. However, we found differences for milk yield in exon 2 (Jarmelista ecotype) and exon 4 (Ribatejano ecotype) and for protein percentage in exons 1 and 2 (Ribatejano ecotype).
Tables 5, 6 and 7 present the least squares means obtained for milk and protein yield. Jarmelista animals with pattern A/B in exon 2 are superior milk yielding (P<0.05) and Ribatejano animals with pattern A/B in exon 4 are also superior milk yielding(P<0.05) (Table 5). Similar results were observed for cumulative milk yield (Table 6). Ribatejano animals with pattern A/B in exon 1 and pattern B/B in exon 2 have a positive correlation with protein The five exons and the 5' and 3'-untranslated regions (5'-UTR and 3'-UTR) of the oGH gene were screened for mutations using PCR-single strand conformation polymorphism (PCR-SSCP) procedures in 523 Serra da Estrela ewes and were found to be highly polymorphic. The region extending across and between the GH2-N and GH2-Z copies was sequenced allowing the design of primers for the specific PCR amplification of each copy. These were cloned and sequenced in 20 animals representative of all SSCP patterns. The corresponding genotypes were established for each copy following nucleotide sequencing of SSCP alleles. Twenty-four polymorphic sites were found at the GH2-N (or GH1) and fourteen at the GH2-Z copies. Eight amino acid substitutions were predicted at the GH2-N and six at the GH2-Z copies. Milk yield adjusted to 150 lactation days was analysed for the genotype of each oGH gene copy taken separately or together (associated genotypes) by restricted maximum likelihood (REML) through a univariate best linear unbiased prediction (BLUP) animal model with repeated measures. Significant
associations between genotypes and milk yield were observed. Within GH2-N genotypes there was a milk yield differential of 21.4+/-0.2 l/150 d between the most (N7) and the least (N5) productive ones. Within GH2-Z genotypes there was a differential of 21.6+/-0.2 l/150 d between the most (Z8) and the least (Z1) productive ones. The effect of associated GH2-N and GH2-Z genotypes revealed a differential of 39.6+/-0.3 l/150 d between the most (N1+Z7) and the least (N3+Z2) productive associated genotypes. The results show that GH2-N and GH2-Z genotypes significantly affect milk yield in Serra da Estrela ewes. Moreover, the apparent joint effect of GH2-N and GH2-Z genotype could improve milk yield in 25% as compared with the mean milk production of the analysed population.
percentage(Table 7). In the analyses of the fat and protein content, the unstructured covariance matrix allowed the best fit.
Table 5: Least squares means ( S.E.) for milk production associated with patterns of exon 2 (Jarmelista ecotype) and exon 4 (Ribatejano ecotype) of Serrana goat breed.
Pattern Exon 2 Pattern Exon 4
A/B 173.9 b 13.5 A/B 252.3 b 16.9
B/B 118.9 a 28.8 A/C 228.7 ab 51.6
A/B/C 190.4 a 25.1
A/B/D/E 220.5 ab 59.6
A/B/C/F 210.4 ab 25.8
a, b; means with like lower cases do not differ (P>0.05).
Table 6: Least squares means ( S.E.) for cumulative milk production associated with patterns of exon 2 (Jarmelista ecotype) and exon 4 (Ribatejano ecotype) of Serrana goat breed.
A/B 221.1 b 5.4 A/B 299.9 b 6.9
B/B 177.8 a 14.1 A/C 308.3 b 19.8
A/B/C 259.7 a 14.4
A/B/D/E 321.2 ab 48.1
A/B/C/F 265.8 ab 24.7
a, b; means with like lower cases do not differ (P>0.05).
Table 7: Least squares means ( S.E.) for protein content associated with patterns of exon 1 and 2 Serrana goat breed Ribatejano ecotype.
Pattern Exon 1 Pattern Exon 2
A/B 3.58 b 0.193 A/B 3.47 a 0.191
A/A 3.18 a 0.225 B/B 3.83 b 0.236
The present results reinforce the idea that SSCP analysis is a valuable approach for the establishment of allelic variants in genes that express hormones associated with productive traits. In particular, the search of genetic markers located in the gGH gene is within the scope of this method. Indeed SSCP polymorphisms that exert a positive influence in milk production have previously been established at the GH gene from the caprine Algarvia breed (Malveiro et
al., 2001). Namely, polymorphisms in exons 4 and 5 of this gene were shown to be associated
with that trait. The results here presented concerning Serrana goat, the most representative indigenous Portuguese caprine breed, were obtained with twice the number of animals than that of the Algarvia breed. They corroborate the existence of an association of polymorphisms in exon 4 of the gGH gene with milk production. Moreover, polymorphisms in exon 2 are also associated with this trait and polymorphisms in this exon and in exon 1 are associated with protein content.
The high SSCP polymorphism observed in the gGH raises the question of their importance and significance. Do these polymorphisms correspond to silent mutations or to actual amino acid mutations and are these mutations implicated in structural modifications that influence the biological activity of the hormone? In this regard it is interesting to notice that several aminoacids coded by exons 2 and 4 are part of the sites of GH that are involved in its binding with GH receptor (GHR)/GH binding protein (GHBP) (Cunningham et al., 1991).
Exon 4 appears to be the most polymorphic region in goat breeds. Sequencing of patterns corresponding, firstly to this region and then to the other exons is underway. The information thus obtained would allow the characterisation of putative mutations of amino acid residues of the gGH, eventually involved in variations of milk yield and in GH binding to its receptor.
4.
Conclusions
SSCP polymorphisms can be rapidly and inexpensively detected in a population of dairy animals and their associations with milk traits studied. In spite of gene duplication it is possible to genotype variants of the gGH gene using the PCR-SSCP technique. We have applied this approach to look for associations between polymorphisms observed in exons 1-5 of the GH gene from the autochthonous Portuguese caprine Serrana breed and milk traits. Milk production is under the influence of polymorphisms detected in exons 4 and 2 and protein content is under the influence of polymorphisms detected in exons 1 and 2. The positive influence of exon 4 in milk production established in this study confirms previous findings in caprine Algarvia breed suggesting that this region of the gGH gene is particularly important and deserves further studies.
Molecular and structural characterization of the GH genetic variants should allow the implementation of fast analytic procedures based on specific PCR directed to mutations in exons where gGH genetic PCR-SSCP patterns are likely to be associated with milk traits. The application of these procedures to larger populations is expected to lead to the establishment of highly significant association between the detected polymorphism and milk traits.
Acknowledgments
We acknowledge Fundação para a Ciência e Tecnologia (FCT) (PRAXIS XXI 3/3.2/CA/1991/95) for financial support and Associação de Criadores de Reprodutores de Gado Oeste" (ACRO), Associação de Criadores de Caprinos e Ovinos do Ribatejo Oeste" (ACORO) and "Associação de Criadores da Guarda" (AGRIGUARDA). for providing blood samples and zootechnical data. I.C.S. and M. R. M. thank FCT for their PRAXIS XXI Ph.D. grants (BD/18061/98 and BD/1140/2000, respectively).
REFERENCES
-Chikuni, K., Terada, F., Kageyama, S., Koishikawa, T., Kato, S., and Ozutsumi, K. (1991). Identification of DNA sequence variants for amino acid residue 127 of bovine growth hormone using the polymerase chain chain reaction method. Animal Science and Technology, 62: 660-666.
-Chrenek, P., Kmet, J., Sakowski, T., Vasicek, D., Huba, J. and Chrenek, J. (1998) Relationships of growth hormone genotypes with meat production traits of Slovak Pied bulls.
Czech J. Anim. Sci. 43: 541-544.
-Cunningham, B.C., Ultsch, M., Abraham, M. de Vos, Mulkerrin, M.G., Clauser, K.R. and Wells, J.A. (1991). Dimerization of the extracellular domain of the human growth hormone receptor by a single hormone molecule. Science, 254: 821-825.
-Falaki, M., Sneyers, M., Prandi, A., Massart, S., Corradini, C., Formigoni, A., Burny, A., Portetelle, D., Renaville, R., (1996). TaqI growth hormone gene polymorphism and milk production traits in Holstein-Friesian cattle. Anim. Sci. 63, 175-181.
-Falaki, M., Prandi, A., Corradini, C., Sneyers, M., Gengler, N., Massart, S., Fazzini, U., Burny, A., Portetelle, D. and Renaville, R. (1997). Relationships of growth hormone gene and milk protein polymorphisms to milk production traits in Simmental cattle. J. Dairy Res. 64: 47-56. -Fugita, K., Silver J., Peden, K., (1992). Changes in both gp120 and gp41 can account for increased growth potential and expanded host range of human immunodeficiency virus type 1.
J. Virol. 66, 4445-4452.
-Gion J-M, Rech P, Grima-Pettenati J, Verhaegen D, Plomion C, (2000), "Mapping candidate genes in Eucalyptus with emphasis on lignification genes", Molecular Breeding, 6: 441-449. -Grochowska, R., Sorensen, P., Zwierzchowski, L., Snochowski, M., Lovendahl, P. (2001) Genetic variation in stimulated GH release and in IGF-I of young dairy cattle and their associations with the leucine/valine polymorphism in the GH gene. J. Anim. Sci. 79(2): 470-476.
-Hart, I.C., Bines, J.A., Morant, S.V., 1980. The secretion and metabolic clearance rates of growth hormone, insulin and prolactin in high and low yielding cattle at four stages of lactation.
Life Sci. 27, 1839-1847.
-Høj, S., Fredholm, M. Larsen, N.J. and Nielsen, V.H. (1993) Growth hormone gene polymorphism associated with selection for milk fat production in lines of cattle. Anim. Genet. 24: 91-96.
-Kioka, N., Manabe, E., Abe, M., Hashi, H., Yato, M., Okuno, M., Yamano, Y., Sakai, H., Komano, T., Utsumi, K. and Iritani, A. (1989) Cloning and sequencing of goat growth hormone gene. Agric. Biol. Chem. 53: 1583-1587.
-Kirkpatrick, B.W. (1992) Detection of three-allele single strand conformation polymorphism (SSCP) in the fourth intron of the bovine growth hormone gene. Anim. Genet. 23: 179-181.
-Lagziel, A., Lipkin, E. and Soller, M. (1996) Association between SSCP haplotypes at the bovine growth hormone gene and milk protein percentage. Genetics 142: 945-951.
-Lagziel, A. and Soller, M. (1999). DNA sequence of SSCP haplotypes at the bovine growth hormone (bGH) gene. Anim. Genet. 30: 362-365.
-Lee, B.K., Lin, G.F., Crooker, B.A., Murtaugh, M.P., Hansen, L.B. and Chester-Jones, H. (1996). Association of somatotropin (BST) gene polymorphism at the 5th exon with selection for
milk yield in Holstein cows. Domest. Anim. Endocrinol. 13:373-381.
-Lucy, M.C., Hauser, S.D., Eppard, P.J., Krivi, G.G. and Collier, R.J. (1991). Genetic polymorphism within the bovine somatotropin (bST) gene detected by polymerase chain reaction and endonuclease digestion. J. Dairy Sci.74(Suppl. 1): 284. (Abstr.).
-Lucy, M.C., Hauser, S.D., Eppard, P.J., Krivi, G.G., Clark, J.H., Bauman, D.E. and Collier, R.J. (1993) Variants of somatotropin in cattle: gene frequencies in major dairy breeds and associated milk production. Domest. Anim. Endocrinol. 10(4): 325-333.
-Malveiro, E., Pereira, M., Marques, P.X., Santos, I.C., Belo, C., Renaville, R. and Cravador, A. (2001) Polymorphisms at the five exons of the growth hormone gene in the Algarvia goat: possible association with milk traits. Small Rumin. Res. 41: 163-170.
-Morohoshi, F., Hayashi, K., Munakata, N., 1991. Molecular analysis of Bacillus subtilis ada mutants deficient in the adaptative response to simple alkylating agents. J. Bacteriol. 173, 7834-7840.
-Neibergs, H., Dietz, A., Womack, J., 1993. Single strand conformation polymorphisms (SSCP) detected in five bovine genes. Anim. Genet. 24: 81-84.
-Ofir, R. and Gootwine, E. (1997) Ovine growth hormone gene duplication – structural and evolutionary implications. Mamm. Genome 8: 770-772.
-Orita, M., Iwahana, H., Kanazawa, H., Hayashi, K. and Sekiya, T. (1989) Detection of polymorphisms of human DNA by gel electrophoresis as single-strand conformation polymorphisms. Proc. Natl. Acad. Sci. USA. 86: 2766-2770.
Pravenec, M., Simonet, L., Kren, V., St.Lezin, E., Levan, G., Szpirer, J., Szpirer, C., Kurtz, T., (1992). Assignment of rat linkage group V to chromosome 19 by single strand conformation polymorphism analysis of somatic cell hybrids. Genomics 12, 350-354.
-Peel, C., Bauman, D.E., (1987). Somatotropin and lactation. J. Dairy Sci. 70, 474-486.
-Reinecke, R.L., Barnes, M.A., Akers, R.M. e Pearson, R.E. (1993). Effect of selection for milk yield on lactation performance and plasma growth hormone, insulin and IGF-I in first lactation Holstein cows. J. Dairy Sci. 76(Suppl.1): 286.
-Renaville, R., Massart, S., Shojae, D., Prandi, A., Devolder, A., Sneyers, M., Falaki, M., Burny, A. and Portetelle, D. (1995) Growth hormone. insulin-like growth factor-I and their plasma binding proteins in different cattle breeds. Zoot. Nutr. Anim. 21: 75-84.
-Sabour, M.P., Lin, C.Y. and Smith, C. (1997). Association of genetic variants of bovine growth hormone with milk production traits in Holstein cattle. J. Anim. Genet. 114: 435-442.
-Schlee, P., Graml, R., Schallenberger, E., Schams, D., Rottmann, O., Olbrich-Bludau, A., Pirchner, F., (1994a). Growth hormone and insulin-like growth factor I concentrations in bulls of various growth hormone genotypes. Theor. Appl. Genet. 88, 497-503.
-Schlee, P., Graml, R., Rottmann, O. and Pirchner, F. (1994b). Influence of growth hormone genotypes on breeding values of Simmental bulls. J. Anim. Breed. Genet. 111: 253-256.
-Shariflou, M.R., Moran, C. and Nicholas, F.W. (2000). Association of the Leu (127) variant of the bovine growth hormone (bGH) gene with increased yield of milk, fat, and protein in Australian Holstein-Friesians. Australian J. Agricultural Research. 51: 515:522.
-Unanian, M.M., DeNise, S.K., Zhang, H.M., and Ax, R.L. (1994). Rapid Communication: Polymerase chain reaction – Restriction fragment length polymorphism in the bovine growth
hormone gene. J. Anim. Sci. 72: 2203.
-Yao, J., Aggrey, S.E., Zadworny, D., Hayes, J.F. and Kuhnlein, U. (1996). Sequence variations in the bovine growth hormone gene characterized by single-strand conformation polymorphism (SSCP) analysis and their association with milk production traits in Holsteins. Genetics 144: 1809-1816.
-Valinsky, A., Shani, M. and Gootwine, E. (1990). Restriction fragment length polymorphism in sheep at the growth hormone locus is the result of variation in gene number. Anim. Biotechnol. 1(2): 135-144.
-Vasilatos, R., Wangsness, P.J., (1981). Diurnal variations in plasma insulin and growth hormone associated with two stages of lactation in high producing dairy cows. Endocrinology 108, 300-304.
-Vukasinovic, N., DeNise, S.K. and Freeman, A.E. (1999). Association of growth hormone loci with milk yield traits in Holstein bulls. J. Dairy Sci. 82: 788-794.
-Yamano, Y., Abe, M., Mikawa, S., Kioka, N., Manabe, E., Sakai, H., Komano, T., Utsumi, K., and Iritani, A. (1991). Structural analysis of repetitive DNA sequences in the goat growth hormone gene region, Agric. Biol. Chem., 55(3):633-639.
-Zhang, H.M., Brown, D.R., Denise, S.K., Ax, R.L., (1992). Nucleotide sequence determination of a bovine somatotropin allele. Anim. Genet. 23, 578.
-Zhang, H.M., Brown, D.R., DeNise, S.K. and Ax, R.L. (1993.) Rapid Communication: Polymerase chain reaction – Restriction fragment length polymorphism analysis of the bovine somatotropin gene. J. Anim. Sci. 71: 2276.
Bauman D.E. 1999. Somatotropin mechanism in lactating cows: from basic science to commercial application. Dom. Anim. Endocrinol. 17, 101-116.
Baldi, A. (1999) Manipulation of milk production and quality by use of somatotropin in dairy ruminants other than cow, Dom. Anim. Endocrinol. 17, 131-137.
Breier B.H. (1999) Regulation of protein and energy metabolism by the somatotropic axis.
Dom. Anim. Endocrinol. 17: 209-218.
Scaramuzzi RJ, Murray JF, Downing JA, Campbell BK.The effects of exogenous growth hormone on follicular steroid secretion and ovulation rate in sheep.