CLAUDIA CÁCERES CUADROS
MICRO-IMAGERIE PAR RÉSONANCE
MAGNÉTIQUE NUCLÉAIRE DE LA
DISTRIBUTION DE L’EAU LIQUIDE DANS LE
BOIS D’ÉRABLE À SUCRE AU-DESSOUS DU
POINT DE SATURATION DES FIBRES
Mémoire présenté
à la Faculté des études supérieures de l’Université Laval dans le cadre du programme de maîtrise en sciences du bois
pour l’obtention du grade de maître en sciences (M.Sc.)
DÉPARTEMENT DES SCIENCES DU BOIS ET DE LA FORÊT FACULTÉ DE FORESTERIE, DE GÉOGRAPHIE ET DE GÉOMATIQUE
UNIVERSITÉ LAVAL QUÉBEC
2009
Résumé
La présente étude a pour but d’évaluer le comportement à la sorption du bois d’érable à sucre (Acer saccharum Marsh.) à des teneurs en humidité au-dessous du point de saturation des fibres (PSF). Ces teneurs en humidité d’équilibre atteintes en désorption aux humidités relatives au-dessus de 60% divergent en fonction de l’état initial de saturation. Ce phénomène est nommé "hystérèse à la saturation d’humidité". Plusieurs auteurs ont attribué cette différence à la présence d’eau liquide lors de la désorption. On a alors voulu déterminer la localisation de l’eau liquide dans la structure du bois. Des essais de désorption ont été réalisés entre 58% et 96% d’humidité relative à 21°C à partir de deux états d’humidité, soit le PSF ou la saturation intégrale. On a ainsi pu comparer des échantillons équilibrés à un même niveau d’humidité relative mais avec ou sans présence d’eau liquide. L’imagerie par résonance magnétique, technique basée sur les temps de relaxation des protons, a servi à visualiser la présence d’hydrogène (sous la forme de H20) dans les échantillons. Cette méthode est sensible aux niveaux d'hydratation des tissus ligneux. Les temps de relaxation nous aident à distinguer les types d’eau présents dans le bois étant donné qu’ils sont liés à la grandeur et à la proportion des différents tissus ligneux. L’intensité du signal (représentée par la variation de couleur dans les images de résonance magnétique) a permis de discerner la distribution d’eau liquide dans les tissus du bois. Tel que prévu, l’eau liquide a été principalement observée dans les échantillons provenant d’une saturation intégrale du bois. Finalement, les observations montrent que l’eau restant dans le bois au-dessous du PSF serait localisée dans les bouts de lumina des fibres ligneuses simpliciponctuées, étant donné qu’elles semblent être les tissus les plus isolés du bois d’érable à sucre. La possibilité que cette eau puisse rester enfermée dans les espaces intercellulaires associés aux fibres ligneuses simpliciponctuées reste encore à vérifier.
Avant-Propos
Ce document est présenté sous la forme d’un mémoire de publication. Il a été conçu selon les critères de présentation adoptés par le comité des programmes de 2ième et 3ième cycles en sciences du bois de l’Université Laval, en juillet 1998.
On retrouve dans cet ouvrage une revue de littérature au chapitre 1, laquelle a été divisée en quatre parties couvrant les aspects principaux du présent sujet de recherche. Les propriétés physiques du bois reliées à l’hygroscopicité, les bases théoriques de l’imagerie par résonance magnétique (IRM), l’application de la technique de résonance magnétique nucléaire (RMN) dans le domaine du bois et finalement l’utilisation de l’IRM aussi dans le domaine du bois y sont présentées.
Le chapitre 2 décrit le matériel utilisé et explique en détail la méthodologie suivie lors de cette étude.
Enfin, un article scientifique rédigé en anglais est présenté dans le chapitre 3 :
Magnetic resonance micro-imaging of liquid water distribution in sugar maple wood below fiber saturation point. Cet article sera soumis sous peu pour fins de
publication dans la revue scientifique Wood and Fiber Science.
Les résultats du présent travail ont été également présentés lors du congrès : II Latin American Workshop on Non-destructive Testing of Wood Products, qui a eu lieu à San José, Costa Rica, du 2 au 4 Décembre 2008.
Je voudrais remercier sincèrement au professeur Roger Hernández, directeur de ce projet de maîtrise, qui m’a guidé avec patience au cours de chacune de ces étapes et a été toujours présent pendant sa réalisation. Merci aussi à tous les personnes qui ont contribué à ce projet, spécialement aux techniciens du Centre de Recherche sur le Bois.
Finalement, je tiens à remercier Damien pour son support inconditionnel et à tous les amis qui ont été avec moi d’une façon ou l’autre tout au long de mes deux années d’études.
A Dios, ante todo. A mi papá, por siempre alentarme a ser mejor. A mi mamá, por su amor incondicional. Y a mis hermanos, por estar siempre conmigo.
Table des matières
Résumé... ii
Avant-Propos ... iii
Table des matières... vi
Liste des tableaux... viii
Liste de Figures... ix Introduction... 1 Chapitre 1... 3 1. Revue de littérature ... 3 1.1 Physique du bois ... 3 1.1.1 Domaine hygroscopique ... 3
1.1.2 Hystérèse de la sorption d’humidité... 4
1.1.3 Point de saturation des fibres (PSF)... 8
1.1.4 Hystérèse à saturation d’humidité... 10
1.1.5 Effet de l’hystérèse à saturation sur les propriétés du bois ... 12
1.2 Bases théoriques de l’imagerie par résonance magnétique... 16
1.2.1 Le magnétisme nucléaire... 16
1.2.2 Mouvement de précession et fréquence de Larmor ... 17
1.2.3 Résonance magnétique nucléaire (RMN) ... 19
1.2.4 Phase d’excitation et de relaxation... 19
1.2.5 Temps T1 et T2 ... 22
1.2.6 Signal en IRM ... 23
1.2.7 Contraste en T1, T2 et densité protonique ... 27
1.2.8 Formation de l’image ... 30
1.2.9 Facteurs de qualité de l’image en IRM ... 34
1.2.10 Les paramètres en exploration IRM... 36
1.3 La résonance magnétique nucléaire dans le domaine du bois... 38
1.4 Imagerie par résonance magnétique dans le domaine du bois ... 49
1.5 Hypothèses et objectifs du présent travail... 57
Chapitre 2... 58
2. Matériel et méthodes expérimentales... 58
2.1 Espèce ligneuse ... 58
2.2 Préparation et sélection des éprouvettes ... 58
2.3 Traitements préalables des éprouvettes... 61
2.4 Essais de sorption... 62
2.4.1 Caractéristiques générales... 62
2.4.2 Méthode des solutions salines saturées... 62
2.5 Imagerie par résonance magnétique... 65
2.6 Images par microscope électronique à balayage (MEB et MEBE)... 66
Chapitre 3... 67
3. Article scientifique... 67
3.1 Abstract ... 67
3.2 Introduction and background ... 67
3.3 Materials and methods ... 70
3.3.1 Sorption tests... 70
3.3.3 Environmental scanning electron microscopy (ESEM) and
scanning electron microscopy (SEM) tests... 72
3.4 Results and discussion ... 73
3.4.1 Wood hygroscopicity ... 73
3.4.2 MR micro-imaging analysis... 76
3.4.3 MR micro-images - SEM parallel... 80
3.5 Summary and Conclusions... 85
3.6 Acknowledgements ... 86
Conclusions et Recommandations ... 87
Liste des tableaux
Tableau 1.1
...47 Teneur en humidité d’équilibre (Héq) et temps de relaxation T2 en fonction de l’humidité relative (HR) à 25°C du bois d’érable à sucre.
Tableau 1.2
...48 Teneur en humidité d’équilibre (H ) et temps de relaxation T2 en fonction de l’humidité relative (HR) à 25°C du bois de hêtre à grandes feuilles.
éq
Tableau 1.3
...48 Teneur en humidité d’équilibre (H ) et temps de relaxation T2 en fonction de l’humidité relative (HR) à 25°C du bois de huayruro .éq
Tableau 2.1 Caractéristiques physico-mécaniques du bois d’érable à sucre ...59
Tableau 2.2 Caractéristiques anatomiques du bois d’érable à sucre...59
Tableau 2.3 Caractéristiques des conditions de sorption utilisées...60
Table 3.1 Characteristics of the moisture sorption conditions used in this experiment...72
Table 3.2
...76 Equilibrium moisture content (EMC) as a function of relative humidity (RH) at 21°C for different sample orientations of sugar maple wood.
Liste de Figures
Figure 1.1 Isothermes de sorption d’humidité du bois d’érable à sucre à 21°C... 5
Figure 1.2
... 7 Isothermes de sorption d’humidité du bois d’érable à sucre aux températures de 5° 21º 35° et 50°C.
Figure 1.3
... 12 Équilibre hygroscopique moyen de l’aubier de pruche de l’ouest à 21°C, obtenu avec la méthode des plaques et membranes poreuses sous pression, aux humidités très élevées.
Figure 1.4
... 14 Retraits radial et tangentiel du bois de hêtre en fonction de son humidité d'équilibre en adsorption et en désorption à environ 25°C.
Figure 1.5
... 14 Coefficients de retrait du bois d’aubier de l’érable à sucre en fonction de sa teneur en humidité d'équilibre. Le symbole ■ représente la valeur estimée du PSF.
Figure 1.6
... 15 Coefficients de retrait du bois de tornillo en fonction de sa teneur en humidité d’équilibre.
Figure 1.7 La structure de l’atome... 18
Figure 1.8 Le spin et moment magnétique du proton... 18
Figure 1.9 Les protons s’orientent selon la direction d’un champ magnétique extérieur (Bo) avec l’apparition d’un vecteur d’aimantation
macroscopique M (Σ = zoM ) ... 18.
Figure 1.10 Mouvement de précession... 19
Figure 1.11
... 21 L’onde RF provoque des transitions du niveau de basse énergie E1 au niveau de haute énergie E2.
Figure 1.12 Effet des impulsions RF sur le vecteur d’aimantation macroscopique M ... 21
Figure 1.13 Interaction entre l’onde de radiofréquence et les protons placés dans le champBo... 22
Figure 1.14
... 24 Courbe exponentielle de repousse de l’aimantation longitudinale en fonction du T1.
Figure 1.15
... 24 Courbe exponentielle de disparition de l’aimantation transversale en fonction du T2.
Figure 1.16 Signal de précession libre... 25
Figure 1.17 Notion de T2*... 26
Figure 1.18
... 29 Chronologie des événements au cours du phénomène de l’écho de spin.
Figure 1.19
... 29 Chronologie des impulsions RF et paramètres de la séquence d’écho spin
Figure 1.20 Champ de vue, matrice et plan de coupe. ... Erreur ! Signet non défini.
Figure 1.21 A) Le gradient de sélection de coupe Gss, B) Le gradient de
fréquence Gω, C) Le gradient de codage de phase Gφ.Erreur ! Signet non défini.
Figure 1.22 Mise en place des impulsions RF et des trois gradients qui vont se succéder au cours d’une même excitation. .... Erreur ! Signet non défini.
Figure 1.23
... 39 Relation entre l’intensité d’absorption nucléaire (Ip) et la teneur en humidité du bois d’érable à sucre.
Figure 1.24
... 39 Temps de relaxation transversale T2 de l’eau dans des copeaux du thuya occidental à température ambiante en fonction de la teneur en humidité. ○, composante de relaxation transversale lente, T2s; ●, fraction de toute la population des protons qui relaxent avec le temps de relaxation transversale lent; □, composante de relaxation transversale rapide, T2f .
Figure 1.25 Comportement des concentrations d’eau en T2 en fonction de la
teneur en humidité (x – T2 lent, ▲- T2 moyen et ■ – T2 rapide).Erreur ! Signet non défin
Figure 1.26 Courbes séparées de l’eau des lumina des trachéides du bois initial (Bi), de l’eau des lumina des trachéides et des rayons du bois final (Bf) et du bois des parois cellulaires (Pc) exprimés en g d’eau par
cm de bois, en comparaison avec une image MEB de l’échantillon.3 Erreur ! Signet non dé Figure 1.27 Superposition du profil de distribution de l’eau liée en direction
radiale mesurée par RMN avec le profil de la masse volumique du
bois anhydre obtenu par densitométrie aux rayons X.Erreur ! Signet non défini.
Figure 1.28 Profil continu du T2 de l’eau dans le bois juvénile (―) et le bois de duramen (---) de l’épinette blanche... Erreur ! Signet non défini.
Figure 1.29 Profil radial de la distribution d’eau dans les lumina d’un échantillon de bois d’aubier d’épinette blanche à quatre teneurs en
humidité, vis-à-vis d’une image MEB du même échantillon.Erreur ! Signet non défini.
Figure 1.30
... 47 Profil continu de relaxation transversale spin-spin (temps T2) du bois d’aubier (a) à des H sous le PSF et (b) à des H au-dessus du
PSF. éq éq
Figure 1.31
... 51 Image du 14 jour de séchage du bois de chêne blanc produite par le balayeur RMN. Une claire différence de contraste est observée entre le contour et le cœur de chaque planche.
ième
Figure 1.32
... 53 Comparaison des images par RMN des planches de trois épaisseurs durant un cycle de séchage de 14 jours du bois de chêne blanc. Les zones plus blanches indiqueraient une teneur en humidité plus élevée.
Figure 1.33 Image transversale par IRM du bois de frêne... 53
Figure 1.34
... 55 IRM tridimensionnelle de la masse spécifique apparente de l’eau à six temps de drainage.
Figure 1.35
... 55 Vue transversale d’une tranche du pin taeda qui avait été submergé pendant 26 heures.
Figure 1.36
... 56 Image montrant la différentiation des tissus individuels d’une branche de bois de hêtre par l’amélioration du contraste entre tissus ayant des teneurs en humidité différentes. Les zones plus claires signalent des teneurs en humidité plus élevées
Figure 1.37
... 56 Image tridimensionnelle de la distribution spatiale de l’eau dans les tissus individuels d’une branche de bois de hêtre.
Figure 2.1
... 59 Coupes microscopiques du bois d’érable à sucre: (A) coupe transversale (75x); (B) coupe tangentielle (75x) (d’après Panshin et de Zeeuw 1980).
Figure 2.2
... 60 Forme et dimensions des échantillons utilisés : (a) longitudinal (b) radial (c) tangentiel (les limites des cernes y sont montrées).
Figure 2.3
... 64 Schéma de la distribution des cellules de sorption dans les bassins (Hmax : désorption à partir de la saturation intégrale; PSF : désorption à partir du PSF).
Figure 3.1
... 75 Equilibrium moisture content (EMC) as a function of relative humidity for sugar maple wood at 21°C (standard errors did not exceed the symbol size).
Figure 3.2
... 75 Comparison of EMC values of different studies obtained in desorption from full saturation state for sugar maple wood.
Figure 3.3
... 79 MR transverse micro-images of the samples equilibrated at 96% RH. A) From full saturation with an EMC of 31.1%. B) From FSP with an EMC of 28.9%.
Figure 3.4
... 79 MR tangential micro-images of the samples equilibrated at 96% RH. A) From full saturation with an EMC of 31.1%. B) From FSP with an EMC of 28.9%.
Figure 3.5
... 80 MR transverse micro-images of the samples equilibrated at 86% RH. A) From full saturation with an EMC of 21.8%. B) From FSP with an EMC of 20.7%.
Figure 3.6
... 81 MR tangential micro-images of the samples equilibrated at 86% RH. A) From full saturation with an EMC of 21.8%. B) From FSP with an EMC of 20.7%.
Figure 3.7
... 81 MR transverse micro-images of the samples equilibrated at 76% RH. A) From full saturation with an EMC of 17.1%. B) From FSP with an EMC of 17.4%.
Figure 3.8
... 84 A) MR transverse micro-image of one sample equilibrated at 96% RH from full saturation (31.1% EMC). B) SEM image of the same section. The red circle indicates the position of figure 3.9. The white lines indicate the position of figure 3.10A and 3.10B.
Figure 3.9
... 84 SEM transverse image of the limit of growth ring of the sample equilibrated at 96% RH (31.1% EMC) from full saturation.
Figure 3.10
... 85 ESEM transverse image of two different sections of the sample equilibrated at 96% RH (31.1 % EMC) from full saturation.
Introduction
Le bois est la ressource la plus importante de l’industrie forestière et, par ce fait l’approfondissement des connaissances sur son comportement et ses propriétés dans des conditions différentes d’humidité est nécessaire afin de l’utiliser de manière adéquate et optimale.
Le bois est un matériau que se distingue par sa grande hygroscopicité. Il tend comme tel vers un équilibre de teneur en humidité avec l’air qui l’entoure, en restituant ou en prenant de la vapeur d’eau. Cette eau peut se trouver dans le bois sous deux formes: l’eau liée ou hygroscopique et l’eau liquide ou libre. La sorption d’humidité est caractérisée par un point auquel les parois cellulaires du bois sont saturées d’eau liée tandis que les cavités cellulaires sont dépourvues d’eau liquide ; ce point s’appelle « point de saturation des fibres (PSF) ».
En désorption, le comportement du bois aux humidités relatives au-dessus de 60% est fonction de l’état initial de saturation du bois. À partir de l’état saturé, la courbe de sorption d’humidité va se situer toujours au-dessus d’une courbe de désorption normale (à partir du PSF). Ce comportement a été nommé « l'hystérèse à saturation d’humidité ». Cette hystérèse indique que la perte d'eau liée a lieu en présence d’eau liquide, ce que contredit le concept traditionnel du PSF.
Parmi les essais non-destructifs sur le bois une des techniques de plus en plus utilisée est la résonance magnétique nucléaire (RMN). Elle permet d’enregistrer le signal émis suite au changement des spins des protons d’hydrogène présents dans le bois sous forme d’eau, grâce aux phénomènes de relaxation longitudinale et transversale. Ces phénomènes se traduisent en termes des temps de relaxation soit: le temps T1 et le temps T2, respectivement. La RMN a été reconnue par son efficacité pour étudier la distribution de l’humidité dans le bois. Plusieurs études ont en effet démontré qu’elle nous permet d’identifier les types d’eau dans le bois à différentes teneurs en humidité. Dans le bois des feuillus, on retrouve trois composantes du temps T2. Le T2 lent représente l’eau liquide logée dans les lumina des vaisseaux. Le T2 moyen correspond à
l’eau liquide localisée dans les lumina des fibres et du parenchyme. Enfin, le T2 rapide représente l’eau dans les parois cellulaires.
Le signal donné par le RMN peut être transformé dans une image à l’aide de l’imagerie par résonance magnétique (IRM). L’IRM permet de dessiner une carte du temps T2 dans laquelle on arriverait à distinguer l’eau libre de l’eau liée. L’IRM a été utilisée pour visualiser la structure interne du bois, le flux et la distribution de l’eau pendant le séchage, le drainage d’eau sous pression et pour différencier les tissus sains de ceux avec pourriture. La qualité de l’image est définie par des paramètres inhérents à l’appareil utilisé, aux caractéristiques du matériel utilisé et au réglage choisi pour le balayage. En effet, l’image sera le résultat de la combinaison précise de tous ces facteurs, ce qui permettra d’interpréter l’image de manière adéquate.
Le but du présent travail a été de déterminer la localisation de l’eau liquide dans la structure du bois d’érable à sucre au-dessous du point de saturation des fibres. Des essais de désorption ont alors été réalisés entre 58% et 96% d’humidité relative à 21°C à partir de deux états de saturation différents, soit le PSF et la saturation intégrale. Cela nous a permis de comparer des échantillons avec et sans présence d’eau liquide, équilibrés à un même niveau d’humidité relative, en se servant de l’IRM. Ces images ont été aussi couplées avec des images obtenues par microscopie électronique à balayage afin de mieux cerner la localisation de l’eau liquide dans le bois.
Chapitre 1
1. Revue de littérature
1.1 Physique du bois
1.1.1 Domaine hygroscopique
Le bois est un matériau qui se caractérise par son hygroscopicité. Il tend ainsi vers un équilibre de teneur en humidité suite à un échange de vapeur d’eau avec l’air qui l’entoure. Cet équilibre se traduit par une teneur en humidité spécifique, laquelle est fonction principalement de l’humidité relative de l’air ambiant (HR), de la température, de l’histoire de sorption ainsi que de la pression (Bizoň 1995). Cependant, la teneur en humidité d’équilibre du bois peut également être affectée par l’espèce, la teneur en extraits, la masse volumique, le contrefil ainsi que par les contraintes mécaniques (Siau 1995, Hernández 2007a).
Généralement, le bois comporte des molécules d’eau sous deux formes : l’eau liée ou l’eau hygroscopique et l’eau liquide ou l’eau libre. L'eau liée est retenue par des liaisons hydrogène aux parois cellulaires, principalement à la cellulose et aux hémicelluloses, et à un degré moindre aux groupements hydroxyle de la lignine. La quantité d’eau liée est limitée par le nombre de sites disponibles pour la sorption et par le nombre de molécules d’eau qui peuvent être contenues dans un site de sorption (Siau 1995). Elle est ainsi principalement associée à la zone amorphe des chaînes de cellulose (Nakamura et al. 1981). Par contre, l’eau liquide remplit partiellement ou totalement les cavités cellulaires et elle est la première à sortir du bois au séchage car elle est retenue plus faiblement que l’eau liée (Almeida 2006).
Les variables fondamentales qui contrôlent la quantité d'eau dans le bois sont la constitution et la structure du bois elle-même, la température et la concentration de l'eau autour des tissus cellulaires du bois (Urquhart 1960). En effet, la teneur en humidité d’équilibre (Héq) du bois augmente avec l’humidité de l’air ambiant et diminue avec l’élévation de la température. La fonction qui représente ce comportement s’appelle l'isotherme de sorption. Cette courbe exprime la relation entre la quantité de l'eau à l’intérieur du bois et la concentration à l’extérieur, à une température constante (Urquhart 1960). Silva (2007) a déterminé que l’équilibre hygroscopique en désorption des bois de bouleau jaune et de hêtre, a été plus faible à 50°C qu’à 25°C, ce qui confirme l’influence de la température sur l’hygroscopicité du bois. D’ailleurs, ces mêmes variables sont celles qui affectent considérablement les principales caractéristiques physiques du bois. Goulet (1968) et Djolani (1970) ont qualifié les changements des propriétés du bois avec sa teneur en humidité comme «effets de premier ordre de la sorption d’humidité» dans le bois. Ces changements ont lieu principalement dans le domaine hygroscopique, lequel s'étend de l'état anhydre (H=0%), qui constitue la teneur en humidité du bois à une humidité relative de l'air très faible (HR ≈ 0%), jusqu'au point de saturation des fibres (PSF) (aux environs de 30% Héq à 20°C), qui correspond à un équilibre en adsorption dans la vapeur d'eau saturée (HR ≈ 100%) (Hernández 1989).
1.1.2 Hystérèse de la sorption d’humidité
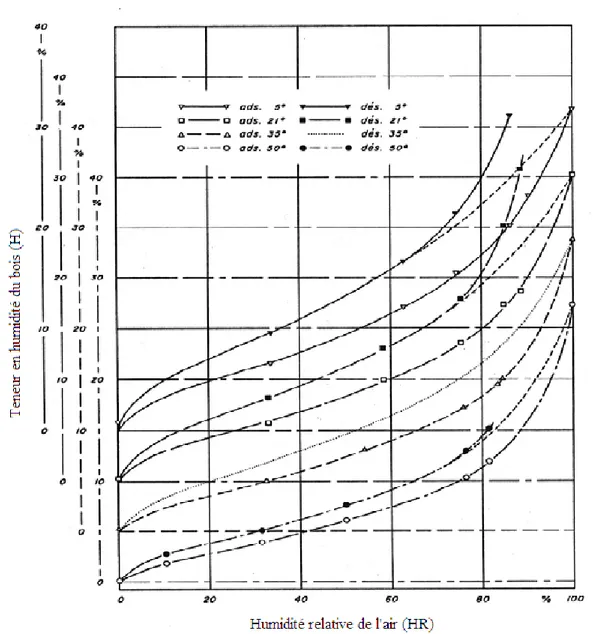
La teneur en humidité d’équilibre du bois n’est pas tout à fait une fonction simple du degré hygrométrique et de la température de l’air ambiant (figure 1.1). Il existe un phénomène appelé « hystérèse de la sorption d’humidité » dans le bois qui décrit la façon dont l’équilibre hygroscopique est atteint, soit par adsorption depuis l’état anhydre ou soit par désorption depuis l’état saturé (Goulet 1967). En effet, la boucle nous indique la valeur limite des écarts entre la perte et la prise d’humidité par le bois. La figure 1.1 montre clairement cette hystérèse de la sorption d’humidité pour le bois d’érable à sucre à une température de 21ºC.
Hystérèse à saturation d’humidité
Hystérèse de la sorption d’humidité
Figure 1.1 Isothermes de sorption d’humidité du bois d’érable à sucre à 21°C (Goulet
1968).
Almeida (2006) décrit la présence de trois zones distinctes dans la courbe de sorption du bois :
La zone à faibles valeurs de HR correspond à la fixation de la première couche de molécules d’eau sur les sites de sorption des membranes cellulaires. Ici, l’énergie de liaison est importante et les molécules d’eau sont peu mobiles,
À des valeurs de HR intermédiaires, les molécules d’eau viennent se fixer sur la première couche déjà absorbée, l’énergie de liaison va ainsi diminuer avec le nombre de couches,
À des valeurs élevées de HR, voisinant 100%, les molécules absorbées par les différents sites vont interagir et former un film liquide continu sur la surface des
pores (condensation capillaire). Lorsque l’humidité relative atteint 100%, la teneur en eau d’équilibre du bois correspondra à celle de la saturation intégrale.
Différentes hypothèses ont été présentées par Djolani (1970) pour expliquer le phénomène d’hystérèse de sorption. Une première théorie simple et valable jusqu’aujourd’hui est celle d’Urquhart et Williams (1924). Elle fait appel essentiellement aux radicaux hydroxyles (OHֿ) des matières ligno-cellulosiques : ces derniers sont les principaux responsables de la sorption des molécules d’eau qu’ils attirent mais ils tendent aussi, quoiqu’à un degré moindre, à s’attirer les uns aux autres c’est-à-dire à se saturer mutuellement. Ces dernières liaisons seraient plus nombreuses à l’état anhydre qu’à l’état saturé et elles seraient aussi plus nombreuses à l’adsorption qu’à la désorption, rendant ainsi l’accès aux radicaux OHֿ plus difficile dans le premier cas que dans le second, ce qui expliquerait également l’hystérèse de la sorption d’humidité.
Une deuxième théorie est celle d’Ant-Wuorinen et Visapää (1960). Ils montrent que la boucle d’hystérèse pourrait également avoir comme origine les différents taux de cristallinité de la matière ligno-cellulosique dont l’effet pourrait être plus que proportionnel aux différences elles-mêmes.
Une autre théorie est celle de Malmquist et Söderström (1996), qui mentionne que la sorption se produit le long des molécules unidimensionnelles de cellulose, principalement dans ses zones amorphes. Cependant, on assume aussi qu’il existe une sorption plus faible dans la zone cristalline. La sorption physique, y compris le gonflement du sorbant, et la sorption chimique de l'eau aux radicaux hydroxyles (OHֿ) libres sont couplées ensemble. La sorption physique est déterminée par la nature du gonflement et par la nature structurale de la zone amorphe des microfibrilles. La sorption chimique dépendra du changement d’accessibilité des radicaux hydroxyles (OHֿ) libres par rapport aux molécules d'eau. Il en résulte qu’il y aura une structure de la matière ligneuse qui sera différente à l’adsorption qu’à la désorption, soit une « structure hystérésique » des membranes ligno-cellulosiques, laquelle présenterait une plus grande cohésion dans le premier cas que dans le second.
Figure 1.2 Isothermes de sorption d’humidité du bois d’érable à sucre aux températures
de 5° 21º 35° et 50°C (d’après Djolani 1970).
L’effet de la température sur la boucle d’hystérèse a été étudié par plusieurs chercheurs. Djolani (1970) a observé que l’hystérèse de la sorption d’humidité du bois d’érable à sucre diminue avec l’élévation de la température entre 5°C à 50°C (figure 1.2). Kolin et Stevanovic (1996) qui ont étudié le comportement de quelques espèces commerciales de feuillus entre 20ºC et 80ºC et de résineux entre 20ºC et 90ºC, sont arrivés à une conclusion semblable. Ils suggèrent que ce phénomène pourrait être expliqué par des changements de la structure interne des microfibrilles de cellulose.
D’autre part, Djolani (1970), Goulet et Fortin (1975) et Laforest et Plamondon (1976) ont rapporté que l’orientation et surtout les dimensions des éprouvettes peuvent affecter la vitesse des échanges d’humidité, mais sans avoir aucune influence sur l’hystérèse de sorption.
1.1.3 Point de saturation des fibres (PSF)
Le point de saturation des fibres (PSF) est une caractéristique très importante du bois. Au début du dernier siècle, Tiemann (1906) l’a défini comme étant la teneur en humidité à laquelle les parois des cellules sont complètement saturées d’eau liée tandis qu’il n’y a pas d’eau libre dans les cavités cellulaires.
D’après Stamm (1964), le point de saturation des fibres est la teneur en humidité au dessous de laquelle les propriétés du bois comme le gonflement, le retrait, la résistance mécanique et la résistance électrique, commencent à changer.
1.1.3.1 Facteurs affectant le PSF
D’après Skaar (1988) on peut citer les facteurs suivants:
La quantité d’extractibles exerce un effet inversement proportionnel sur le PSF, ces substances agissent comme charges ou comme agents de remplissage des pores et même de blocage des voies d’accès à l’intérieur des membranes. Cependant, Hernández (2007a) a mentionné que les extraits du bois peuvent se comporter en tant qu'entités hygroscopiques, hydrophobes ou neutres. La réponse des extractibles à la sorption du bois peut également être affectée par leur localisation et distribution à l’intérieur du bois, par l'humidité relative et par la température à laquelle les mesures sont faites.
Le PSF augmente avec la température jusqu’à 0°C, tandis qu’entre 0°C et 100°C le PSF diminue à un taux moyen de 0,1% H par 1°C d’augmentation (Siau 1995). Le PSF augmente avec la diminution de la masse volumique du bois,
particulièrement au-dessous de 400 kgmֿ3 (Feist et Tarkow 1967). Pour expliquer cela, Čudinov (1981) a divisé l’eau des membranes en humidité liée par attraction moléculaire (fonction des constituants du bois) et humidité liée par capillarité (fonction de l’espace disponible principalement à la surface interne des cellules et
dans la sous-couche S2 des parois cellulaires). L’humidité liée par capillarité serait intimement liée à la masse volumique du bois et conséquemment, un bois moins dense présenterait une plus grande quantité d’espaces disponibles et aurait un PSF plus élevé.
Le contrefil semble provoquer des efforts internes dans le bois, ce qui produirait un effet négatif sur le PSF. Une espèce avec fil droit aurait une valeur du PSF supérieure à une autre avec contrefil (Hernández 2007a).
Le fait que le PSF soit déterminé en adsorption ou en désorption a un effet marqué sur l’estimation de ce dernier, cet écart est de 8% à 10%. Stone et Scallan (1967) ont déterminé que le PSF de l’épinette noire est de 40% Héq par la méthode de la plaque poreuse en désorption, valeur nettement supérieure à la valeur traditionnelle de 30% Héq. Toutefois, plusieurs facteurs peuvent influencer et alors causer des erreurs lors de la détermination du PSF en désorption, comme par exemple l’eau liquide qui peut rester « piégée » dans le lumen des éléments ligneux les moins perméables (Menon et al. 1987; Hernández et Bizoň 1994 ; Hernández et Pontin 2006 ; Almeida et Hernández 2006a,b et Silva 2007).
1.1.3.2 Méthodes de détermination du PSF
Almeida (2006) présente une liste détaillée des diverses méthodes pour déterminer le PSF, chacune ayant un niveau de précision propre. Nous pouvons ainsi citer les suivantes:
Courbes de sorption : extrapolation des isothermes de sorption en adsorption jusqu’à 100% HR. La difficulté de cette méthode est de contrôler l’humidité relative de l’air au-dessus de 98% HR, puisque des changements de température de l’ordre de 0,1°C peuvent avoir un effet significatif sur l’HR. Un autre facteur qui peut influencer la valeur de PSF est la condensation de l’eau dans les cavités cellulaires : le bois présente des capillaires si petits qu’ils provoqueront la condensation à des valeurs d’humidité relative de l’air proches de 99% (Stamm 1971; Siau 1995). Cette condensation sera plus ou moins importante en fonction des espèces, ce qui peut être relié à la distribution des micropores dans la paroi cellulaire. Hernández (2007b) a trouvé que l’essai d’adsorption sur l’eau distillée pour la détermination du PSF n’était pas applicable pour quatre des neuf bois tropicaux étudiés parce qu’il se produisait de la condensation de la vapeur d'eau, même lorsque la température pendant l'adsorption était contrôlée à 0,01°C.
Plaque de tension (en adsorption) : la différence entre cette méthode et d’autres méthodes de sorption est que les teneurs élevées de HR sont contrôlées avec une grande précision (Stamm 1971; Siau 1995).
La teneur en humidité du bois influence de façon importante plusieurs de ses propriétés. L’hypothèse que le PSF délimite le changement de plusieurs propriétés du bois, est utilisée pour déterminer ce point. Quelques propriétés du bois employées pour déterminer le PSF sont le retrait et le gonflement volumique, la résistance mécanique, la résistance électrique, la transmission des rayons bêta, etc. Kelsey (1956) a mesuré les points d’intersection du retrait radial et tangentiel et a noté que, dû à la non linéarité du retrait aux valeurs faibles et élevées d’humidité relative, seulement les mesures faites entre 35% et 65% HR doivent être utilisées pour déterminer ces points d’intersection.
La technique de centrifugation pour forcer l’eau liquide à sortir du bois. L’utilisation de cette méthode sur des espèces imperméables peut générer des valeurs erronées (Perem 1954). Choong et Tesoro (1989) ont observé des valeurs de PSF plus élevées de 30% en utilisant la technique de centrifugation. Ils ont conclu que de l’eau liquide resterait encore dans la structure du bois même avec des temps prolongés de centrifugation à une vitesse très élevée.
Des techniques de résonance magnétique nucléaire (RMN) (Araujo et al. 1994) et de calorimétrie différentielle à balayage (DSC) (Simpson et Barton 1991; Repellin et Guyonnet 2005) peuvent aussi être utilisées pour déterminer le PSF. Ces deux méthodes se révèlent très intéressantes puisqu’elles mesurent directement la quantité d’eau dans les parois cellulaires. Araujo et al. (1994) ont observé que le PSF déterminé par RMN est similaire à celui calculé à partir du modèle de sorption de Hailwood-Horrobin. D’autre part, la technique de la DSC donne des résultats semblables à ceux obtenus par la méthode d’intersection du retrait volumique (Repellin et Guyonnet 2005).
1.1.4 Hystérèse à saturation d’humidité
Une désorption faite à partir de l’état vert ou saturé d’eau donne toujours lieu à une courbe située au-dessus de la courbe de désorption normale (à partir du PSF), aux
hautes humidités. La figure 1.1 montre ce phénomène dans les cas des courbes en désorption obtenues pour les expériences 1 et 2. Ces courbes viennent rejoindre la courbe de désorption normale (expérience 3) à une humidité relative comprise entre 60% et 70% (HR) dépendant des espèces. Goulet (1967) a qualifié ce phénomène comme « l’hystérèse de sorption à saturation ».
Il y a une tendance à attribuer l'hystérèse à la saturation à une première perte irréversible de l'hygroscopicité après le séchage initial du bois vert ou imbibé d’eau. Cependant, plusieurs auteurs ont montré sa présence suite à des désorptions subséquentes (Goulet 1968 ; Fortin 1979 ; Cloutier et Fortin 1991 ; Goulet et Hernández 1991 ; Hernández et Bizoň 1994 ; Hernández et Pontin 2006 ; Almeida et Hernández 2006a,b et Silva 2007).
Goulet et Hernández (1991) ont décrit l'hystérèse à la saturation comme la différence entre l'équilibre obtenu par désorption à partir du PSF et celui atteint par désorption à partir du bois qui contient de l'eau libre. En effet, l'hystérèse à la saturation n'est pas limitée à la première désorption ou désorption initiale, mais plutôt à une perte de l’eau liée en présence de l'eau libre.
L'importance de cette hystérèse variera selon la teneur en humidité initiale du bois. Si la désorption se fait depuis l'état complètement saturé, alors la courbe obtenue correspondra à la teneur en humidité maximale pour une condition d’humidité relative de l’air donnée. Le terme courbe de désorption limite est donc employé pour décrire ce comportement. Ainsi, toute courbe de désorption obtenue à partir d'une teneur en humidité initiale inférieure serait située en-dessous de cette courbe de désorption limite (Hernández et Bizoň 1994, Defo et al. 1999).
La figure 1.3 montre les isothermes de sorption du bois de la pruche de l’Ouest à 21°C à des humidités relatives très élevées. On voit aussi un fort comportement hystérétique, qui selon Fortin (1979) est attribuable à "l'effet de la bouteille d'encre". L’auteur explique que le système capillaire du bois se compose de cavités reliées par des canaux étroits et que la variation des dimensions entre les différents types de cavités reliées en série suggère que la désorption tend à être régie par un potentiel d'eau inférieur, qui est déterminé par les sections plus étroites des pores. Par contre, l'adsorption tend à être
Figure 1.3 Équilibre hygroscopique moyen de l’aubier de pruche de l’ouest à 21°C,
obtenu avec la méthode des plaques et membranes poreuses sous pression, aux humidités très élevées (Fortin 1979).
régie par un potentiel d'eau plus élevé, qui dépend des sections plus grandes des pores (Fortin 1979). La courbe de désorption est donc fortement influencée par le diamètre des canaux qui relient les cavités cellulaires, tandis que la courbe d’adsorption est surtout régie par le diamètre de ces cavités (Almeida 2006).
1.1.5 Effet de l’hystérèse à saturation sur les propriétés du
bois
Tel que mentionné précédemment, le PSF a été défini comme la teneur en humidité d’équilibre au-dessous de laquelle la plupart des propriétés du bois sont affectées, tandis qu’au-dessus de celle-ci ces propriétés demeurent inchangées (Stamm 1964, Skaar 1988, Siau 1995). Ainsi, la résistance mécanique du bois augmente avec la diminution de la teneur en humidité en-dessous du PSF, alors qu’elle reste inchangée au-dessus de celui-ci (Bodig et Jayne 1982). En effet, une perte d’humidité dans les parois cellulaires du bois amène à une augmentation des propriétés mécaniques tandis qu’une perte de
l’eau liquide logée dans les cavités cellulaires n’a aucun effet. Par conséquent, le PSF a été incorporé dans des modèles pour ajuster les propriétés mécaniques du bois en fonction de la teneur en humidité (Bodig et Jayne 1982); de même que dans les modèles d'ajustement du retrait et de la masse volumique du bois (Siau 1984; Skaar 1988). Cependant, plusieurs résultats montrent que cet énoncé n’est pas tout à fait exact car il ne s’agirait pas d’un « point » spécifique mais plutôt d’un intervalle d’humidité. Ainsi, Stevens (1963) a montré que le retrait du bois de hêtre était présent même au-dessus du PSF (figure 1.4). Étant donné que le retrait a été obtenu à l’équilibre d’humidité, l’augmentation identifiant le gradient d’humidité comme étant l’élément déclencheur du retrait semble peu probable.
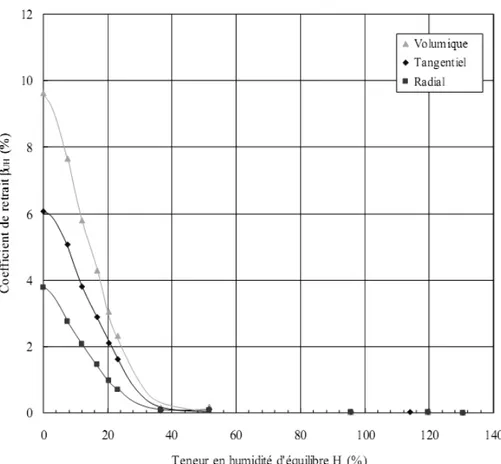
Plus tard, Hernández et Bizoň (1994) ont étudié le retrait transversal et volumique de l’érable à sucre. Le retrait de ce bois a commencé à environ de 42,5% Héq malgré que son PSF était de 31% Héq. Cela indique qu’il y avait de l’eau liquide au moment où la perte de l’eau liée des parois cellulaires a débuté. Cette eau liquide resterait confinée dans les endroits les plus imperméables du bois. La figure 1.5 montre clairement que le retrait du bois débute bien au-dessus du PSF.
D’autre part, Hernández et Pontin (2006), ont étudié la relation entre la teneur en humidité d’équilibre et le retrait pour trois bois tropicaux soit le tornillo, le pumaquiro et le huayruro. Ils ont établi le PSF de chacun à 28%, 22,5% et 21,5% Héq respectivement. En désorption le retrait a commencé à 51,5% Héq pour le tornillo (figure 1.6), 36,3% pour le pumaquiro et 77,3% pour le huayruro. Ainsi, ils ont confirmé que, même à l’équilibre, le retrait du bois débute avant que toute l’eau libre ne soit sortie du bois. Ils ont aussi remarqué que les différences de rétention et de drainage de l’eau libre entre chaque essence sont dues principalement à ses caractéristiques anatomiques propres.
Figure 1.4 Retraits radial et tangentiel du bois de hêtre en fonction de son humidité
d'équilibre en adsorption et en désorption à environ 25°C (d'après Stevens 1963).
Figure 1.5 Coefficients de retrait du bois d’aubier de l’érable à sucre en fonction de sa
teneur en humidité d'équilibre. Le symbole ■ représente la valeur estimée du PSF (d’après Hernández et Bizoň 1994).
Figure 1.6 Coefficients de retrait du bois de tornillo en fonction de sa teneur en
humidité d’équilibre (d’après Hernández et Pontin 2006).
Le travail d’Almeida et Hernández (2006a) sur deux espèces tropicales (congona et cachimbo) et une espèce tempérée (hêtre à grandes feuilles) à également montré que les changements des propriétés physiques du bois débutent à des teneurs d'humidité au-dessus du PSF. Ils suggèrent ainsi qu’à des niveaux élevés de potentiel hydrique, l’eau liquide qui reste encore dans le bois pourrait être enfermée dans les cellules reliées par les plus petits capillaires du bois. Ceci correspondrait aux ouvertures dans les membranes des ponctuations simples situées entre les cellules du parenchyme radial. La rétention de cette eau liquide dans les rayons vient du fait que ces tissus sont considérés comme les moins perméables dans le bois des feuillus (Wheeler 1982, Siau 1984).
Silva (2007) a étudié la relation entre la teneur en humidité d’équilibre obtenue en désorption et les propriétés du bois de bouleau jaune et de hêtre à grandes feuilles à deux températures, soit 25°C et 50°C. L’hypothèse de ce travail était que si la perméabilité du bois augmente avec la température, cela devrait faciliter le drainage de
l’eau confinée dans les sites les plus imperméables du bois. L’humidité du début du retrait devrait alors être plus faible à 50°C qu’à 25°C. Pour le bouleau jaune, le retrait a en effet débuté à 42% Héq à 25°C et à 35% Héq à 50°C. Le PSF pour cette espèce fut estimé à 31% Héq à 25°C et à 29% Héq à 50°C. L’augmentation de la température aurait en effet facilité le drainage de l’eau libre chez cette espèce.
Cela ne fut pas le cas pour le bois de hêtre, où le début du retrait fut noté à 36% Héq à 25°C et à 40% Héq à 50 °C. En effet, la grande proportion des rayons dans cette espèce affecte ses propriétés dimensionnelles et probablement en même temps la teneur en humidité d’équilibre à laquelle le retrait débute. L’augmentation de la température jusqu’à 50°C aurait provoqué un léger collapse dans les rayons, ce qui aurait fait que le retrait à 50°C ait débuté plus tôt qu’à 25°C. Le phénomène de collapse localisé a également été évoqué pour expliquer le comportement particulier du huayruro, un bois présentant du parenchyme longitudinal paratrachéal en couches (Hernández et Pontin 2006).
1.2 Bases théoriques de l’imagerie par résonance
magnétique
1.2.1 Le magnétisme nucléaire
La matière est composée des atomes des éléments spécifiques. La figure 1.7 montre la structure d’un atome. Les atomes sont composés de deux parties importantes, le noyau et les électrons. Le nucléus est composé de deux différentes composantes, protons et neutrons, qui collectivement s’appellent nucléons. Ces nucléons sont maintenus ensemble par les forces nucléaires. Les protons et les neutrons ont environ la même masse, mais les protons sont chargés positivement tandis que les neutrons n’ont pas de charge (Chakeres 1992).
Ces particules ont un mouvement de rotation individuelle autour d’un axe passant par leur propre centre, ce qui induit un moment cinétique. Cette propriété particulière s’appelle « spin ». Plus spécifiquement, à l’intérieur du noyau, les protons (chargés positivement) ont une charge qui tourne ce qui induit autour d’eux un champ
magnétique appelé moment magnétique. La figure 1.8 montre le moment magnétique qui est représenté par le vecteur d’aimantation (Kastler 1997). Seul le noyau d’hydrogène, formé d’un seul proton, joue un rôle important en imagerie aujourd’hui (l’IRM protonique).
Lorsque les protons d’un échantillon sont placés dans un champ magnétique externe Bo,
ils vont avoir tendance à s’orienter selon la direction de ce dernier. Ils vont se distribuer en deux populations tournant autour de Bo: l’une orientée dans le même sens que Bo
(parallèle) et l’autre dans le sens contraire à Bo (antiparallèle) (Kastler 1997) (figure
1.8).
Au total, les protons orientés dans le sens parallèle sont toujours en excès et ils vont être à l’origine de l’apparition d’un vecteur d’aimantation M . À l’équilibre, ce vecteur est aligné sur Bo selon OZ (Kastler 1997) (figure 1.9).
1.2.2 Mouvement de précession et fréquence de Larmor
Les spins ont un mouvement de rotation et décrivent un cône autour de l'axe de : c'est le mouvement de précession. On peut apparenter le mouvement de précession au mouvement d'une toupie qui bascule et reste en équilibre de rotation (Hoa 2007) (figure 1.10). En effet, les protons précessent autour de
o B
o
B avec un angle donné, ce qui fait apparaître une composante longitudinale Mz (aimantation longitudinale); à l’équilibre, on l’appelle Mzo (Kastler 1997).
La vitesse de précession est proportionnelle à l'intensité du champ magnétique et elle est caractérisée par la fréquence de précession ω
o B
0 (nombre de rotations par seconde): ω0= γ B0, où ω0 est la fréquence angulaire de Larmor et γ est le rapport gyromagnétique (spécifique à chaque noyau) (Hoa 2007).
Figure 1.7 La structure de l’atome (adapté de : ttp://www.danieltrimpey.com/learn/electronics/index.htm).
Sens parallèle
Sens antiparallèle
Figure 1.8 Le spin et moment magnétique du proton (adapté de : http://chemistry.umeche.maine.edu/CHY431/NMR/NMR-3.html).
Figure 1.9 Les protons s’orientent selon la direction d’un champ magnétique extérieur
(Bo) avec l’apparition d’un vecteur d’aimantation macroscopique M (Σ =Mzo) (adapté de Kastler 1997).
Figure 1.10 Mouvement de précession (adapté de Kastler 1997).
1.2.3 Résonance magnétique nucléaire (RMN)
La résonance magnétique nucléaire (RMN) consiste à étudier les modifications d’aimantation des noyaux d’une substance sous l’action conjointe de deux champs magnétiques: un champ magnétique statique fixe élevé (Bo) et un champ
électromagnétique tournant (B1) (onde électromagnétique ou de radiofréquence RF)
(Kastler 1997).
1.2.4 Phase d’excitation et de relaxation
Il peut y avoir une interaction entre une onde de radiofréquence (onde RF) et les spins en précession, laquelle s’appelle phénomène de résonance. Il correspond à un transfert d'énergie entre deux systèmes ayant la même fréquence. L'onde RF va ainsi produire un phénomène de résonance seulement si sa fréquence est la même que celle de la précession des spins. Elle va apporter de l'énergie au système de spins, ce qui est la phase d'excitation (Hoa 2007). La figure 1.11 montre des protons au cours de l’application d’une onde RF 90°, où il y a une transition de protons du niveau de basse énergie E1 vers le niveau de haute énergie E2 (les protons parallèles s’orientent antiparallèles) ; cela s’appelle la mise en « phase » des protons (Kastler 1997).
Les ondes RF utilisées en imagerie par résonance magnétique ont des durées très brèves, de l’ordre de la milliseconde. Ce sont des « impulsions ». Le transfert d’énergie de
l’onde RF aux noyaux se traduit par un basculement de l’aimantation totale M par rapport à sa position initiale. On utilise le plus souvent des impulsions de 90° et 180° (Doyon 2001). La figure 1.12 montre les effets des impulsions RF sur le vecteur d’aimantation macroscopique M . Une impulsion de 90° bascule M dans le plan x0y (on l’appelle ). Une impulsion de 180° inverse le vecteur d’aimantation (on l’appelle - ) (Kastler 1997). xy M zo zo M M
Lorsque l'émission de radiofréquence (onde RF) est interrompue, le système va restituer l'énergie absorbée pour retourner à l'état d'équilibre de départ: c'est la phase de relaxation (Hoa 2007). C’est grâce à la relaxation des protons que le phénomène de RMN devient observable.
La figure 1.13 montre tout le cycle d’interaction entre l’onde de radiofréquence et les protons placés dans le champ . Au début, les protons soumis au champ magnétique vont se mettre en état d’équilibre apparaissant ainsi une composante longitudinale de l’aimantation. Tout de suite, un apport d’énergie (excitation) par une onde RF (impulsion de 90°) dont la fréquence est égale à la fréquence de Larmor (condition de résonance) entraîne, par égalisation des protons sur les deux niveaux d’énergie et mise en phase des spins, respectivement une disparition de la composante longitudinale
o B o B zo M z M
et une apparition d’une composante transversale Mxy de l’aimantation.
Cet état est instable, et dès la fin de l’excitation, il va y avoir un retour à l’état d’équilibre (stable) au cours duquel les phénomènes inverses vont avoir lieu. D’une part, progressivement, par transitions inverses E2 → E1 (antiparallèles → parallèles), l’aimantation longitudinale zM repousse progressivement (relaxation T1). D’autre part, par déphasage rapide des spins, l’aimantation transversale Mxy décroît rapidement
Figure 1.11 L’onde RF provoque des transitions du niveau de basse énergie E1 au
niveau de haute énergie E2 (adapté de Kastler 1997).
Figure 1.12 Effet des impulsions RF sur le vecteur d’aimantation macroscopique
Figure 1.13 Interaction entre l’onde de radiofréquence et les protons placés dans le
champ Bo (adapté de Kastler 1997).
1.2.5 Temps T1 et T2
La phase de relaxation n’est pas instantanée. Son évolution au cours du temps est caractérisée par deux temps, T1 et T2, qui varient suivant l’état normal du tissu (Doyon 2001). D’une part, progressivement, par transitions inverses (antiparallèles – parallèles), l’aimantation longitudinale Mz repousse progressivement (relaxation T1). D’autre part par déphasage rapide des spins, l’aimantation transversale Mxy décroît rapidement
(relaxation T2) (Kastler 1997).
1.2.5.1 La relaxation longitudinale (T1)
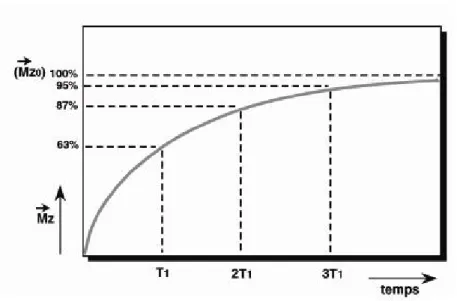
La relaxation longitudinale, dite relaxation T1 ou encore relaxation « spin-réseau », correspond au retour à l’équilibre énergétique du système après l’excitation (J.F. Le Bas 2008). Le T1 caractérise la repousse de l’aimantation longitudinale Mz au cours du temps. Il correspond, après l’arrêt d’une impulsion de 90°, au temps mis par Mz pour atteindre 63% de sa valeur d’équilibre (Doyon 2001) (figure 1.14). Le T1 varie avec la
structure moléculaire ainsi que l’état solide ou liquide de la matière. Il croît également avec l’intensité du champ magnétique statique Bo (Kastler 1997).
1.2.5.2 La relaxation transversale (T2)
La relaxation transversale est due au déphasage des spins, dès l’arrêt de l’impulsion RF. Le déphasage est lié aux interactions spins-spins qui créent des hétérogénéités de champ, et donc de fréquence de précession. Alors, l’aimantation transversale Mxy
décroît rapidement. C’est ce qu’on appelle la relaxation transversale (T2) ou également relaxation « spin-spin ». Cette chute de l’aimantation transversale suit une courbe exponentielle décroissante caractérisée par le temps T2. Le T2 correspond au temps mis par l'aimantation transversale pour revenir à 37 % de sa valeur initiale, en d’autres termes elle correspond à 63% de décroissance (Hoa 2007) (figure 1.15). Ce temps de relaxation T2 est toujours inférieur au temps de relaxation T1. Il dépend lui aussi de la mobilité des atomes ou des molécules sur lesquelles ces atomes d'hydrogène sont engagés (J.F. Le Bas 2008).
xy
M
1.2.6 Signal en IRM
1.2.6.1 Signal de précession libre (FID)
La détection de l’aimantation nucléaire est effectuée en plaçant dans le plan perpendiculaire à Bo, une bobine de détection ou « antenne ». Le mouvement de
rotation de l’aimantation transversale crée dans l’antenne un courant électrique induit, que l’on peut mesurer après amplification, et qui constitue le signal de RMN. Ce signal, appelé signal d’induction libre ou Free Induction Decay (FID) (figure 1.16), apparaît
sous forme d’oscillations amorties car l’aimantation transversale, qui est la seule à induire un courant, décroît au cours du temps. Le signal est positif ou négatif selon que l’extrémité du vecteur d’aimantation est dirigée vers l’antenne ou en sens inverse. La mesure de ce courant induit, permet la détermination des paramètres caractéristiques du signal RMN : sa fréquence, sa durée, son amplitude et éventuellement sa phase (Doyon 2001).
Figure 1.14 Courbe exponentielle de repousse de l’aimantation longitudinale en
fonction du T1 (Kastler 1997).
Figure 1.15 Courbe exponentielle de disparition de l’aimantation transversale en
Figure 1.16 Signal de précession libre (adapté de Kastler 1997).
1.2.6.2 Notion de T2*
Si le champ était parfaitement homogène et si l'échantillon placé dans ce champ n'induisait aucune inhomogénéité, la courbe de décroissance de la composante transversale permettrait d'avoir accès directement au temps de relaxation T2. En fait, le champ n'est jamais parfaitement homogène (J.F. Le Bas 2008). En plus des inhomogénéités de champ d’origine moléculaire dû au déphasage des spins (T2), le champ magnétique a des inhomogénéités d’origine « instrumentale » ou « propre » qui sont constantes et qui vont entraîner un déphasage encore accru des spins. Ainsi le signal de FID observé est lié à la fois aux inhomogénéités du champ d’origine moléculaire, auxquelles s’ajoutent les inhomogénéités propres (constantes) du champ magnétique externe . On utilise le symbole T2* pour représenter la conjonction de ces deux effets (Kastler 1997) (figure 1.17).
o B o B o B o B o B
1.2.6.3 Séquence d’écho de spin
La méthode pour s’affranchir des inhomogénéités propres du champ principal externe et rétablir la cohérence de base des spins s’appelle la séquence d’écho de spin (Hoa 2007).
o B
Figure 1.17 Notion de T2* (adapté de Kastler 1997).
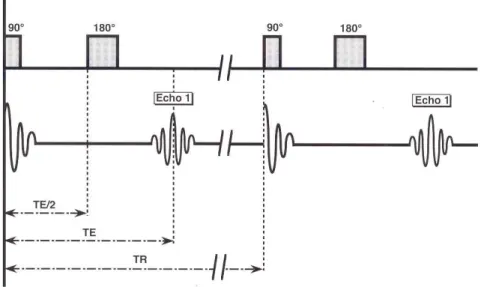
La figure 1.18 montre la chronologie du phénomène de l’écho de spin. Au temps t=0, tout de suite après une impulsion de 90°, le vecteur Mxy ou MTm (T pour transversal) est
maximal. Ensuite, les spins se déphasent rapidement à cause des inhomogénéités de champ d’origine moléculaire et aussi à cause des inhomogénéités propres de Bo et le
vecteur MT diminue ainsi en T2*. Au temps TE/2 (demi-temps d’écho) une impulsion
RF de 180° est appliquée, celle-ci va inverser les déphasages sans modifier le sens de rotation. Les spins qui précessaient le plus rapidement vont se retrouver à présent derrière les plus lents et comme les vitesses de précession restent identiques, au bout d’un temps 2 TE/2 = TE (temps d’écho), les spins seront de nouveau en phase, engendrant un signal dit d’écho de spin, lequel peut être mesuré. Cependant, les spins ne sont pas parfaitement rephasés. Ce déphasage est la conséquence de la relaxation
spin-spin (irréversible), le vecteur MT'm (signal mesuré) est légèrement plus petit que MTm
(Kastler 1997).
Toutefois le cycle qu’on vient de décrire, ne permet d’obtenir qu’une ligne d’une matrice d’image. Pour obtenir les lignes suivantes (128 ou 256 lignes) il est nécessaire de répéter, pour chaque ligne, le cycle d’impulsions de 90° et 180° (figure 1.19) (Kastler 1997).
Chaque séquence élémentaire de deux impulsions peut être répétée après un intervalle de temps TR appelé « temps de répétition » (figure 1.19) (Doyon 2001). Kastler (1997) mentionne que le TR correspond au temps de passage d’une ligne sur l’autre mais également au temps de repousse de l’aimantation longitudinale. Les deux paramètres TE et TR, appelés paramètres d’acquisition, sont choisis pour privilégier un signal dépendant préférentiellement soit de T1 ou soit de T2 (Doyon 2001).
1.2.7 Contraste en T1, T2 et densité protonique
Le contraste en IRM correspond à la traduction des signaux RMN (aimantation) en niveaux de gris (noir : signal faible, blanc : signal élevé). Ce contraste traduit les différences en temps de relaxation et, dans une moindre mesure, les différences en densité des protons. Les trois facteurs T1, T2 et densité des protons interviennent toujours à des degrés divers. L’operateur peut, par le choix des paramètres de la séquence, favoriser l’un de ces facteurs dans la génération du signal, c’est-à-dire « pondérer » la séquence (Kastler 1997).
Kastler (1997) donne une équation du signal de spin écho simplifiée:
Dans cette équation, le signal est proportionnel à la densité protonique. Il dépend également de deux fonctions dont l’une, , traduit la décroissance exponentielle en
T2 de MT (modulée par le TE) et l’autre, , traduit la repousse exponentielle en
T1 de ML (modulée par le TR). Enfin, le signal dépend également du flux dans les
voxels.
1.2.7.1 Signal pondéré en densité protonique
Si le temps de répétition TR est « long », soit très supérieur aux valeurs des temps T1 des échantillons examinés, l’intensité du signal de mesure est pratiquement indépendante de T1. Avec un TE « court », choisi nettement inférieur aux valeurs des T2 des échantillons, l’intensité du signal mesuré ne dépend pratiquement pas non plus de T2 : on dit alors que le signal est pondéré en ρ (Doyon 2001).
1.2.7.2 Signal pondéré en T2
Par contre, avec un TE « long », choisi du même ordre de grandeur que celui des temps T2 des échantillons, associé à un TR « long », l’intensité du signal dépend des valeurs de ρ et T2 : on dit alors que le signal est pondéré en T2 (Doyon 2001).
Figure 1.18 Chronologie des événements au cours du phénomène de l’écho de spin
(d’après Kastler 1997).
Figure 1.19 Chronologie des impulsions RF et paramètres de la séquence d’écho spin
1.2.7.3 Signal pondéré en T1
Si le TE est nettement inférieur à T2, l’intensité du signal ne dépend pratiquement pas des valeurs des temps T2 des échantillons examinés. Il suffit alors de choisir un temps de répétition TR « court » pour que l’intensité du signal dépende essentiellement de T1 et ρ. Le signal obtenu est dit pondéré en T1 (Doyon 2001).
1.2.8 Formation de l’image
La création des images en résonance magnétique requiert l’ajout d’une information spatiale au signal pour attribuer une position aux différents signaux. Pour cela, on choisit d’abord le plan de coupe au sein duquel seront ensuite définies les directions horizontale et verticale (Hoa 2007).
Les champs de vue, représentant la taille du plan de coupe, varient de 1 à 50 cm. Leur dimension maximale est principalement déterminée par la dimension des antennes d’émission et de réception (Doyon 2001).
Un voxel est le volume réel dans l'espace où le signal mesuré se produit. Il est défini tant par sa position dans le champ magnétique que par ses dimensions. Les données de chaque voxel sont affichées par l'écran d'ordinateur comme une matrice de lignes et de colonnes qui correspondent toutes les deux à la position du voxel dans le champ magnétique et à l'intensité du signal qu’il produit. Dans l'affichage de l'image, les points de données dans les colonnes et les lignes s’appellent pixels d'image. Quand le signal du voxel est plus fort, plus brillant est l'intensité de chaque pixel sur l'écran. Le nombre de lignes (Lp ou Np) et de colonnes (Cf ou Nf) de pixels qui forment l'image s’appelle la matrice image (figure 1.20). En général, une matrice image est constituée de 128, 256 ou 512 lignes et de 128 à 512 colonnes (Chakeres 1992). Les dimensions du champ de vue et la taille de la matrice déterminent la résolution spatiale (dimension du pixel) (Kastler 1997).
En IRM, la localisation spatiale du signal de l’image fait appel à l’utilisation de gradients. Un gradient représente le taux de variation (pente) d’une donnée physique dans une direction de l’espace. Les gradients linéaires de champs magnétiques sont
utilisés pour le codage spatial de l’image. Ces gradients créés par des bobines appelées bobines de gradients (où passe un courant électrique), vont se superposer au champ magnétique principal Bo. Le champ magnétique principal augmente ainsi de façon
linéaire suivant la direction où le gradient est appliqué (Kastler 1997).
1.2.8.1 Le codage de phase et fréquence
Pour réaliser une image par IRM, il faut d’abord, à l’intérieur d’un volume donné, sélectionner un plan de coupe dans lequel on applique un premier gradient de champ (linéaire) appelé gradient de sélection de coupe Gss (ou Gz) (Kastler 1997). Le codage de phase et de fréquence ont pour but la localisation du signal dans les deux directions de la coupe sélectionnée. La première direction de la coupe est repérée par la méthode de « codage par la fréquence ». Elle consiste à appliquer, pendant l’acquisition du signal, un gradient de champ Gω perpendiculaire au gradient de sélection de la coupe. Les protons situés dans la coupe, sur une ligne perpendiculaire à la direction du gradient Gω, ont alors la même fréquence de résonance. La seconde dimension du plan de coupe est « codée par la phase ». Dans cette méthode, un gradient Gφ de durée et d’amplitude déterminés, est appliqué immédiatement après l’impulsion d’excitation. Il a pour effet de modifier progressivement la phase des moments magnétiques élémentaires situés dans la direction de ce gradient (Doyon 2001) (figure 1.21).
Un gradient de codage par la phase est appliqué avant chaque acquisition. Pendant l’acquisition des signaux, un gradient de codage par la fréquence est appliqué dans une direction perpendiculaire au gradient de codage par la phase. Plusieurs signaux sont acquis successivement par répétition d’une séquence élémentaire. D’un signal à l’autre, l’intensité du gradient de codage par la phase est changée. On obtient ainsi une matrice de signaux qui, après transformation de Fourier, donne l’image de la coupe (Doyon 2001).
1.2.8.2 La transformée de Fourier
Une transformée de Fourier est un outil qui convertit des fonctions de temps à fonction de fréquence. C’est une technique mathématique pour convertir des données du domaine de temps en données du domaine de fréquence et vice versa. Le processus
Figure 1.20 Champ de vue, matrice et plan de coupe (Kastler 1997).
A B
C
Figure 1.21 A) Le gradient de sélection de coupe Gss, B) Le gradient de fréquence
ressemble à un musicien qui entend un ton (le signal du domaine de temps) et il détermine quelle note (la fréquence) est jouée (Hornack 2008). Elle nous permet alors d’extraire les différentes fréquences individuelles d’un signal complexe contenant plusieurs fréquences (Kastler 1997).
Pour décomposer une image en 2 dimensions, on effectue une transformée de Fourier 2D. La première étape de la transformée de Fourier 2D consiste à appliquer une transformée de Fourier 1D dans une première direction (par exemple ligne par ligne). L‘étape suivante consiste à appliquer une nouvelle transformée de Fourier 1D sur le résultat obtenu à la première étape, selon la deuxième direction cette fois-ci (colonne par colonne) (Hoa 2007). Cette technique d’acquisition de l’image consiste donc à associer un codage par la phase à un codage par la fréquence (Doyon 2001). Le résultat de la transformée de Fourier 2D d’une image est le plan de Fourier. Le plan comprend les informations d'intensité et de phase de chaque composante fréquentielle de l’image, selon les axes x et y (Hoa 2007).
1.2.8.3 Durée d’acquisition d’une séquence IRM
La mise en place des impulsions RF et gradients serait comme suit (figure 1.22) :
1. L’impulsion de 90° et simultanément la sélection du plan de coupe par Gss (l’impulsion est alors sélective dans le plan de coupe déterminé) ;
2. L’impulsion de 180° et le codage par la phase des lignes de la matrice par GΦ ; 3. Le codage par la fréquence des colonnes de la matrice par Gω qui doit coïncider avec la réapparition du signal (lors du rephasage des spins par l’impulsion de 180°), c’est-à-dire au moment de la lecture du signal.
Tout ce cycle, qui décrit les événements au cours d’une même excitation, ne permet d’obtenir l’information que d’une première ligne de la matrice image. Cette matrice généralement comporte 256 lignes. Pour obtenir les lignes suivantes il faut répéter selon la même chronologie l’ensemble des événements de la séquence. Le temps de passage d’une ligne sur l’autre (d’un cycle à l’autre) n’est nul autre que le temps de répétition TR. Mais, pour améliorer la qualité de l’image il est possible de repasser plusieurs fois
Figure 1.22 Mise en place des impulsions RF et des trois gradients qui vont se
succéder au cours d’une même excitation (Kastler 1997).
sur chaque ligne avant d’aller à la ligne suivante en répétant l’ensemble des événements de la séquence. Le nombre de passages sur chaque ligne est appelé nombre d’accumulations ou d’excitations Nex. Alors, pour calculer le temps d’acquisition Tac nécessaire pour obtenir une image, il faut le temps de répétition TR, le nombre de lignes Np de la matrice et le nombre d’excitations Nex. Tac = TR x Np x Nex (Kastler 1997).
1.2.9 Facteurs de qualité de l’image en IRM
1.2.9.1 Le rapport signal sur bruit (S/B)
Le rapport signal sur bruit (S/B) est fonction de la moyenne de l’intensité du signal par rapport à l’importance de la variation du bruit. Le bruit est la fluctuation aléatoire indésirable de l'intensité du signal qui est toujours présent. Il vient perturber le signal émis par les protons excités de la coupe d’intérêt (Hoa 2007). Le bruit est un signal indésirable produit par l'échantillon, l'environnement externe et le système électronique de résonance magnétique. L’imagerie par résonance magnétique dépend avant tout du rapport S/B et lui-même est une fonction de divers facteurs complexes (l’intensité du champ, la séquence d’ondes RF, les caractéristiques propres de l’échantillon, l’antenne utilisée, le nombre d’excitations et la bande passante de réception). Ce rapport est une mesure de la qualité du signal : plus haut est le rapport, meilleure est la qualité d'image.