Étude des déterminants épigénétiques de facteurs de risque de la maladie cardiovasculaire
Texte intégral
Figure
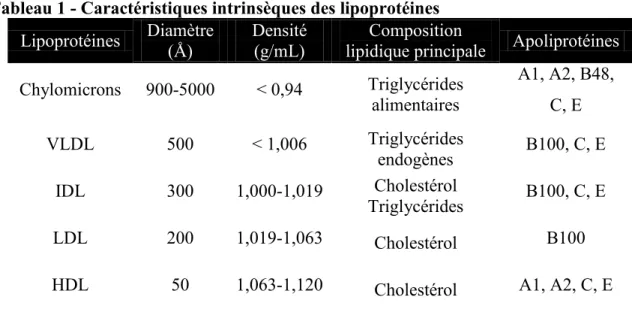
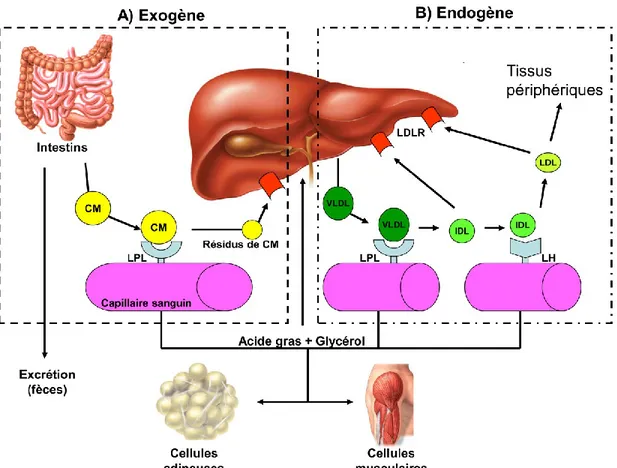
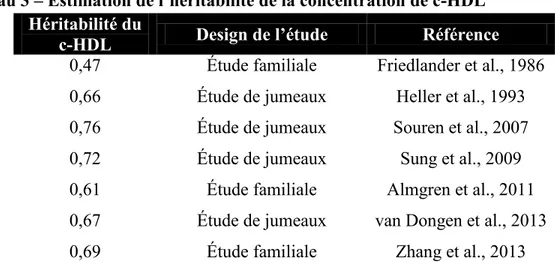
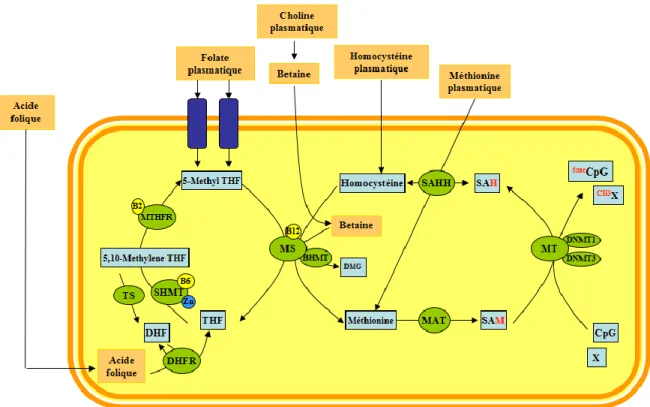
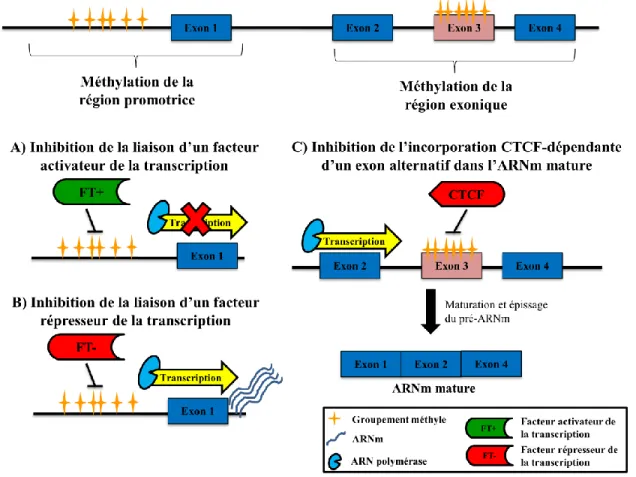
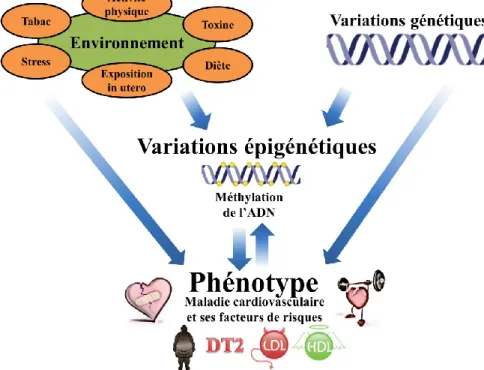

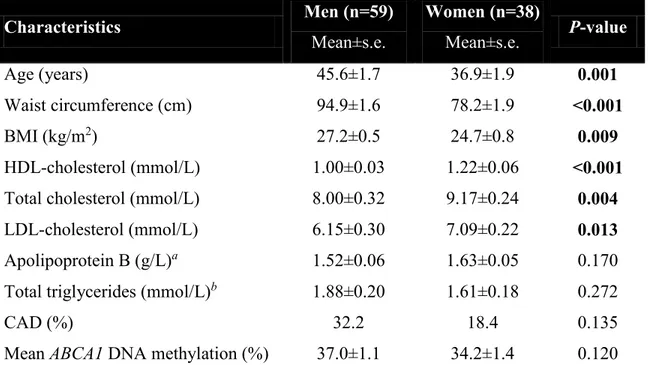
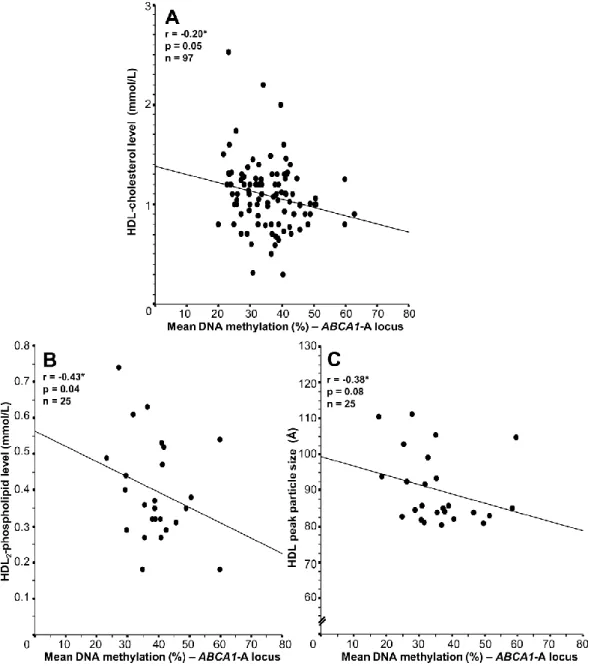
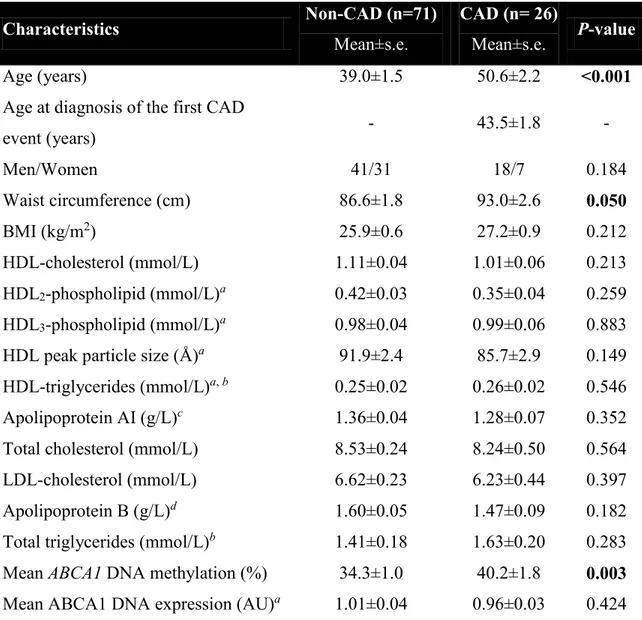
Documents relatifs
Ainsi, la cohorte E3N a permis de mettre en évidence les possibilités et les limites de prise en compte de l’évaluation de la santé des personnes âgées par une
Bien que les élévations des D-dimères reconnaissent une étiologie multifactorielle, nos résultats nous amèneraient à conclure que les élévations des D-dimères
Cette étude peut résonner au sein des CEMPN sur la prise en charge des personnels navigants, et donner lieu à d’autres études plus analytiques par exemple sur l’intérêt
En raiso n des limites propres à ce type d’étude, il était nécessaire de confirmer ces résultats dans une étude en population de plus grande ampleur, raison pour laquelle nous avons
L'objectif de l'étude était de déterminer le risque de la MTEV lié aux polymorphismes génétiques (G1691A du facteur V Leiden, G20210A de la prothrombine, C677T
Même si la santé des salariés et les caractéristiques propres à chaque individu (âge, sexe, corpulence....) peuvent moduler le niveau d’exposition, les
Pour progresser dans la prévention de la démence, il est important d’identifier les facteurs modifiables qui agissent en amont, c’est-à-dire ceux qui
La plupart des producteurs savent que des taux plus élevés de prostaglandine nuisent à la saine fixation du corps jaune et au bon déroulement de la gestation, et peuvent entraîner
