Binding of Zearalenone, Aflatoxin B1, and Ochratoxin A by Yeast-Based Products: A Method for Quantification of Adsorption Performance
12
0
0
Texte intégral
Figure


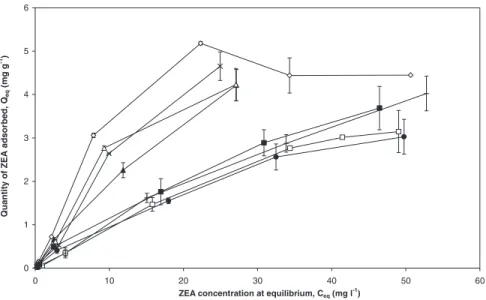
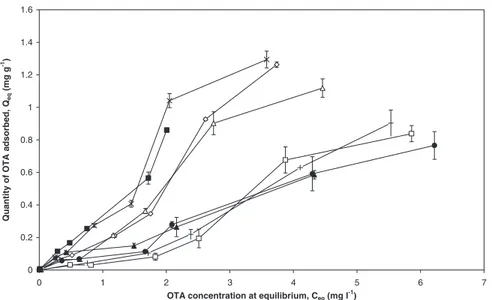
+4
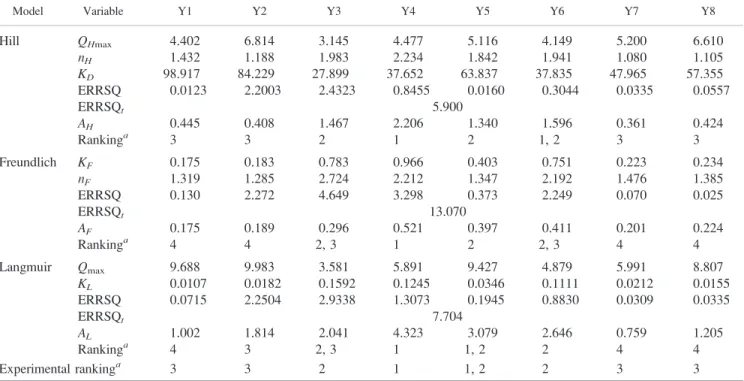
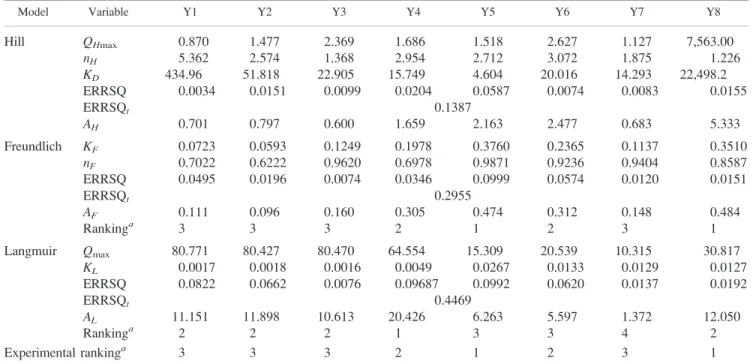

Documents relatifs