Identification d'effecteurs du pouvoir pathogène et de voies métaboliques chez l'oomycète Aphanomyces euteiches par une approche génomique
127
0
0
Texte intégral
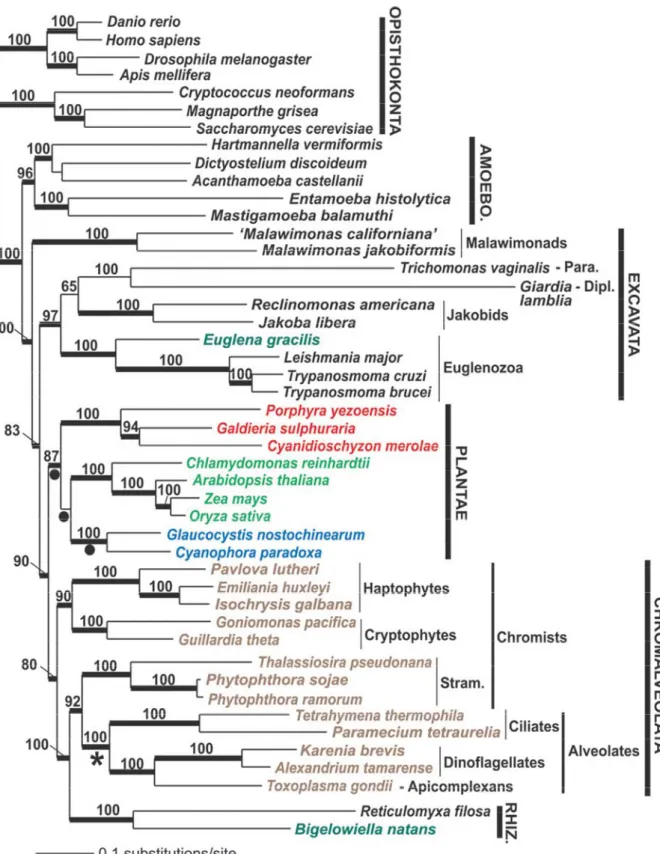
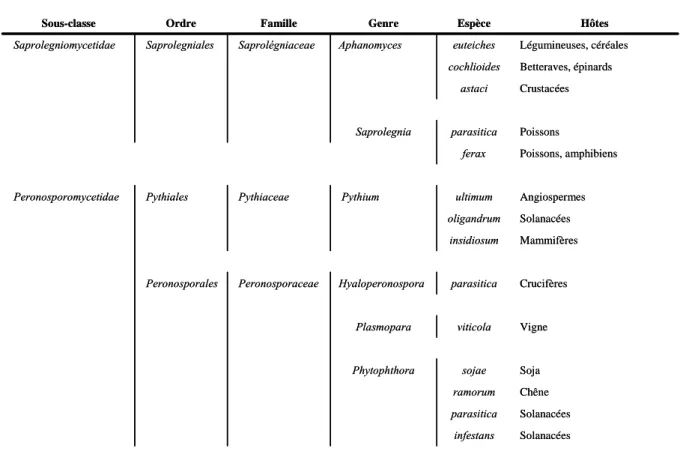
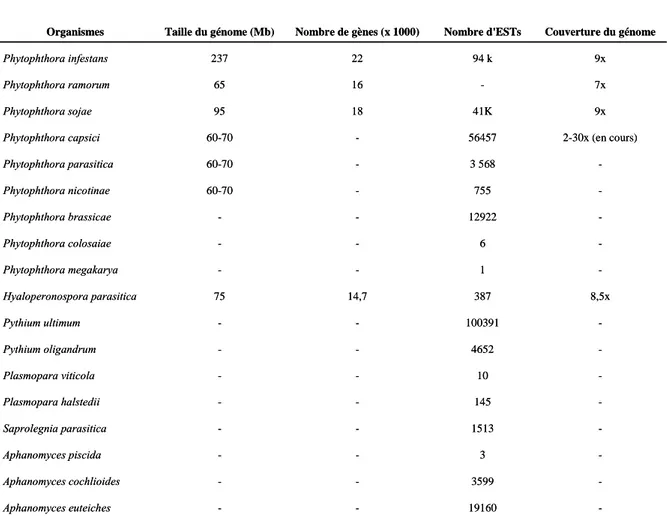
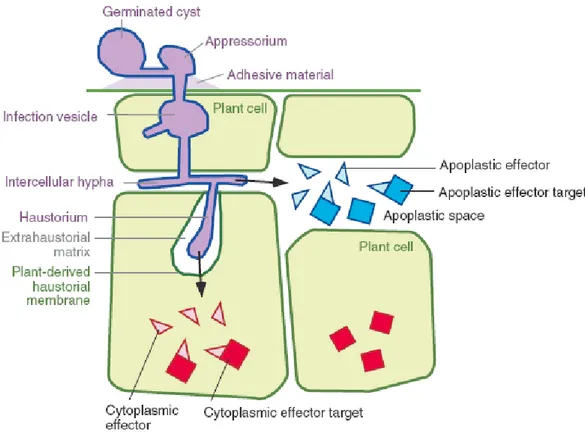
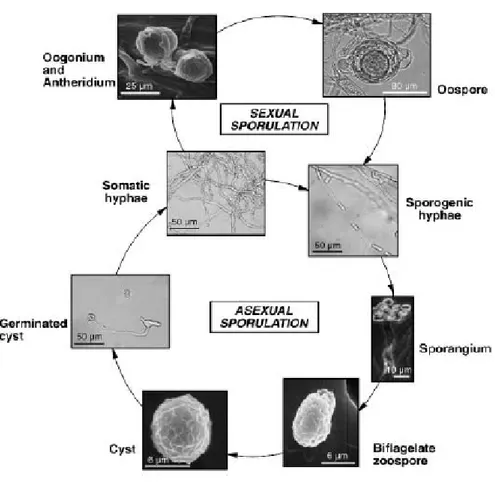
Figure

+7
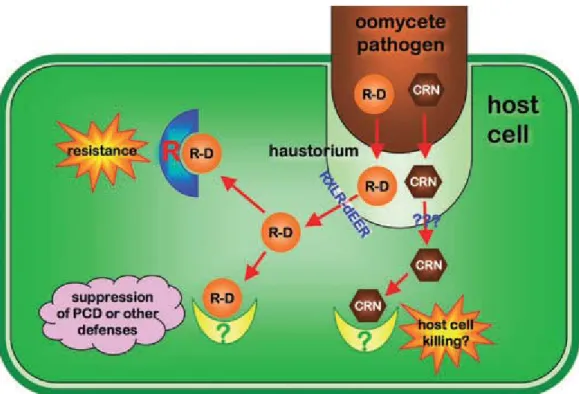
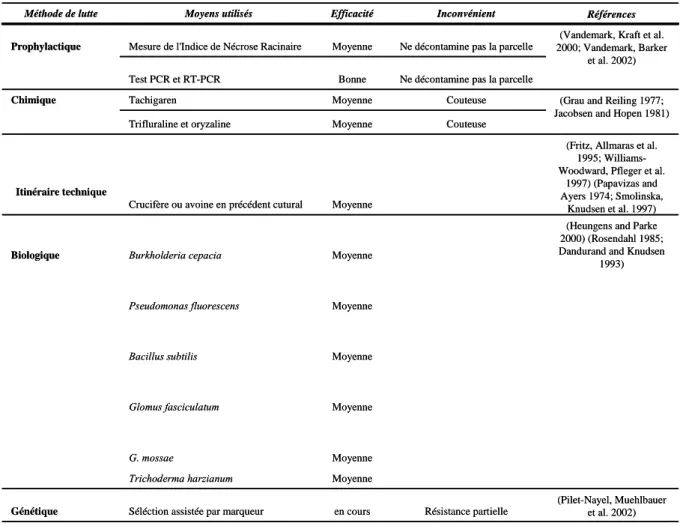
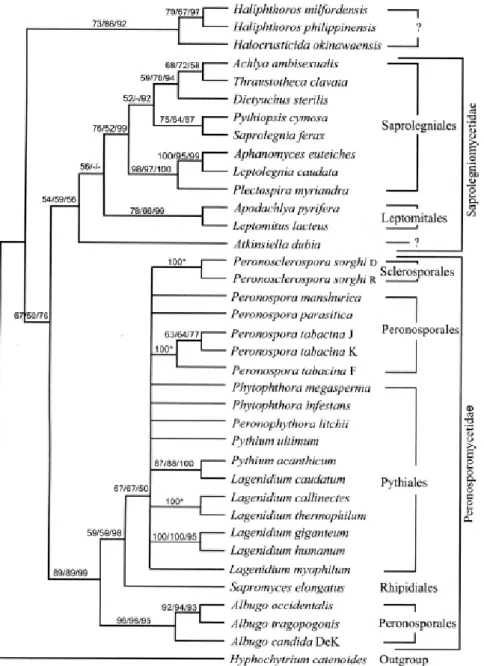
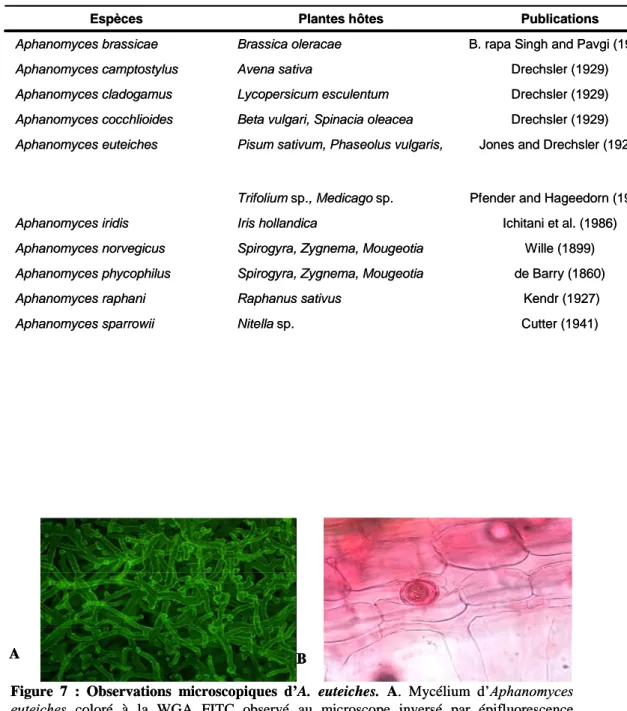
Documents relatifs
