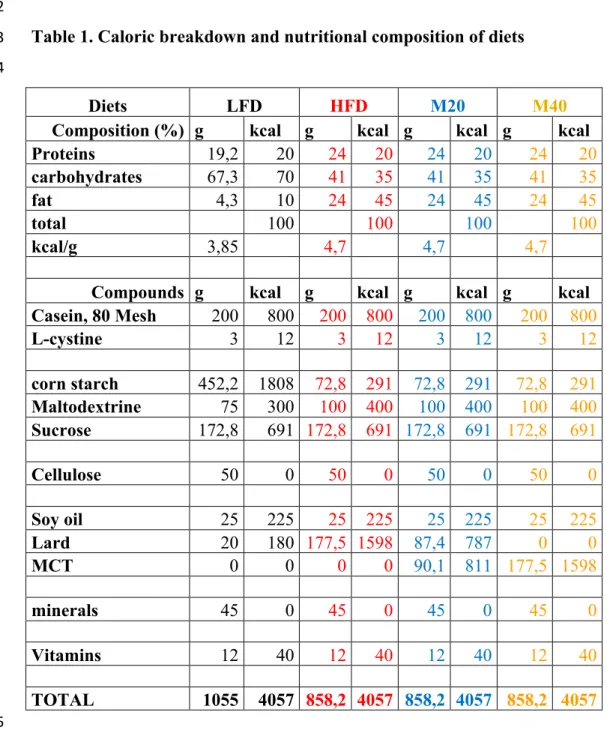
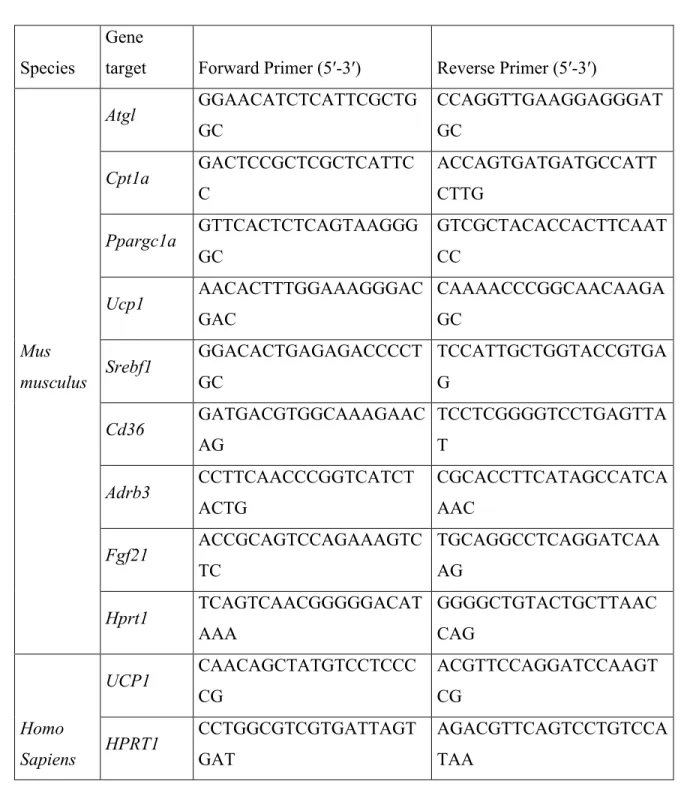
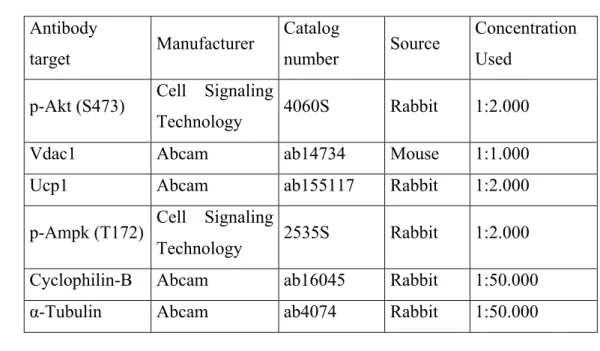
A high-fat diet enriched in medium chain triglycerides triggers hepatic thermogenesis and improves metabolic health in lean and obese mice
44
0
0
Texte intégral
Figure
Documents relatifs