DÉTERMINANTS DU RECRUTEMENT LOCAL ET DE LA DISPERSION NATALE CHEZ UNE POPULATION D’HIRONDELLE BICOLORE
par
Esther Carle-Pruneau
Mémoire présenté au Département de biologie en vue de l’obtention du grade de maître ès sciences (M.Sc.)
FACULTÉ DES SCIENCES UNIVERSITÉ DE SHERBROOKE
Le 1er avril 2021
Le jury a accepté le mémoire de Esther Carle-Pruneau dans sa version finale
Membres du jury
Professeur Dany Garant Directeur de recherche Département de biologie Université de Sherbrooke
Professeur Marc Bélisle Codirecteur de recherche
Département de biologie Université de Sherbrooke
Professeure Fanie Pelletier Évaluatrice interne Département de biologie Université de Sherbrooke
Professeur Mark Vellend Président-rapporteur Département de biologie Université de Sherbrooke
REMERCIEMENTS
J’aimerais envoyer un énorme merci à mon directeur Dany Garant pour son encadrement et sa confiance. Sous ta direction je n’ai jamais senti de pression et je me suis sentie libre de développer ce projet en lien avec mes intérêts. Merci aussi pour ta compréhension envers les différentes étapes difficiles que j’ai dû surmonter spécifiquement cette dernière année. Un merci semblable à mon co-directeur, Marc Bélisle. Comme plusieurs autres étudiants ont déjà mentionné, merci pour les longues discussions passées dans ton bureau à se demander du comment au pourquoi et pour les conversations animées durant le midi. Merci aux membres de mon comité, Fanie Pelletier et Mark Vellend, pour leurs précieux conseils, encouragements et soutien. Je remercie François Rousseu pour son aide en analyse statistique et en codage ainsi que pour le temps précieux que tu m’as accordé. Un énorme merci aux différentes personnes qui ont croisé mon chemin durant les 3 années de terrain d’Hirondelle. Un merci bien spécial et rempli d’amour à mes girls, Ève Courtois et Audrey Sigouin. Vous êtes des femmes fortes et intelligentes à plein de niveaux. Vous avez marqué autant mon cheminement professionnel que personnel. Un immense merci aux membres du labo Garant, spécifiquement à Chelsey, Carolyne et Philippine pour votre immense support et aide tout au long de ma maîtrise. Je remercie également tous les autres membres des labos Bélisle, Pelletier et Festa-Bianchet pour les beaux moments de rire et de folie passés en votre compagnie. Un merci spécial à ma famille. Cette dernière année de maîtrise a été riche en émotions et je vous remercie d’avoir simplement été là. Un immense merci à ma mère qui a été un gros pilier pour passer au travers cette épreuve que la vie a décidé de mettre sur mon chemin. Sans toi, il aurait été difficile de garder la motivation de continuer. Merci à mon beau-père pour ton soutien et tes nombreuses questions sur la recherche, c’est toujours un plaisir de te répéter à chaque fois ce que je fais. Un doux merci à mon copain de me donner autant d’amour et de soutien dans ce que je fais et d’être présent. Finalement, je remercie mon père. Malgré ta lourde absence, merci papa d’avoir toujours cru en moi et d’avoir été aussi fier de ta ti-fille. Je ressens encore cette fierté aujourd’hui à tout moment et ça fait grandement du bien, je t’aime.
SOMMAIRE
L’imprévisibilité engendrée par les perturbations environnementales anthropiques amène une forte pression sur les comportements des espèces sauvages qui exploitent les habitats touchés par ces perturbations. L’intensification agricole est une de ces perturbations. Celle-ci s’est accrue depuis les années 1970 et a mené, entre autres, à l’ajout d’intrants chimiques dans l’environnement, à l’altération des habitats et à l’utilisation de machineries lourdes. Ces pratiques sont actuellement largement présentées dans la littérature comme étant des causes potentielles du déclin observé chez les insectivores aériens. Pour des espèces qui exploitent plusieurs aires géographiques durant leur cycle de vie, telle que les espèces migratrices, l’effet additif de ces pratiques sur ces différents territoires peut accentuer les conséquences néfastes sur les individus. Ces pratiques sont donc sujettes à affecter la survie des juvéniles, leurs mouvements et par conséquent, leur recrutement dans les populations.
L’objectif de cette étude est donc d’évaluer les déterminants du recrutement local et de la dispersion natale chez l’Hirondelle bicolore (Tachycineta bicolor), un insectivore aérien migrateur qui présente des déclins importants de ses populations nichant dans le Nord-Est de l’Amérique du Nord. Chez cette espèce, ces processus restent méconnus et nécessitent une attention particulière afin d’améliorer nos connaissances sur la dynamique spatiale de ces populations. J’aborde ces aspects grâce au suivi d’une population d’Hirondelle bicolore depuis 2004 et ce, au sein d’un système d’étude composé de 40 fermes situées le long d’un gradient d’intensification agricole du Sud du Québec.
Mes résultats indiquent que le recrutement est faible dans le système d’étude et qu’il est fortement influencé par les traits des juvéniles. Mes résultats montrent aussi que les distances de dispersion natale au sein du système sont généralement faibles et influencées par différents facteurs selon le sexe. Les femelles utiliseraient l’information sociale autour de leur ferme de naissance, tandis que les mâles seraient influencés par la présence de compétiteurs et l’âge maternel. Par ailleurs, la sélection des fermes de reproduction semble s’effectuer à de multiples
vi échelles spatiales. À grandes échelles, elle favorise le recrutement au sein de fermes possédant des caractéristiques similaires à celles des fermes de naissance, alors qu’à plus petites échelles, elle est aléatoire et semble dépendre de facteurs de dispersion, tels que la densité de congénère et de compétiteur inter-spécifique, plutôt que des caractéristiques environnementales.
Mes résultats apportent un éclairage sur les mécanismes qui régulent les processus de la dynamique de cette population d’Hirondelle bicolore. Les faibles taux de recrutement au sein de cette population en diminution d’effectifs montrent notamment l’importance des juvéniles dans la dynamique de populations en déclin. Ce résultat souligne en effet la nécessité d’améliorer nos connaissances sur les facteurs potentiels influençant la survie des juvéniles à l’extérieur des aires de reproduction. De plus, comme les recrues semblent disperser près de leur site de naissance et rechercher des habitats de reproduction semblables à leur habitat de naissance, cela renforce l’idée qu’il faut accroître les efforts de gestion non seulement sur les sites d’arrêts migratoires ou d’hivernage, mais aussi au niveau des aires de reproduction de cette espèce afin de maintenir le recrutement. À ce titre, les limites et les perspectives futures de ma recherche sont discutées en détail dans le troisième chapitre de ce mémoire.
Mes recherches montrent l’importance de la survie des juvéniles et de leur comportement de dispersion dans la dynamique des populations aviaires, et ce spécialement dans un contexte de changements environnementaux. Le recrutement local étant un taux vital important dans le maintien et la croissance des populations, les changements anthropiques rapides du paysage risquent d’affecter le succès reproducteur des individus et d’augmenter les coûts reliés à la dispersion. Ceci montre l’importance de mener plus d’étude sur les causes potentielles de survie et au moment que celles-ci agissent afin d’améliorer, entre autres, nos efforts de gestion chez les espèces en déclin.
Mots clés : dispersion natale, dynamique des populations, espèce migratrice, habitat natal, Hirondelle bicolore, intensification agricole, recrutement local
TABLE DES MATIÈRES
CHAPITRE 1 INTRODUCTION ... 1
1.1 Dynamique des populations et recrutement local ... 1
1.1.1. Survie des juvéniles ... 3
1.1.2. La dispersion natale ... 6
1.1.2.1. Variabilité de la dispersion natale ... 6
1.1.2.2. Facteurs de dispersion liés à l’écologie……….8
1.1.2.3. Facteurs de dispersion liés au phénotype……….10
1.1.3. Sélection d’habitat ... 12
1.1.3.1. Le comportement de prospection………13
1.1.3.2. Sélection d'habitat non aléatoire……….13
1.1.3.3. Sélection densité-dépendante………..14
1.1.3.4. Sources d'information lors de la sélection d'habitat……...…….……15
1.1.3.5. La sélection d'habitat à multiple échelle……….17
1.2 Objectif et importance du projet ... 18
1.3 Méthodologie générale ... 18
1.3.1. Espèce à l’étude ... 18
1.3.2. Aire d’étude ... 20
1.4 Hypothèses et prédictions ... 20
CHAPITRE 2 DÉTERMINANTS DU RECRUTEMENT LOCAL ET DE LA DISPERSION NATALE CHEZ UNE POPULATION DE PASSEREAUX EN DÉCLIN ... 23
2.1 Introduction à l’article ... 23
2.2 Abstract ... 25
2.3 Introduction ... 26
2.4 Methods ... 31
viii
2.4.2. Monitoring of nestlings and adults ... 32
2.4.3. Landscape characterization and nest-box occupancy ... 33
2.4.4. Statistical analyses ... 33
2.4.5. Local recruitment ... 34
2.4.6. Natal dispersal distance ... 35
2.4.7. Natal vs breeding habitat difference ... 36
2.5 Results ... 37
2.5.1. Local recruitment ... 37
2.5.2. Natal dispersal ... 41
2.5.3. Natal vs breeding habitat differences ... 44
2.6 Discussion ... 49
2.6.1. Local recruitment ... 49
2.6.2. Natal dispersal ... 51
2.6.3. Natal vs breeding habitat difference ... 54
2.7 Conclusion ... 55
2.8 References ... 56
2.9 Supplementary material ... 67
CHAPITRE 3 DISCUSSION ET CONCLUSION ... 74
3.1 Retour sur les objectifs et résultats ... 74
3.1.1. Recrutement local ... 76
3.1.2. Dispersion natale ... 77
3.1.2.1. Mâles……….78
3.1.2.2. Femelles………79
ix
3.2 Limite du projet et perspectives futures ... 80
3.2.1. Limites ... 80
3.2.2. Perspectives ... 82
3.3 Conclusion ... 84
ANNEXE 1………85
LISTE DES TABLEAUX
Table 1. Results of generalized linear models for the number of Tree swallow
fledglings per clutch farm-year that recruited into the study system
between 2005 and 2019. 39
Table 2. Results of generalized linear models of natal dispersal distance of
Tree swallow fledglings (males and females) that recruited into the study system between 2005 and 2019.
42
Table 3. Results of linear models assessing the relationship between the
proportion of extensive cultures surrounding breeding and natal sites (within 5 or 20 km) of Tree swallow fledglings that recruited into the
study system between 2005 and 2019 (nID = 190). 45
Table 4. Results of generalized linear (mixed) models assessing the
relationship between the local nest-box occupancy by conspecifics or House sparrows on the breeding and natal sites of Tree swallow fledglings that recruited into the study system between 2005 and 2019
(nID = 190). 46
Table S1. Results of generalized linear model of natal dispersal distance for
males and females recruits of 1-year old tree swallows in southern
Québec. 67
Table S2. Results of generalized linear model of natal dispersal for recruits of
LISTE DES FIGURES
Figure 1. Représentation schématique de la dynamique spatiale d’une
population. Modifiée de Benton et Bowler (2012).
2
Figure 2. Distribution of the 40 farms in southern Québec, Canada, where Tree
swallows were monitored between 2004 and 2019.
32
Figure 3. Relationship between rate of local recruits per clutch farm-year and
A) average fledging mass B) Minimum fledging date C) ENSO index during fall migration for Tree swallows in southern Québec.
40
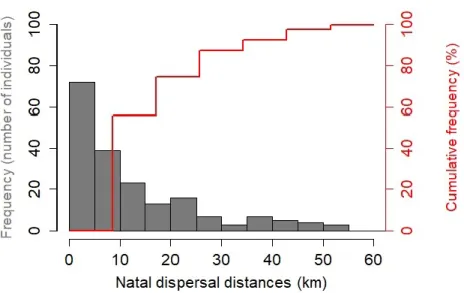
Figure 4. Distribution of natal dispersal distances (km) of Tree swallow
fledglings that recruited into the study system between 2005 and
2019. 41
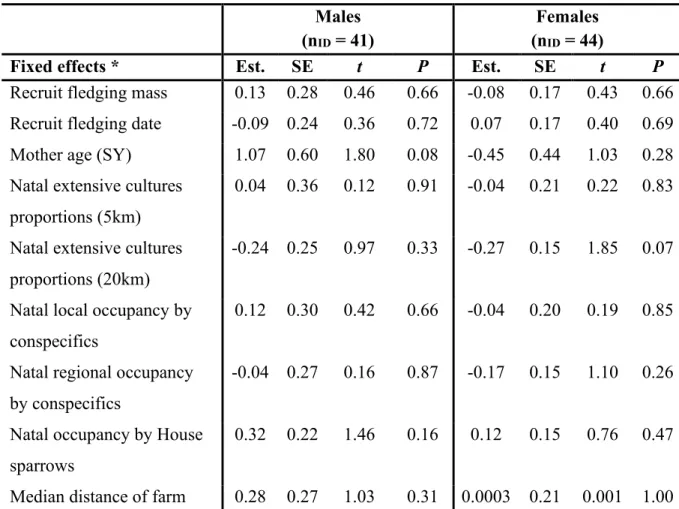
Figure 5. Relationships between natal dispersal distances (km) of Tree swallow
fledglings that recruited into the study system between 2005 and 2019 and A) Natal occupancy by House sparrows of male recruits, B) mother age of male recruits, and C) Natal regional occupancy by conspecifics of female recruits.
43
Figure 6. Relationship between natal and breeding site characteristics of Tree
swallow fledglings that recruited into the study system between 2005 and 2019. A) proportion of extensive cultures within 20 km. B) proportion of extensive cultures within 5 km. C) local nest-box occupancy by conspecifics. D) local nest-box occupancy by House sparrows.
xii
Figure 7. Comparison of the observed coefficients of the relationships linking
breeding to natal habitat characteristics of Tree swallow fledglings that recruited into the study system between 2005 and 2019 (Tables 3 and 4) to coefficients obtained from simulations whereby recruits selected breeding farms at random over the whole system (histogram). A) proportion of extensive cultures within 20 km. B) proportion of extensive cultures within 5 km. C) local nest-box occupancy by conspecifics. D) local nest-box occupancy by House sparrows.
48
Figure S1. Histogram of the observed natal dispersal distances (km) (grey bars)
and the probability curve (black line).
69
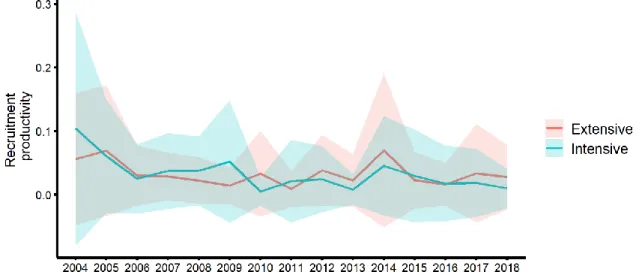
Figure S2. Mean annual recruitment productivity of extensive cultures sites (red)
and intensive cultures sites (blue) across years.
70
Figure S3. Means distribution of simulated dispersal distances (grey bars) and
average observed in the population (red line).
xiii
Figure S4. Comparison of the observed coefficients of the relationships linking
breeding to natal habitat characteristics of Tree swallow fledglings that recruited into the study system between 2005 and 2019 (Tables 3 and 4) to coefficients obtained from simulations whereby recruits selected breeding farms from probability distances (Fig. S3). A) proportion of extensive cultures within 20 km. B) proportion of extensive cultures within 5 km. C) local nest-box occupancy by conspecifics. D) local nest-box occupancy by House sparrows.
72
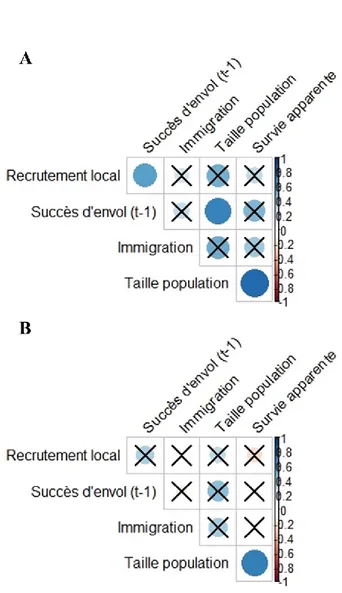
Figure S5. Structure des A) corrélations et B) corrélations partielles de Spearman
des taux vitaux annuels pour la population à l’étude d’Hirondelle bicolore.
LISTE DES ABRÉVIATIONS ET DES SIGLES
ASY After-second-year
ENSO El Niño-Southern Oscillation
rs Coefficient de corrélation de Spearman
rs-p Coefficient de corrélation partiel de Spearman
CHAPITRE 1 INTRODUCTION
1.1. Dynamique des populations et recrutement local
Les études sur la dynamique de population évaluent les mécanismes potentiels pouvant faire varier la taille d’une population dans le temps et l’espace. Selon Turchin (2003), une population correspond à un groupe d’individus d’une même espèce vivant à l’intérieur d’une aire géographique suffisamment grande pour permettre la reproduction ainsi que des comportements de dispersion et de migration. Les modèles théoriques de dynamique de population comprennent quatre facteurs principaux : la naissance (N) et l’immigration (I) de nouveaux individus font augmenter la taille d’une population, tandis que la mortalité (M) et l’émigration (E) d’individus diminuent cette taille (Molles et al., 2017). Les calculs des taux per capita permettent ensuite de mieux comprendre comment et pourquoi la taille d’une population change temporellement. Ces taux sont calculés en divisant le nombre d’individus de N, I, M ou E par le nombre total
d’individus dans la population au temps t (Nt). La taille de la population peut ainsi être estimée
au moment t + 1 à l’aide de l’équation suivante :
𝑁𝑡+1 = 𝑁𝑡+ 𝑁𝑡𝑛 + 𝑁𝑡𝑖 − 𝑁𝑡𝑚 − 𝑁𝑡𝑒 (Équation 1.1)
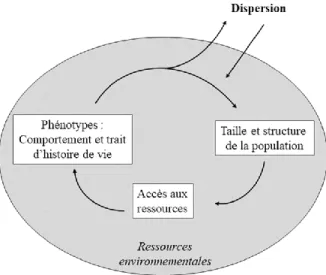
où n, i, m et e sont respectivement les taux per capita du nombre de naissance, d’immigration, de mortalité et d’émigration (Molles et al., 2017). Pour mieux comprendre les mécanismes qui régissent ces taux, il est important d’évaluer l’ensemble des composantes de la dynamique spatiale d’une population. Cette dynamique associe la taille de la population au temps t aux ressources présentes dans l’environnement. Cette interaction détermine la disponibilité des ressources pour les individus pouvant engendrer un débalancement dans leur accès soit par une augmentation de la densité ou par une diminution des ressources. De ce débalancement émergeront différents phénotypes contexte- et condition-dépendant qui développeront un comportement et des caractéristiques de dispersion respectivement à ce contexte (Figure 1, de
2 Benton et Bowler 2012). La dispersion correspond au mouvement exécuté par un individu entre des parcelles d’habitats dans un objectif de reproduction (Clobert et al., 2009).
Figure 1 Représentation schématique de la dynamique spatiale d’une population. Modifiée de Benton et Bowler (2012). La zone grise représente une population.
Les comportements de dispersion des individus entre deux événements de reproduction peuvent autant se faire vers d’autres populations qu’à l’intérieur de la même population (Benton et Bowler, 2012). Cette dispersion résulte du recrutement des individus dans les populations. Ce recrutement peut se faire par immigration lorsque les individus dispersent vers une autre population. Il peut aussi être local si un individu juvénile survit et fait sa première reproduction à l’intérieur de sa population d’origine (Lande et al., 2003). Chez plusieurs espèces, le recrutement local est désigné comme un facteur important dans la dynamique de population (Gaillard et al., 1998; Muths et al., 2011; Wehausen et al., 1987). Or, les mouvements de dispersion sont très variables entre les espèces et résultent en un grand continuum. Alors que certaines espèces ne bougent qu’une seule fois, d’autres bougent continuellement, tandis que d’autres vont en plus avoir des déplacements migratoires. Conséquemment, cette variation de
3 mouvement amène une variation dans le recrutement local rendant nécessaires les études de dynamique spatiale spécifiques au taxon d’intérêt (Benton et Bowler, 2012).
Chez les espèces aviaires, la différence de recrutement local est très marquée entre les espèces résidentes et migratrices. En effet, le pourcentage de retour des juvéniles dans leur population de naissance pour les espèces résidentes peut aller jusqu’à 40%, tandis que chez les espèces migratrices, il se situe habituellement entre 0 et 14% (Weatherhead et Forbes, 1994). Ce faible recrutement local est principalement une conséquence des comportements de dispersion plus élevée chez les espèces migratrices (Newton, 2011) et une plus grande pression sur la survie des juvéniles qui doivent faire face à une plus grande variabilité environnementale durant leurs déplacements entre les aires de reproduction, de migration et d’hivernation (Studds et al., 2008; Weegman et al., 2017). Il est aussi possible d’observer une variabilité du recrutement à l’intérieur même d’une espèce selon le contexte environnemental (Paquet et al., 2018; Winkler et al., 2005) et/ou social (Daniel et Pradel, 2000; Hafner et al., 1998) dans lequel les populations évoluent. Ainsi, étant donné la complexité de ces processus et leurs impacts démographiques, il est important de bien comprendre les déterminants du recrutement local et de la dispersion dans les populations aviaires en milieu naturel.
1.1.1. Survie des juvéniles
Pour des espèces à courte durée de vie, les taux de survie pré-reproduction sont relativement faibles (Low et Pärt, 2009). Par exemple, Naef-Daenzer et al. (2001) rapportent que les taux de survie de la Mésange charbonnière (Parus major) et de la Mésange noire (Periparus ater), deux espèces de passereaux, chutent drastiquement à 50% seulement 10 jours après l’envol. Différents facteurs peuvent être en cause pour expliquer cette mortalité, tels que les caractéristiques individuelles et la pression de prédation (Naef-Daenzer et al., 2001). La masse à l’envol des juvéniles a notamment été identifiée comme une caractéristique importante de la survie (Greño et al., 2008; Monrós et al., 2002; Shutler et al., 2006). En effet, un juvénile avec une masse à l’envol plus grande aura plus de chance de survie et donc une plus grande
4 probabilité de retour (Cox et al., 2018; Hafner et al., 1998). La masse est entre autres influencée par la qualité de l’habitat et l’effort d’alimentation des parents durant le développement des oisillons (Monrós et al., 2002). Ces derniers facteurs sont souvent reliés, car plus la saison de reproduction avance, moins les ressources sont abondantes ou de bonne qualité et donc plus l’effort de recherche alimentaire doit être important (Tarof et al., 2011; Verboven et Visser, 1998). De ce fait, pondre tôt dans la saison de reproduction devient avantageux pour les femelles et favorise la production de jeunes ayant une masse plus élevée (Berzins et al., 2020; Garant et al., 2007a; Lombardo et al., 2020; Müller et al., 2005). La taille de ponte est aussi susceptible d’influencer le nombre d’oisillons envolés (Millet et al., 2015) et par conséquent, la probabilité que des oisillons de cette nichée survivent (Shutler et al., 2006). L’aptitude à pondre tôt et à produire une grande nichée semble toutefois dépendre de la masse corporelle de la femelle; une femelle plus massive à une meilleure capacité à pondre des grandes tailles de nichées tôt dans la saison que les femelles plus légères (Müller et al., 2005; Pellerin et al., 2016).
La grande variabilité présente dans les taux vitaux des juvéniles reflète probablement le fait que leur survie est fort sensible aux changements environnementaux (Clutton-Brock, 1988; Gaillard et Yoccoz, 2003), notamment ceux reliés au climat ou aux conditions météorologiques locales (Berzins et al., 2020). En effet, les températures locales durant la saison de reproduction sont susceptibles d’affecter plusieurs traits, tels que le développement et la croissance des oisillons (Ardia et al., 2009). Par exemple, des températures chaudes au printemps favorisent les femelles à pondre tôt dans la saison (Visser et al., 2009) et améliore le développement embryonnaire des œufs (Ardia et al., 2009) et la croissance corporelle des oisillons (Pérez et al., 2008). Les juvéniles sont toutefois sensibles aux températures extrêmes, froides ou chaudes, qui peuvent avoir un effet négatif sur leur croissance via l’utilisation plus élevée des réserves énergétiques (Greño et al., 2008; Mccarty et Winkler, 1999), par une plus faible disponibilité des ressources alimentaires (Mccarty et Winkler, 1999) ou par un faible apport alimentaire (Tapper et al., 2020).
5 Pour les espèces migratrices, des conditions climatiques difficiles le long de la route migratoire, ainsi que sur les sites de repos ou d’hivernation, peuvent causer des taux de mortalité importants (Cox et al., 2020; Newton, 2011; Studds et al., 2008; Weatherhead et Forbes, 1994; Weegman et al., 2017). En effet, ces espèces peuvent faire face à des conditions extrêmes telles que des tempêtes de neige ou de grêle, du froid, des orages, de forts vents et des éclairs (Newton, 2011). Il a d’ailleurs été montré chez plusieurs espèces que les juvéniles souffrent d’une plus grande mortalité que les adultes durant ces longues phases de déplacement (Owen et Black, 1989; Rotics et al., 2016; Strandberg et al., 2010). Plusieurs raisons peuvent expliquer cette différence, telle que de plus faibles capacités de navigation (Thorup et al., 2003), une susceptibilité aux conditions météorologiques (Owen et Black, 1989) et l’utilisation d’une plus grande réserve d’énergie durant le vol (Rotics et al., 2016). Outre les conditions climatiques, différentes menaces présentes sur les sites de reproduction, d’arrêts migratoires et d’hivernation sont susceptibles d’affecter naturellement la survie des juvéniles, dont la prédation (Cox et al., 2020; Naef-Daenzer et al., 2001), l’exposition aux parasites (McKim-Louder et al., 2013) et l’abondance insuffisante ou incertaine de nourriture (Adams et al., 2006).
Les activités humaines induisent aujourd’hui une modification des habitats qui vient s’ajouter aux pressions amenées par les menaces naturelles et ainsi affecter la survie des juvéniles. Par exemple, les oisillons de l’Hirondelle bicolore (Tachycineta bicolor) présentaient des masses plus légères et une charge parasitaire plus élevée, lorsqu’élevés dans des sites pollués par l’industrie des sables bitumineux de Fort McMurray, Alberta, que ceux ayant grandi dans des sites contrôles non pollués (Gentes et al., 2007). Par ailleurs, la perte d’habitat de reproduction est un facteur potentiel qui prend de plus en plus d’ampleur avec la modification des paysages par le biais de l’action humaine. Par exemple, chez des espèces forestières dont la Paruline bleue (Setophaga caerulescens), les habitats avec une densité élevée d’arbustes sont avantageux pour le succès reproducteur et la survie des juvéniles (Holmes et al., 1996), mais ceux-ci se voient souvent diminués par les activités de déforestation (Imbeau et al., 2001). En général, les activités humaines changent la complexité des structures d’habitat pouvant potentiellement créer des habitats de moins bonnes qualités pour les individus exploitant ces territoires (Scott et al., 2006).
6 1.1.2. La dispersion natale
Quoique la littérature présente plusieurs définitions de la dispersion (Bowler et Benton, 2005; Clobert et al., 2009; Delattre et al., 2013; Ronce, 2007), la plupart réfèrent toutefois aux mêmes fondements génétiques (Clobert et al., 2009) et spatiaux (Cote et al., 2017; Ronce, 2007). La dispersion comprend la dispersion natale, le mouvement entre le site de naissance et le premier site de reproduction, et la dispersion de reproduction, le mouvement entre deux sites de reproduction successifs (Greenwood, 1980; Ronce, 2007). Ainsi, les décisions de quitter, de se déplacer et de s’établir sont les trois étapes exécutées par les individus (Bowler et Benton, 2005; Cote et al., 2017). La dispersion assure un flux génique entre les parcelles (Clobert et al., 2009) pouvant être aléatoire ou non (Jacob et al., 2015). Un comportement de dispersion est considéré aléatoire lorsque les individus se déplacent entre les parcelles indépendamment de leur phénotype et du contexte environnemental (Jacob et al., 2015). Une dispersion non aléatoire résulterait en partie plutôt du choix d’habitat apparié (matching habitat choice) où l’adéquation entre le phénotype de l’individu et l’environnement améliore la valeur adaptative de l’individu (Edelaar et al., 2008). Ce processus non aléatoire est susceptible d’être plus important pour les espèces à forte mobilité et qui dispersent sur de grandes distances et il contribuerait à augmenter les niveaux d’adaptation locale et à stabiliser la dynamique d’une population tout en conservant une certaine variabilité génétique (Delattre et al., 2013; Edelaar et al., 2008; Garant et al., 2007b; Ronce, 2007). Les comportements de dispersion peuvent donc évoluer entre les populations comme à l’intérieur d’une population en fonction des effets phénotypiques et environnementaux (Clobert et al., 2009; Johnson et Gaines, 1990; Paradis et al., 1998). Ce projet se concentre plus spécifiquement sur les comportements de dispersion natale.
1.1.2.1. Variabilité de la dispersion natale
La dispersion est un trait plastique chez les individus (Bowler et Benton 2005, Ronce 2007, Benard et McCauley 2008, Delattre et al., 2013, Jacob et al., 2016) qui peut évoluer différemment entre les espèces, mais aussi fluctuer entre les individus d’une même espèce. Les décisions de dispersion se prennent en fonction des différentes informations que les individus
7 récoltent dans leur environnement (Clobert et al., 2009; Delattre et al., 2013). En général, les distances de dispersion natale sont plus importantes pour les espèces d’oiseaux migrateurs comparativement aux espèces résidentes (Paradis et al., 1998). Ceci peut être expliqué en partie par une déviation des déplacements lors du retour des individus dans l’aire de reproduction (Weatherhead et Forbes, 1994). Étant donné que les espèces migratrices font face à des coûts de dispersion importants durant leur migration en subissant les variations environnementales, il est possiblement parfois plus avantageux pour ces dernières de sélectionner le premier site disponible que de retrouver leur site de naissance (Weatherhead et Forbes, 1994). Au contraire, les espèces résidentes subiraient des coûts de recherche et d’installation si elles décidaient de disperser vers un territoire non familier alors qu’elles ont la possibilité de rester sur leur territoire de naissance pour l’ensemble de leur cycle de vie (Weatherhead et Forbes, 1994). La dispersion natale peut aussi varier entre les sexes comme à l’intérieur d’un même sexe. Chez les oiseaux, la dispersion natale est fréquemment plus importante chez les femelles que chez les mâles (Greenwood et Harvey, 1982; Newton, 2011). Cette dichotomie peut être due à une allocation d’énergie différente entre les sexes ou à l’optimisation de la valeur adaptative inclusive des femelles. En effet, les mâles sont généralement les premiers arrivés aux sites de reproduction et vont utiliser beaucoup d’énergie pour défendre le site (Rendell et Robertson, 1990). Il devient plus avantageux pour les mâles de défendre un site avec lequel ils sont déjà familiers, au lieu de disperser vers un site inconnu (Greenwood, 1980). De plus, chez certaines espèces vivant en groupe, les mâles vont parfois hériter du territoire de leur père, forçant les jeunes femelles apparentées à ces mâles à se déplacer ailleurs afin d’éviter la consanguinité (Newton, 2011). Les femelles vont aussi plutôt utiliser leur énergie afin de chercher et sélectionner le site qui présente des caractéristiques favorables au succès reproducteur. Elles auraient alors plutôt tendance à dévier de leur site natal en se déplaçant d’un site à un autre avant de s’établir au plus favorable (Greenwood 1980). La dispersion natale peut aussi varier à l’intérieur d’un sexe pour une même espèce. Par exemple, une étude sur les Gobe-mouches à collier (Ficedula albicollis) a montré que les jeunes mâles philopatriques obtenaient un meilleur succès reproducteur que les jeunes mâles immigrants (c.-à.-d. disperseurs) (Pärt, 1994). Cet avantage reproducteur peut être expliqué par la connaissance des sites de bonne qualité qui sera plus développée pour les mâles
8 philopatriques (Pärt, 1994). Cette variabilité dans la dispersion natale et les distances de dispersion peut être liée à des facteurs écologiques ou des facteurs phénotypiques.
1.1.2.2. Facteurs de dispersion liés à l’écologie
Les facteurs écologiques font varier les coûts liés à la dispersion ayant comme conséquence d’influencer les différentes stratégies de dispersion (Weatherhead et Forbes, 1994). Ces facteurs écologiques peuvent être reliés à l’environnement social et physique de l’individu. D’abord certains individus vont disperser de leur site natal afin d’éviter la consanguinité (Bowler et Beton, 2005). En dispersant de leur site natal, cela favorise la variabilité génétique des populations, ce qui peut optimiser l’aptitude des individus à répondre aux différents habitats rencontrés durant la migration (Weatherhead et Forbes, 1994). La densité de congénères peut également affecter la dispersion des individus. Une faible densité de congénères peut occasionner un effet Allee (Kim et al., 2009; Matthysen, 2012) via lequel les individus qui se retrouvent sur des sites à faibles densités n’ont pas avantage à y rester du fait d’une valeur adaptative attendue réduite, provoquant leur dispersion vers des sites à plus grandes densités. Ceci a été montré chez l’espèce coloniale du Fou à pieds bleus (Sula nebouxii), chez qui la présence de congénères peut être avantageuse par l’augmentation des opportunités de copulation hors-couple, la défense du territoire et la détection de prédateurs (Kim et al., 2009). Par contre, une forte présence de congénère augmente le risque de parasitisme ou encore les interactions compétitives directes ou non sur l’accès aux ressources (Bowler et Benton, 2005; Matthysen, 2012).
Les facteurs écologiques de dispersion liés à l’environnement physique dépendent principalement de la qualité intrinsèque du site dans lequel l’individu a grandi. Cette qualité peut autant dépendre de la présence de parasites (Boulinier et al., 2001; Brown et Brown, 1992), que du niveau de prédation (Marzluff, 1988) et/ou de compétition interspécifique sur un site (Weisser, 2001). Ces deux derniers facteurs peuvent amener un individu à disperser plus ou moins loin de son site de naissance afin de sélectionner un nouveau site moins stressant et qui
9 favorisera la valeur adaptative de l’individu (Weisser, 2001). Par exemple, une étude sur les comportements compétitifs entre deux espèces de merlebleus, le Merlebleu de l’Ouest (Sialia mexicana) et le Merlebleu azuré (Sialia currucoides), montre un effet adaptatif dans les comportements de dispersion des individus. Les comportements plus agressifs du Merlebleu de l’Ouest semblent être un mécanisme important dans le déplacement des individus du Merlebleu azuré et favorisent la colonisation de nouveaux territoires par le Merlebleu de l’Ouest. Ce lien entre la dispersion et les comportements agressifs a permis au Merlebleu de l’Ouest de coloniser des nouvelles aires de reproduction comparativement au Merlebleu azuré (Duckworth et Badyaev, 2007). La quantité de ressources alimentaires présente sur le site de naissance est aussi un facteur capable d’influencer les mouvements de dispersion des jeunes suite à l’envol (Belthoff et Dufty, 1969). Cette relation est en partie montrée chez l’Autour des palombes (Accipiter gentilis atricapillus), où les juvéniles présentent des déplacements plus près de leur site de naissance lorsqu’ils ont eu accès à plus de nourriture (Kennedy et Ward, 2003). Ceci suggère que les juvéniles qui expérimentent un site de naissance avec de bonnes ressources alimentaires ont tendance à y rester ou à revenir près de celui-ci pour se reproduire.
Les actions humaines amènent des changements importants dans la structure et la composition de l’environnement (Wiens, 2001). Ces changements sont susceptibles d’influencer l’évolution des comportements de dispersion des individus. La fragmentation d’un habitat est un processus durant lequel l’habitat original est divisé en parcelles plus petites, isolées les unes des autres par une matrice d’habitats différents de l’habitat original et couramment entouré de bordure (Fahrig, 2003; Wilcove et al., 1986). Ce changement de structure peut avoir comme conséquence d’augmenter l’isolement des parcelles en réduisant la connectivité structurale entre les types d’habitats (Delattre et al., 2013) et de réduire la variabilité environnementale à l’intérieur des parcelles tout en augmentant cette variabilité entre les parcelles (Cote et al., 2017). L’effet bordure peut, quant à lui, augmenter la variabilité environnementale d’une parcelle lorsqu’elle est suffisamment grande pour différée de la variabilité présente dans la matrice (Cote et al., 2017). L’intensification des pratiques agricoles est un de ces phénomènes qui amènent un changement du paysage par la perte d’hétérogénéité spatiale (Benton et al., 2003). À ce jour,
10 70% des prairies mondiales (Tews et al., 2013) sont consacrées à des types de cultures intensives telles que le maïs, le soya et l’orge (Benton et al., 2003). L’intensification des pratiques agricoles crée des paysages homogènes à travers la perte d’habitat non cultivables, la simplification des rotations de culture, l’utilisation accrue de fertilisants et de pesticides et l’utilisation de machinerie lourde (Benton et al., 2003; Stanton et al., 2018). Cette homogénéisation peut affecter la survie des espèces aviaires exploitant les paysages agricoles par la perte de ressources, l’augmentation de la compétition ou de la prédation et l’augmentation d’espèces invasives (Jongman, 2002). Il est donc possible d’observer des événements de dispersion à longue ou à courte distance à l’intérieur d’une même population selon les niveaux de fragmentation, d’homogénéisation et de la qualité des habitats exploités (Cote et al., 2017; Delattre et al., 2013; Mathias et al., 2001; Wiens, 2001). Par exemple, une distribution groupée de parcelles pourrait favoriser la dispersion à courte distance (Mathias et al., 2001), tandis qu’une diminution de la taille des parcelles et de leur qualité ferait augmenter les distances de dispersion (Cote et al., 2017). Selon le niveau de fragmentation et la vitesse à laquelle le paysage se modifie, les coûts reliés à la dispersion peuvent devenir importants (Baguette et al., 2012). La variabilité environnementale des parcelles pourrait rendre la sélection d’habitat plus difficile, résultant à un taux élevé de mortalité durant la dispersion et/ou une sélection d’habitat non-optimale (Cote et al., 2017). Ces changements affecteront directement la dynamique spatiale des populations.
1.1.2.3. Facteurs de dispersion liés au phénotype
Des différences phénotypiques sont parfois présentes entre les individus qui dispersent et ceux qui ne dispersent pas ou peu (Clobert et al., 2009; Cote et al., 2017). Ces différences touchent autant les traits physiologiques, comportementaux et morphologiques que les traits d’histoire de vie. En particulier, certains traits individuels définis comme « traits de facilitation » (ex : grande longueur des ailes, meilleure condition corporelle, réserves d’énergie élevées) sont des composantes importantes à prendre en considération pour évaluer les capacités de dispersion (Clobert et al., 2009; Cote et al., 2017). En général, les disperseurs présenteraient des tailles et
11 des conditions corporelles supérieures à celles d’individus non-disperseurs (Anholt, 1990; Clobert et al., 2009; O’Riain et al., 1996). La taille corporelle a déjà été montrée comme étant une composante morphologique importante dans la décision de disperser chez les juvéniles d’espèces résidentes. Les juvéniles avec une taille corporelle plus faible restent plus longtemps sur les sites de naissance et sont susceptibles de disperser à de plus faibles distances que les juvéniles de plus grande taille (Belthoff et Dufty, 1969; Nilsson, 1989). Cette relation chez les espèces migratrices reste par contre floue et peu évaluée. Indirectement, il est possible que la taille corporelle des juvéniles migrateurs influence négativement leur dispersion natale étant donné l’effet de cette caractéristique sur la survie (Cox et al., 2018; Hafner et al., 1998) et favorise leur probabilité de recruter localement.
Les effets maternels peuvent aussi jouer un rôle important dans le développement de la dispersion des jeunes (Cote et al., 2007). Ces effets peuvent être liés à l’environnement dans lequel la mère se reproduit (Duckworth, 2009) ainsi qu’à son expérience de reproduction (Angelier et al., 2007). La capacité des femelles à modifier le sexe-ratio de leur nichée est notamment souvent rapportée dans la littérature aviaire (Baeta et al., 2012a; Duckworth, 2009; Nager et al., 1999; Whittingham et Dunn, 2000). En particulier, une étude sur le Merlebleu de l’Ouest a montré que lorsque les ressources disponibles étaient limitées, les mères produisaient plus de mâles tôt dans la saison. Ces mâles développaient un comportement beaucoup plus agressif et dispersif comparativement à ceux nés avec des ressources élevés (Duckworth, 2009). De ce fait, l’effet maternel a une influence sur les capacités des individus à s’installer sur de nouveaux territoires. Aussi, la condition physique (Whittingham et Dunn, 2000) et l’expérience des mères (Angelier et Chastel, 2009) peuvent influencer la production de jeunes et le sexe-ratio des nichées. Les femelles en bonne condition physique et plus âgées ont de meilleures aptitudes à gérer les coûts reliés à la reproduction, tels que la quête alimentaire et l’approvisionnement des oisillons (Angelier et Chastel, 2009; Baeta et al., 2012b). Elles ont ainsi la capacité d'investir leur énergie dans la production du sexe généralement le plus couteux à produire, à savoir les mâles (Baeta et al., 2012b). Ceci résultera à la production de mâles moins stressés et avec une bonne condition physique tout en assurant un succès de nichée plus important que les plus jeunes
12 femelles. Ces mâles sont alors susceptibles de disperser à de plus faibles distances et de recruter dans la population de naissance pour se reproduire (Tinbergen et Boerlijst, 1990; Whittingham et Dunn, 2000).
1.1.3. Sélection d’habitat
À la suite de la période de déplacement durant la dispersion, les individus doivent faire un choix concernant l’habitat qui répondra à leurs besoins. Plusieurs contraintes peuvent influencer la capacité d’un individu à choisir un habitat. D’abord, la capacité de déplacement de l’espèce doit permettre à l’individu d’accéder au site. Ensuite, l’espèce doit présenter des capacités cognitives développées afin de prendre en considération le temps et l’espace (Danchin et al., 2005). Pour que la sélection soit efficace, le milieu ne doit pas varier aléatoirement dans le temps et présenter des caractéristiques prévisibles afin d’acquérir un habitat optimal pour assurer la survie de l’individu (Arlt et Pärt, 2007; Danchin et al., 2005; Klopfer et Ganzhorn, 1985). Les individus vont chercher et sélectionner des habitats pour s’alimenter et se reproduire (Stamps 2014, Danchin et al., 2005). Par conséquent, comme les sites de reproduction sont généralement utilisés sur une plus longue période de temps que les sites d’alimentation, leur sélection peut devenir coûteuse (Boulinier et Danchin 1997, Danchin et al., 2005). C’est pourquoi s’informer sur les caractéristiques de l’habitat avant de s’y établir devient une stratégie avantageuse (Boulinier et Danchin, 1997; Doligez et al., 2003; Parejo et al., 2007; Valone et Templeton, 2002). Ces caractéristiques peuvent correspondent à la qualité et la quantité des ressources et le risque d’exposition aux prédateurs et aux parasites. La présence des congénères sur les différents sites potentiels est une autre caractéristique pouvant refléter la qualité du milieu (Danchin et al., 2005). En évaluant l’ensemble des habitats exploités par une espèce, il est possible d’observer leurs effets sur la valeur adaptative des individus les occupant (Holmes et al., 1996), pouvant ainsi influencer leur décision de revenir ou non sur ces sites les années suivantes (Petit et Petit 1996).
13 1.1.3.1. Le comportement de prospection
La prospection joue un rôle important dans la sélection d’habitat (Cadiou et al., 1994). Lorsqu’un individu doit choisir un site de reproduction, il doit avoir le temps d’analyser le plus d’éléments possible. Chez certaines espèces aviaires, des individus vont se déplacer dans leur environnement afin d’évaluer les sites potentiels de bonne qualité pour la prochaine année de reproduction. Ce comportement se qualifie de prospection (Boulinier et Danchin, 1997; Danchin et al., 2001, 2005). Les prospecteurs peuvent autant être des jeunes immatures (pré-reproducteurs), des adultes non reproducteurs que des adultes qui ont échoué leur reproduction (Cadiou et al., 1994; Danchin et al., 2005). Notamment, la prospection chez les juvéniles est un élément pouvant influencer la sélection de leur premier site de reproduction et par conséquent, leur retour dans la population natale (Nocera et al., 2006; Pärt et Doligez, 2003; Pegan et al., 2018).
1.1.3.2. Sélection d’habitat non aléatoire
L’hétérogénéité spatiale des conditions environnementales engendre une structure spatiotemporelle des phénotypes à travers le paysage où la valeur adaptative des individus va différer entre les habitats (Michel 2011). Les individus vont alors sélectionner des habitats dépendant à leur phénotype. Cette sélection non aléatoire résulte des comportements de dispersion non aléatoire décrits plus haut (Jacob et al., 2015). Plusieurs sources peuvent se trouver sous ce mécanisme évolutif qui affectera la sélection d’un habitat. La préférence de l’habitat de naissance est une de ces sources qui a été montrée chez plusieurs groupes taxonomiques (insectes, poissons, amphibiens, oiseaux et mammifères) (Immelmann 1975; Stamps 2001; Davis et Stamps, 2004). Ce phénomène se produit lorsqu’un individu exposé de manière précoce à un stimulus dans son environnement de naissance développera un phénotype dépendant à cet environnement. L’individu acquiert une préférence pour ce type d’habitat, ce qui augmente la probabilité qu’il sélectionne ce même habitat plus tard (Stamps 2001; Danchin et al., 2005; Immelmann 1975; Davis et Stamps, 2004). La préférence à l’habitat de naissance
14 serait impliquée dans l’écologie et l’évolution de la sélection d’habitat de trois manières. D’abord, la formation d’un phénotype dépendant à l’environnement de naissance génère des variations individuelles dans la préférence d’habitat d’une population. Les individus vont alors sélectionner un habitat auquel ils sont bien adaptés, occasionnant par le fait même une variation génétique dans un paysage hétérogène. Finalement, ces traits phénotypiques plastiques sont héritables et transmis à la descendance et seront exprimés en fonction de l’environnement de naissance (Davis et Stamps, 2004).
Un second mécanisme non aléatoire qui résulte aussi de la dispersion non aléatoire est le choix d’habitat apparié. Un individu avec un phénotype précis sélectionnera un site avec un environnement qui correspond le mieux à ses capacités d’utilisation. Par exemple, l’individu se déplace vers un environnement qui correspond à son phénotype lié à des caractéristiques telles que l’âge, la taille et le sexe au lieu de changer son trait dépendamment de l’environnement (Clobert et al., 2012; Edelaar et al., 2008). Ce changement dynamique de choix est favorable au maintien de la diversité au sein d’une population en promouvant les adaptations locales, où les individus avec des traits écologiques semblables se rassemblent (Edelaar et al., 2008). Ces processus de sélection d’habitat non aléatoire qui favorisent la diversité deviennent importants dans la dynamique des populations en contribuant à la productivité, à la survie et au recrutement des individus à l’intérieur des habitats disponibles (Holmes et al., 1996).
1.1.3.3. Sélection densité-dépendante
Le modèle de distribution idéal libre de Fretwell et Lucas (1970) amène deux grandes prédictions : (1) les individus sont libres de sélectionner n’importe quel habitat sans coût ni contrainte de déplacement, et ce de manière égale avec les autres espèces résidentes et (2) ils sélectionneront l’habitat idéal pour optimiser leur survie et reproduction. Il suppose aussi que la valeur adaptative des individus sera dépendante négativement à la densité de congénère étant donné la compétition intra-spécifique qui en découle pour l’accès aux ressources (Fretwell, 1969). Cependant, son application en milieux sauvage reste peu plausible étant donné les nombreuses déviations qu’il comporte. Ceci inclut les différences en habileté compétitive des
15 individus, la disponibilité hétérogène des habitats (Sutherland, 1996), les coûts de déplacement (Bernstein et al., 1991) et les différences individuelles de l’évaluation de la qualité des habitats (Edelaar et al., 2008). En évaluant ces composantes dans les populations sauvages, deux hypothèses opposantes ressortent aujourd’hui quant à l’effet de la densité de congénère sur la sélection d’habitat (Stamps, 1991, 1988). Basée sur le modèle théorique de Fretwell et Lucas (1970), l’hypothèse d’évitement prédit que le taux d’établissement sera plus élevé pour des parcelles à moins grande densité (Stamps, 1991) afin de limiter la compétition intra-spécifique (Danchin et al., 2005). L’hypothèse d’attraction prédit plutôt que le taux d’établissement sera plus élevé à mesure qu’il y aura une augmentation de congénère sur la parcelle (Stamps, 1991). Chez certaines espèces, la forte densité d’une parcelle est avantageuse pour le succès reproducteur, la protection aux prédateurs, la défense d’intrus ou pour la prise d’information sur l’habitat (Stamps, 1988). Ces avantages sont alors indices de qualité pour les individus favorisant l’attraction des autres congénères (Stamps, 2001; Danchin et al., 2005; Muller et al., 1997).
1.1.3.4.Sources d’information lors de la sélection d’habitat
Peu importe le processus de sélection d’habitat que les individus utilisent, ils devront analyser plusieurs sources d’informations pour sélectionner un habitat (Doligez et al., 2002 ; Danchin et al., 2001 ; Danchin et al., 2005). L’utilisation de cette information dépendra de sa valeur, définie comme étant la balance entre les coûts pour son obtention et les bénéfices qu’elle apporte une fois utilisée (Danchin et al., 2001). L’optimisation de la valeur de l’information dépend de l’autocorrélation temporelle des conditions environnementales entre le moment de la prise d’information et la décision qui en découle. De plus, les facteurs environnementaux doivent être intégrés dans l’information afin de révéler la qualité globale de l’habitat. Ces derniers facteurs faciliteront l’évaluation et l’étendue de l’information pouvant être utilisée (Danchin et al., 2001).
16 L’abondance de nourriture et l’absence de prédateur sont des éléments seulement évaluables directement une fois que l'individu est installé dans le milieu (Danchin et al., 2004). Or, évaluer l’information directement peut devenir coûteux si ces critères ne se sont pas manifestés durant l’établissement, mais plus tard durant la saison. Pour éviter ce coût, l’information peut être évaluée de manière indirecte sous l’utilisation d’indices sociaux qui révèlent la présence des ressources et des prédateurs (Danchin et al., 2004). Cette source d’information sera principalement utilisée lors du processus de sélection d’habitat densité-dépendante. L’information sociale permet aux prospecteurs de suivre les interactions des congénères et leur comportement à l’intérieur d’un environnement candidat (Danchin et al., 2004; Valone et Templeton, 2002). Cette information peut provenir d’indices involontaires de la performance des congénères dans l’habitat, où les individus vont s’établir à une parcelle en fonction de la densité présente l’année précédente au sein de cette même parcelle (Doligez et al., 2003; Nocera et al., 2006). La présence des congénères avec des exigences similaires à l’individu dévoile un bon indice pour indiquer la localisation des ressources (Danchin et al., 2004). À ce titre, il a été montré que les juvéniles du Goglu des prés (Dolichonyx oryzivorus) utilisent la présence de congénères comme valeur de l’habitat à la fin de leur année de naissance pour déterminer l’habitat d’établissement l’année suivante (Nocera et al., 2006; Parejo et al., 2007).
L’analyse du succès des congénères peut aussi fournir une source d’information indirecte dite publique (Valone et Templeton, 2002). Ce type d’information permet de fournir des indices sur la qualité des ressources environnementales via le succès des congénères dans l’habitat cible (Danchin et al., 2004; Valone et Templeton, 2002). C’est une information désignée comme plus fiable que l’information véhiculée par la densité de congénère (Doligez et al., 2003) et par l’expérience personnelle puisqu’elle présente une estimation plus inclusive des paramètres environnementaux de l’habitat (Danchin et al., 2004). L’information peut provenir de l’activité d’alimentation, du succès reproducteur, des interactions des congénères et du choix de partenaires (Danchin et al., 2004). Les individus vont alors copier la sélection d’habitat des autres pour sélectionner leur propre site (Danchin et al., 2004; Parejo et al., 2007). Ce comportement est souvent utilisé chez les espèces migratrices étant donné la limite de temps
17 qu’elles ont pour évaluer la qualité d’un habitat (Parejo et al., 2007). Par exemple, l’étude de Brown et al. (2000) chez l’Hirondelle à front blanc (Petrochelidon pyrrhonota) montre l’utilisation du succès reproducteur local, évalué par le nombre d’oisillons envolés, comme information pour copier les sites de reproduction lors de l’établissement la saison suivante. Ainsi, un site présentant une augmentation du nombre d’envolé engendre une augmentation d’immigration et une diminution de l’émigration l’année suivante (Doligez et al., 2002).
Certaines espèces vont aussi utiliser leur expérience personnelle pour prendre une décision sur l’habitat à sélectionner (Doligez et al., 2004). Des études sur les adultes reproducteurs de l’Hirondelle bicolore (Lagrange et al., 2017) et l’Hirondelle à front blanc (Brown et al., 2000) présentent des résultats concordant avec cette prise d’information. En effet, ces espèces semblent utiliser leurs propre succès reproducteur et condition physique pour prendre les décisions de rester ou de quitter. Cependant, cette stratégie ne peut pas être utilisée par les juvéniles et les non reproducteurs puisqu’ils n’ont pas l’expérience requise pour prendre une telle décision (Doligez et al., 2003). En somme, ces dernières sources d’informations ne sont pas exclusives et peuvent être utilisées de manière plus ou moins importante selon la variation spatio-temporelle de l’environnement (Doligez et al., 2003). C’est pourquoi il est important d’inclure une variété de sources d’information lors des analyses de sélection d’habitat afin d’avoir une meilleure connaissance des déterminants des choix réalisés.
1.1.3.5. La sélection d’habitat à multiples échelles
Depuis quelques décennies, il est reconnu que la sélection d’un habitat par un individu est déterminée de manière hiérarchique en dépendant de plusieurs échelles spatiales ( Hildén, 1965; Wiens, 1989; Orians et Wittenberber, 1991). Il a été conceptualisé qu’une première sélection d’habitat est faite à l’échelle de l’aire de répartition géographique, suivie d’une sélection au niveau du domaine vital, puis d’une au niveau d’un site spécifique, et finalement d’une sélection des ressources à utiliser (Hutto, 1985; Johnson, 1980). Ainsi, à plus grande échelle, la structure du paysage reliée aux conditions climatiques et à la végétation déterminera la distribution d’une
18 espèce. Tandis qu’à plus fine échelle, se sont plutôt des processus biologiques reliés principalement à des sources d’informations sociales ou publiques qui influenceront la distribution des individus dans l’espace (Bailey et Thompson, 2007; Wiens, 1989). L’analyse de la sélection d’habitat a plusieurs échelles spatial devient primordiale afin d’évaluer les différents facteurs impliqués dans le processus.
1.2. Objectif et importance du projet
Ce projet vise à évaluer les déterminants qui influencent le recrutement local dans une population d’Hirondelle bicolore et ceux qui influencent les comportements de dispersion natale des individus qui recrutent localement dans la population. Je m’attarderai aussi au processus de sélection d’habitat de reproduction des recrues, notamment en évaluant si les habitats de reproduction choisis diffèrent des habitats de naissance. Mon projet permettra d’avoir une meilleure connaissance des processus qui influencent la dynamique de population chez l’Hirondelle bicolore. L’étude du recrutement locale est bien détaillée chez les espèces de passereaux résidents, mais elle demeure très préliminaire et ardue chez les espèces migratrices. Dans mon étude, je fournirai donc de nouvelles connaissances en ce qui concerne le recrutement local sur une longue période de temps (16 années) tout en prenant en considération les effets possibles d’environnements de qualités variables et contrastées, ce qui n’a pas été effectué dans les études précédentes (Cox et al., 2018; Shutler et al., 2006; Taylor et al., 2018; Winkler et al., 2004). Finalement, les résultats de ce projet amèneront une meilleure connaissance de ce qui influence le recrutement et les comportements de dispersion dans une population en déclin.
1.3. Méthodologie générale
1.3.1. Espèce à l’étude
L’Hirondelle bicolore est un passereau (~20 grammes) insectivore et migrateur néotropical de la famille des Hirundinidae nichant principalement en milieux agricoles et humides à travers
19 l’Amérique du Nord (Winkler et al., 2011). Cette espèce est largement étudiée vu sa facilité de manipulation et sa capacité à nicher dans des cavités secondaires, telles que des nichoirs artificiels (Jones, 2003). Dans le Sud du Québec, les individus s’établissent dans leurs nichoirs à la mi-avril, les premiers œufs sont pondus début mai et les premiers oisillons naissent en début juin. La période d’envol se déroule durant le mois de juillet. Les juvéniles et les adultes se regroupent ensuite avant d’entamer leur migration vers le sud au mois d’août. Les populations du Québec hiverneront plus précisément en Floride et à Cuba (Knight et al., 2018).
L’Hirondelle bicolore présente un domaine vital de superficie variable selon les différentes étapes de son cycle de reproduction. Avant la période de ponte, les adultes peuvent couvrir une
superficie de 175 km2 de leur site de reproduction pour s’alimenter et rechercher des partenaires
(Lessard et al., 2014). Cette superficie diminue probablement à 78 km2 après l’incubation pour
l’alimentation des jeunes (Ghilain et Belisle, 2008). Les mâles montrent un comportement plus territorial que les femelles en défendant le nichoir sélectionné (Rendell et Robertson, 1990). Cette espèce est socialement monogame, mais présente un système de reproduction complexe de promiscuité génétique (Porlier et al. 2009; Whittingham et Dunn 2010; Lessard et al., 2014). En effet, 50% à 90% des nids contiennent au moins un oisillon provenant d’un mâle hors couple (Dunn et Whittingham, 2005; Dunn et al., 1994; Lessard et al., 2014; Whittingham et Dunn, 2001).
L’Hirondelle bicolore présente actuellement des déclins de population dans le Nord-Est de l’Amérique du Nord (Michel et al., 2016; Nebel et al., 2010; Shutler et al., 2012). Le recrutement local devient donc un facteur important à étudier afin de mieux comprendre son impact sur la dynamique de population des Hirondelle bicolore. Les études précédentes sur cette espèce rapportent des taux de recrutement variables. Alors que certaines études obtiennent des taux presque nuls (Cox et al., 2018; Taylor et al., 2018; Whittingham et Dunn, 2001), d’autres détectent des taux supérieurs à 5% (Berzins et al., 2020; Lombardo et al., 2020; Shutler et Clark, 2003; Winkler et al., 2005). Par contre, la connaissance des déterminants de ces taux de recrutement reste méconnue. Aussi, comme cette espèce est migratrice, les conditions
20 climatiques rencontrées le long des corridors migratoires et aux divers sites d’hivernation amènent de nombreux défis de survie (Cox et al., 2020; Knight et al., 2019; Studds et al., 2008; Weegman et al., 2017). La dispersion natale chez cette espèce reste aussi peu étudiée. Par exemple, Winkler et al. (2005) présentent des distances de dispersion natale moyennes de 10 km, mais ne rapportent aucun déterminant pouvant expliquer la variation présente dans ces distances. L’étude des comportements de prospection chez les juvéniles montre toutefois qu’ils auraient la capacité de prospecter jusqu’à 20 km de leur site de naissance pour acquérir de l’information (Pegan et al., 2018; Winkler et al., 2005). Cette prise d’information est susceptible d’influencer la dispersion natale et le recrutement local.
1.3.2. Aire d’étude
Les données utilisées dans ce mémoire proviennent du suivi d’un système d’étude situé dans le Sud du Québec, Canada, dans les régions administratives de l’Estrie et de la Montérégie et
couvrant une superficie de 10 200 km2 (Ghilain et Bélisle, 2008). Depuis 2004, un total de 40
fermes est suivi à l’intérieur d’un gradient d’intensification agricole d’est en ouest. Chacune de ces fermes contient un transect composé de 10 nichoirs espacés de 50 m et dont l’ouverture est orientée sud-est. Chaque ferme est visitée aux deux jours afin de faire le suivi des nids, des œufs, des adultes, des oisillons et des conditions environnementales. Dans le cadre de mon projet, les données obtenues de 2004 à 2019 ont été utilisées.
1.4. Hypothèses et prédictions
En me basant sur la littérature résumée plus haut, je m’attends à ce que le recrutement local annuel varie en fonction de l’environnement duquel les juvéniles sont issus l’année précédente, lequel influence la date d’envol, le nombre de juvéniles envolés et leur masse à l’envol. Ainsi, j’anticipe qu’il y aura un plus grand retour des juvéniles lorsque les dates d’envol sont tôt dans la saison, le succès d’envol est élevé sur les sites de naissance, et que les masses à l’envol sont élevées. La compétition interspécifique pour les sites de nidification, la densité de congénères
21 et le type de paysage agricole autour des fermes devraient aussi affecter le recrutement. Je m’attends à ce que les sites de naissance présentant un niveau de compétition interspécifique faible, une densité élevée de congénères et un paysage agricole extensif favorisent le retour des juvéniles dans le système l’année suivante. Je m’attends à ce que les conditions climatiques rencontrées au cours de la première migration d’hivernation et de reproduction aient un fort impact sur la survie des juvéniles durant leur première année. Ainsi, des conditions climatiques favorables, soient des températures clémentes et peu de précipitations, devraient favoriser la survie des juvéniles et par le fait même, leur retour dans leur population de naissance.
Je m’attends également à ce que les recrues locales dispersent de manière non-aléatoire dans le système d’étude. Les distances de dispersion natale des recrues seront influencées par les caractéristiques de leur site de naissance. Je prédis que les recrues nées dans de bons habitats (c’est-à-dire un habitat avec un meilleur potentiel pour le succès reproducteur) devraient disperser sur de plus petites distances ou retourner à leurs sites de naissances. Ces derniers correspondent à des habitats présentant une faible compétition interspécifique pour les sites de nidification, une densité élevée de congénères et une proportion de cultures extensives élevée. Je m’attends aussi à ce que les individus qui se sont envolés tôt dans la saison et qui proviennent de mères plus expérimentées dispersent sur de plus faibles distances dans le système.
Finalement, je m’attends à ce que les recrues qui dispersent de leurs sites de naissance se déplacent vers des sites présentant des caractéristiques plus favorables à la reproduction que leurs sites de naissance. Je prédis donc que les individus nés sur des sites avec une faible proportion de cultures extensives, une faible densité de congénères et une forte densité de compétiteurs interspécifiques auront tendance à disperser vers des sites aux caractéristiques opposées.
CHAPITRE 2
DÉTERMINANTS DU RECRUTEMENT LOCAL ET DE LA DISPERSION NATALE CHEZ UNE POPULATION DE PASSEREAUX EN DÉCLIN
2.1 Introduction à l’article
Cet article porte sur l’évaluation des déterminants du recrutement local et de la dispersion natale chez une population d’Hirondelle bicolore nichant le long d’un gradient d’intensification agricole dans le Sud du Québec. Les déterminants du recrutement local chez cette espèce demeurent méconnus étant donné la difficulté à recapturer les juvéniles au stade adulte dans les populations nicheuses. À l’aide d’une étude longitudinale en cours à l’Université de Sherbrooke depuis 2004, il a été possible de recapturer suffisamment d’individus locaux afin d’évaluer les déterminants qui influencent leur retour dans leur population d’origine, puis d’évaluer les caractéristiques qui influencent leur dispersion natale et le processus de sélection de leur premier site de reproduction. Les résultats présentés permettront de mettre en lumière les mécanismes influençant les comportements de mouvement des juvéniles et la sélection de leur premier habitat de reproduction. Ceux-ci constituent une contribution importante pour l’étude du recrutement local qui est un taux vital important dans la régulation de plusieurs populations aviaires sauvages.
Cet article a été écrit en collaboration avec Dany Garant, Marc Bélisle et Fanie Pelletier. J’y ai contribué significativement par la récolte des données sur le terrain, le traitement des échantillons en laboratoire, les analyses statistiques et la rédaction de la première version complète du manuscrit. Mon directeur, Dany Garant, a supervisé le projet et, avec mon codirecteur, Marc Bélisle, participé à l’élaboration des questions de recherche, à l’interprétation des résultats, à la révision du manuscrit et au financement du projet. Fanie Pelletier a également participé à l’amélioration de ce manuscrit par ses commentaires et a aussi participé à l’encadrement et au financement du projet. Cet article constitue le cœur de ce mémoire et a été soumis à la revue scientifique Oikos à l’hiver 2021.
24 Determinants of local recruitment and natal dispersal in a declining bird population
by
25 2.2 Abstract
Recruitment and juvenile dispersal are important demographic rates and studying their determinants is particularly important in the current context of global anthropogenic perturbations. In birds, and especially for migratory species, assessing these rates is challenging because of the difficulties involved in tracking individuals between their fledging and return to a breeding population. Here we assessed the determinants of local recruitment and natal dispersal distances in Tree swallows (Tachycineta bicolor), a aerial insectivore. To do so, we used a 16-year dataset obtained from the long term monitoring of a population breeding within
a 10 200 km2 study system located along a gradient of agricultural intensification in southern
Québec, Canada. Our results showed that yearly local recruitment rates ranged between 1.0% and 3.2%. Earlier fledging date and heavier fledglings were negatively and positively related to greater recruitment from a given clutch respectively. Our results also showed that natal dispersal distances were generally short (mean = 12.7 ± 13.8 km (SD)) and were influenced by different factors depending on sex. Females dispersed less when conspecific occupancy on breeding site was high, while males dispersed more in the presence of competing House sparrows (Passer domesticus) and when their mother was young. Selection of breeding locations appeared to take place at multiple scales and individuals recruited in sites with characteristics similar to their natal sites. Our results allow to qualify for the first time the information of local recruitment and natal dispersal dynamics in this migratory species. These factors will now be important to consider for the management of this species in order to promote the return and maintenance of individuals in their natal population.
Keyword: farmland birds, local recruitment, migratory species, natal dispersal, natal habitat, population dynamics, tree swallow