Rôle de l’axe CD40L/CD40 dans les cellules endothéliales progénitrices
Texte intégral
Figure
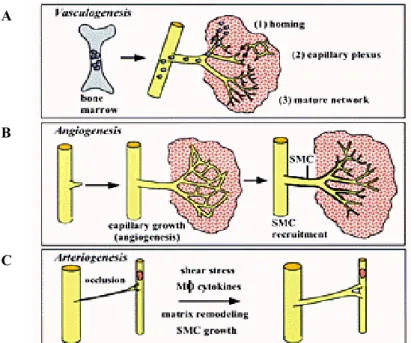
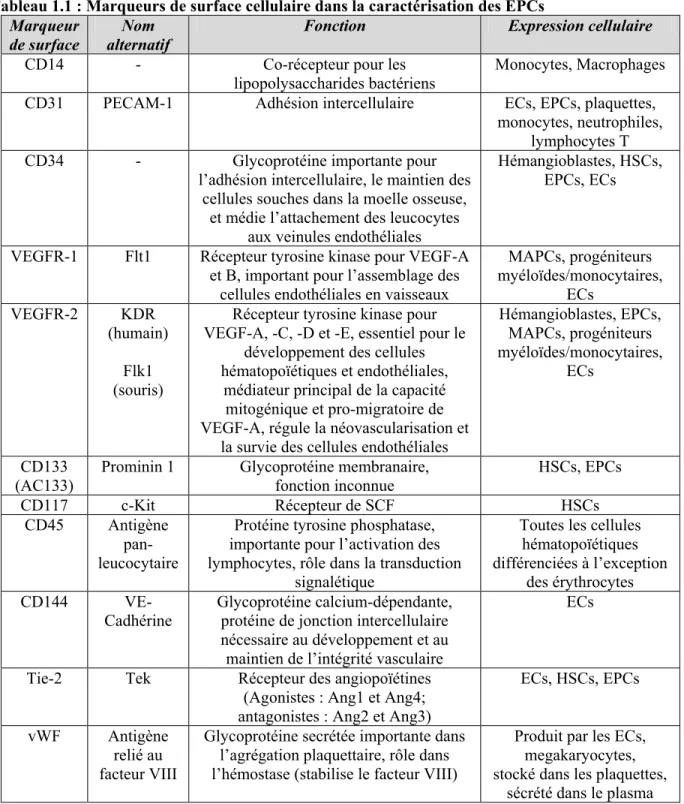

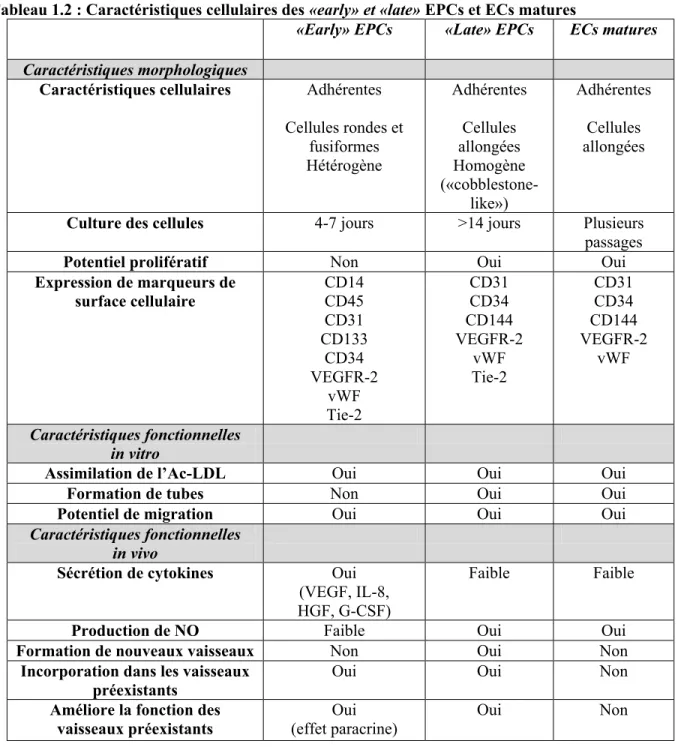
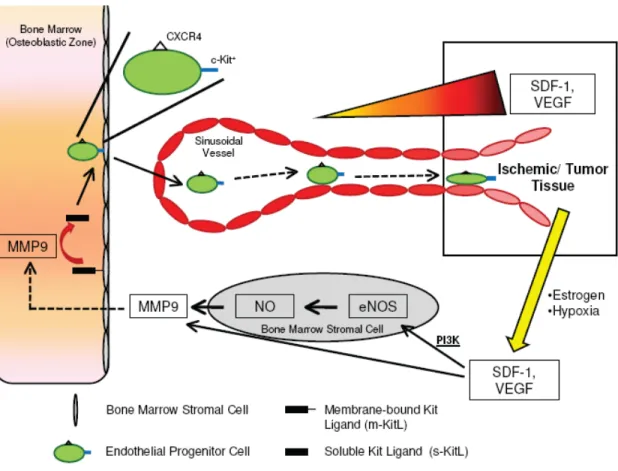


Outline
Documents relatifs
(D) Les cellules Raji ont été prétraitées pendant 30 min. Les résultats représentent une moyenne d'un triplicata d'une expérience répétée trois fois.. Ainsi le fait que
Curieusement, nous avons observé que la petite GTPase Rap1 est essentielle, suite à une stimulation par l’Ang-1, pour la stabilisation des JA ainsi que pour le
leurs interactions avec les polymorphonucléaires neutrophiles entraÎnent une désorganisation et une altération de la paroi vasculaire caractérisées par une
Cette dif férence morphologique s'accompagne d'une différence fonctionnelle avec notamment l'expression à leur surface apicale de molécules d'adhérence impliquées
Ainsi, pour étudier, in vivo, le rôle des cellules endothéliales (CE) JAK2V617F dans la physiopathologie de l’augmentation de l’angiogenèse des NMP, il a
Des analyses immunohistochimiques étudiant les cellules progénitrices hépatiques (totales et en prolifération), les hépatocytes en prolifération et l’expansion des macrophages
GH JDXFKH O8SULJKW &UHHN R QRXV FDPSRQV &HWWH SUHPLqUH pWDSH QRXV SHUPHW GH FRQVWDWHU TXH OH SODQ GH FKHYDXFKHPHQW GH OD QDSSH PDUTXp SDU OH FRQWDFW GHV TXDUW]LWHV GX
/H 0DOP VXSpULHXU GH OD 1DSSH GH 0RUFOHV HW GH O$XWRFKWRQH HVW IRUPp SDU XQH pSDLVVH VpULH GH FDOFDLUHV PDVVLIV QRLUV j JUDLQ WUqV ILQ GpSRVpV HQ PLOLHX WUDQTXLOOH UHODWLYHPHQW
